
长沙地区龟甲竹高生长及光合特性研究
2017-11-17 03:02钟新科
湖南林业科技 2017年3期
钟新科,黄 滔,唐 红
(1.益阳市资阳区林业局, 湖南 益阳 413000; 2.湖南省森林植物园,湖南 长沙 410116)
长沙地区龟甲竹高生长及光合特性研究
钟新科1,黄 滔2,唐 红2
(1.益阳市资阳区林业局, 湖南 益阳 413000; 2.湖南省森林植物园,湖南 长沙 410116)
研究了湖南省森林植物园内龟甲竹的出笋规律、幼竹高生长规律以及光合特性。龟甲竹出笋和高生长规律的研究结果表明:龟甲竹发笋期分为明显的初期、盛期和末期 3 个时期,一般在 3 月中旬开始发笋,历期33 天左右;2016 年,园内龟甲竹共发笋 48 支,其中由于人为损伤和营养不良,5 支退笋;龟甲竹幼竹在高生长过程中,呈现“慢—快—慢”的“S”型生长规律,符合 Logistic 方程;龟甲竹一天中总平均生长量为 17.65 cm,白天生长量更高。光合特性研究结果表明:龟甲竹净光合速率在 12∶ 00 时达到最大值,呈“单峰型”;其光响应与 CO2响应的结果表明龟甲竹利用弱光的能力较强,但 CO2同化能力弱,总体光合能力较弱;PAR、Gs、Tr、Ta 和 Ci 是影响龟甲竹净光合速率日变化的主要生理因子,且影响效应逐渐减弱。
龟甲竹;发笋规律;幼竹高生长规律;光合特性;生理生态因子
龟甲竹(Phyllostachys heterocycla),其秆直立,粗大,高 2.5—4.5 m,属刚竹属[1],外表灰绿色,竹竿下部节间歪斜且突出,犹如龟甲,故称“龟甲竹”,是我国珍稀观赏竹种之一。在生产中,龟甲竹成活率较高,但繁殖率很低。因此,了解其生长特性,科学合理地栽培管理对其繁殖具有重要意义。
目前,竹类植物的研究主要集中于高生长规律及光合特性的研究[2-13],关于竹的分子生物学[14-15]和无性繁殖[16-17]等方面研究鲜有报道。在竹林实际经营过程中, 光合特性和生长规律是培育竹类作物的重要理论基础,大多数竹种的高生长规律均呈现“S”型生长趋势,但不同竹种间高生长的时间、高度、各阶段的历期又有不同。受生理、温度、湿度、地理条件等的影响[3-7],同一地区不同竹种间或不同地区相同竹种间,其发笋规律和光合特性差异明显,且影响光合作用的主要因子也不尽相同。因此,本研究以湖南省森林植物园观赏竹园内龟甲竹为试材,对其发笋规律、高生长规律以及光合特性进行了研究,旨在为龟甲竹在实际生产中的栽培措施、引种培育和科学经营管理提供理论基础,为其广泛应用于园林绿化奠定基础。
1 材料与方法
1.1 研究地概况
湖南省森林植物园地处 113°00' E,28°20'N,海拔 70~85 m,年平均降雨量 1 412.3 mm;年均气温 17.2 ℃,其中极端高温为 40.6 ℃,极端低温为 -11.3 ℃;全年无霜期 281 天。其观赏竹园内,土壤类型为第四纪网纹红壤, pH 值 5.6 左右。
1.2 试验材料
试验材料为生长于湖南省森林植物园观赏竹园内的 2 年生龟甲竹,园内共栽植龟甲竹 50 株。
1.3 试验方法
1.3.1 龟甲竹发笋规律及幼竹高生长规律 2016年发笋期间,每天定时调查 50 株母竹的发笋数,并做好标记,同时记录萌发笋在的成竹及退笋情况。
选择 4 株具代表性的发笋盛期萌发笋,从发笋开始,每天 6∶ 00、18∶ 00 分别测量株高,分析其高生长规律。并在高生长时期的某天从 6∶ 00开始直至次日 6∶ 00,每隔 2 h 测量一次株高,记录龟甲竹幼竹昼夜生长情况。
1.3.2 龟甲竹光合特性研究
(1)光响应曲线和 CO2(ACI)响应曲线测定
于 2016 年 8 月 25 日、9 月 8 日和 9 月 20 日的 09∶ 00—11∶ 00,选取 3 株生长旺盛且长势一致的健壮植株,使用 Li-6400 光合测定仪(美国LICOR 公司)测定竹中部 3 片向阳叶片的净光合速率,结果取平均值。
设定光合测定仪的 CO2浓度固定值为 400 μmol·mol-1,空气相对湿度为 70%,叶室温度为 25 ℃,气体流速为 0.5 L·min-1,分别在 PAR(光合有效辐射)为 0、25、50、100、150、200、400、600、800、1 000、1 200、1 400、1 600 和 1 800 μmol·m-2·s-1的条件下测定叶片的净光合速率( Pn,μmol·m-2·s-1),从而拟合龟甲竹的光响应曲线。
测定 CO2(ACI)响应曲线时,安装 CO2钢瓶后,稳定 20 min,设置光源的光合有效辐射为1200 μmol·m-2·s-1,温度为(25±1)℃,气体流速为 0.5 L·min-1,空气相对湿度为 70%,分别在胞间 CO2浓度(Ci)为 50、100、200、300、400、600、800、1 000 和 1 200 μmol·m-2·s-1的条件下测定叶片 Pn。
(2)光合参数和环境因子日进程的测定
于 2016 年 8 月 23 日、8 月 30 日和 9 月 6 日的 06∶ 00—18∶ 00 选择 3 株生长旺盛长势一致的健康植株,进行不离体测试,测量其中 3 片叶片的光合日变化,每叶片重复记录 3 组数据。
光合参数包括净光合速率和蒸腾速率( Tr,mmol·m-2·s-1)的日变化,并计算叶片水分利用效率(WUE,mmol·mol-1,Pn/Tr)日变化。同时测定生理因子气孔导度( Gs,mmol·m-2·s-1)和胞间 CO2浓度(Ci,μmol·mol-1)的日变化,以及部分环境因子:光合有效辐射(PAR,μmol·m-2·s-1)、气温(Ta,℃)、空气相对湿度(RH,质量分数)和大气 CO2浓度(Ca,μmol·mol-1)。
1.3.3 数据分析 应用 Excel 2010、SPSS 17.0软件对试验数据进行统计分析。此外,以叶子飘等[18]的光合作用新模型进行光响应和 CO2(ACI)响应曲线的拟合。
2 结果与分析
2.1 龟甲竹发笋规律
龟甲竹于3 月12 日开始发笋直至4 月13 日终止发笋,共发笋 48 支,历期 33 天,具体发笋情况见表 1。根据不同时期下发笋数的方差分析结果,龟甲竹的发笋时间明确划分为发笋初期(3月 12 日至 3 月 20 日)、发笋盛期(3 月 21 日至4 月 1 日)和发笋末期(4 月 2 日至 4 月 13 日)3个阶段,且不同阶段间差异极显著(P<0.01)。另外,由于机械损伤和营养缺乏等原因,共退笋 5 支。
2.2 龟甲竹幼竹高生长规律及昼夜生长规律
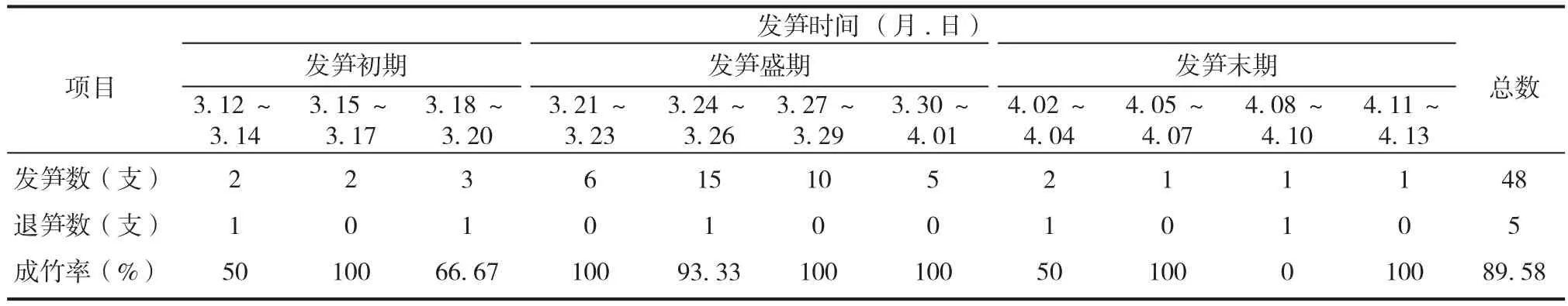
表1 不同发笋时期龟甲竹发笋、退笋统计Tab.1 Shooting and shoots degradation of Phyllostachys heterocycla at different phases
选择 4 支具代表性的萌发笋,记录其高生长数据,绘制龟甲竹的高生长曲线(见图 1)。结果表明,龟甲竹幼竹高生长过程为典型的“S”型生长曲线,即历经初期、快速期和平台期;4 支龟甲竹幼竹株高最大可达 546.5 cm,最小为 449.5 cm,平均株高为 489.95 cm。到达平台期的时间最长为34 天,最短为 27 天。利用 logistic 方程,拟合了龟甲竹幼竹高生长方程为:H = 534.663/(1- e(3.862-0.184t)) (R2=0.999),较好的模拟了龟甲竹幼竹高生长过程,其中 H 为株高,t 为生长时间。
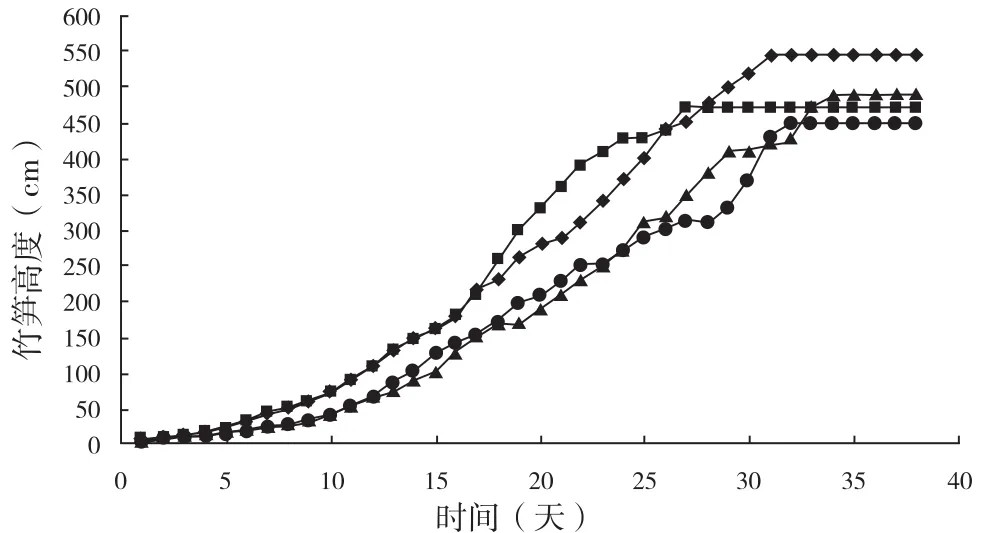
图1 龟甲竹幼竹高生长规律Fig.1 The young bamboo height growth of Phyllostachys heterocycla
4 支萌发笋在 24 h 内的生长变化如表 2 所示,结果表明,白天期间(6∶ 00~18∶ 00),龟甲竹幼竹在 14∶ 00 之前生长量保持平稳增加,在 14∶ 00~16∶ 00 生长量迅速增加至峰值(3.95 cm),随后生长量又降低;夜间(18∶ 00 ~ 次日6∶ 00),龟甲竹生长量的峰值(2.46 cm)出现在 20∶ 00~22∶ 00 时,之后生长量逐渐降低并维持一定的增长速度。24 h 内竹笋总生长量的平均值为 17.65 cm,其中白天生长量是夜间生长量的 1.34 倍,龟甲竹幼竹白天和夜间的平均生长速率分别为 0.84 cm/h 和 0.63 cm/h。
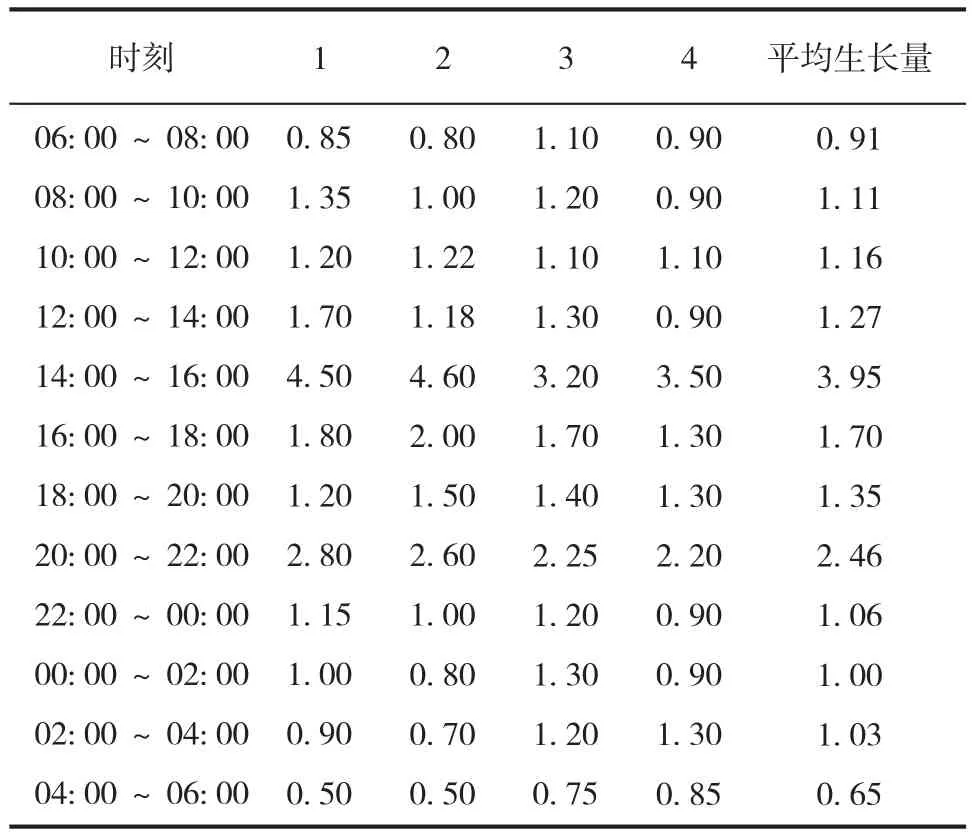
表2 龟甲竹幼竹昼夜高生长量Tab.2 The young bamboo height growth of Phyllostachys heterocycla during 24 hours(cm)
2.3 龟甲竹光合特性研究
龟甲竹 Pn 的光响应曲线和 CO2响应曲线见图 2、图 3,分别以 PAR 和 CO2浓度为自变量X,Pn 为因变量 Y,利用叶子飘等[18]的光合新模型进行拟合,获得了龟甲竹光响应回归方程和 CO2响应回归方程,具体参数见表 3。
龟甲竹在 PAR 相对较低的情况下,光合速率增加更快,当 PAR 高于 200 μmol·m-2·s-1时,其光合速率便基本达到平台期,说明龟甲竹相对而言对于弱光的适应和利用能力可能较强。随着CO2浓度的升高,龟甲竹净光合速率增加的速率几乎不变,说明龟甲竹的 CO2同化能力可能较弱。由图 2 和图 3 可知龟甲竹净光合速率较低,说明其光合能力相对较弱。
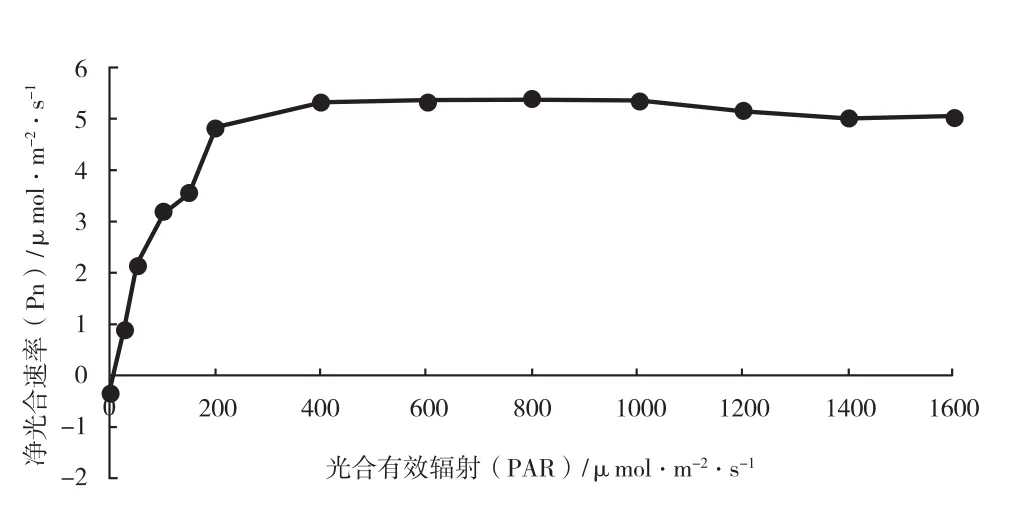
图2 龟甲竹光合-光响应曲线Fig.2 Light response curves of Phyllostachys heterocycla photosynthesis

图3 龟甲竹光合-CO2响应曲线Fig.3 CO2 response curves of Phyllostachys heterocycla photosynthesis

表3 龟甲竹的光响应参数和CO2响应参数Tab.3 The parameters of light response and CO2 response of Phyllostachys heterocycla
龟甲竹的净光合速率日变化以及环境因子的日进程变化见图 4。环境因素是植物光合作用的主要影响因子。由图 4 可知,Ta 在 6∶ 00 至 10∶ 00先上升,10∶ 00 至 16∶ 00 几乎维持不变,16∶ 00 后下降;与 Ta 相反,RH 先下降,在 10∶ 00 后维持不变,16∶ 00 后又逐渐升高;Ca 先下降,12∶ 00时为最低值,维持一段时间后,在 16∶ 00 后迅速上升;PAR 在 12∶ 00 前迅速上升至峰值,之后逐渐降低,18∶ 00 时 PAR 仍高于早上 8∶ 00 前的 PAR 值。
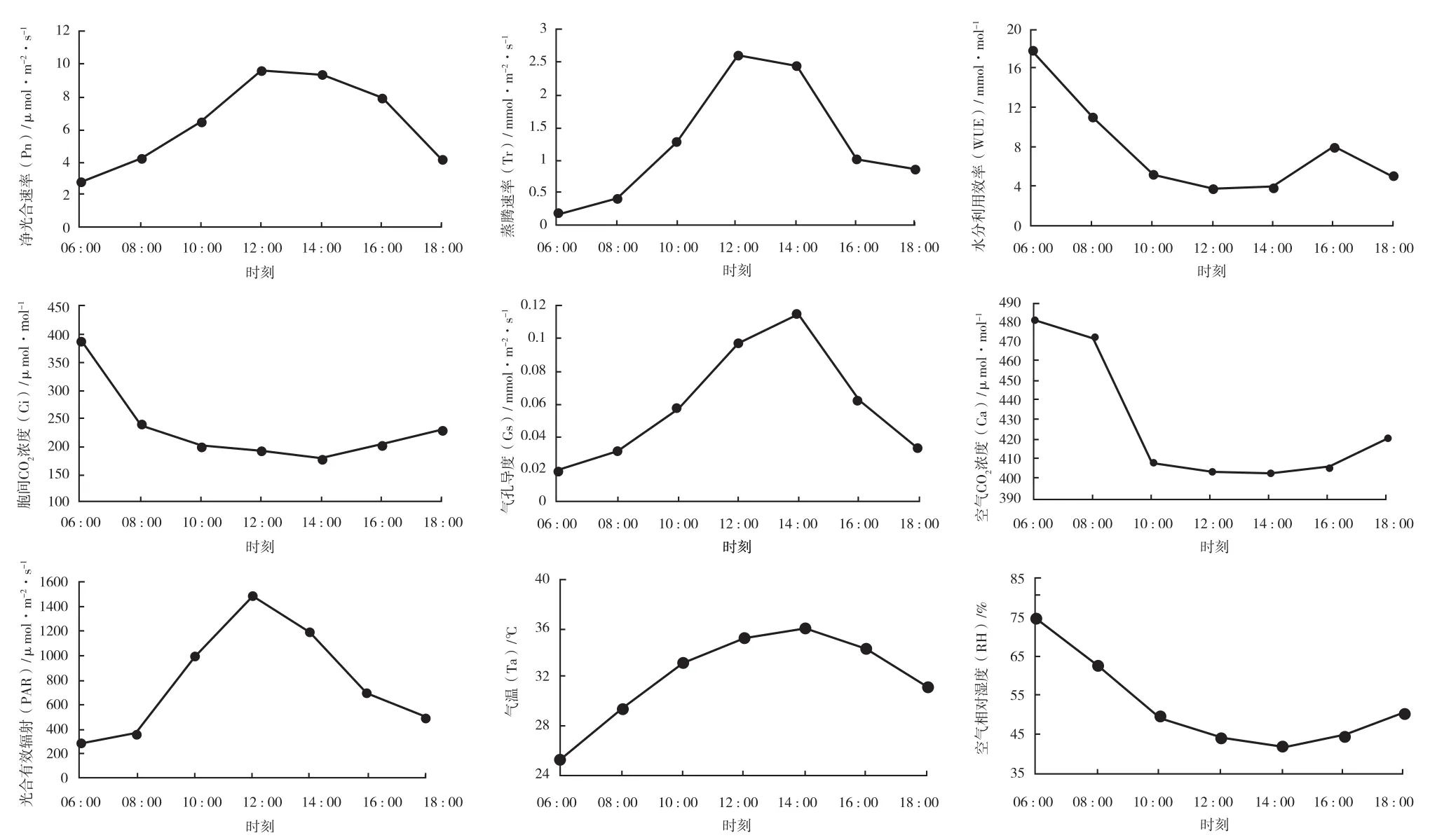
图4 龟甲竹Pn和生理生态因子的日进程变化Fig.4 The net photosynthetic rate and physio-ecological factors daily change of Phyllostachys heterocycla
龟甲竹净光合速率在 10∶ 00 前迅速增加,10∶ 00 至 12∶ 00 之间缓慢增加,12∶ 00 达到最大值,之后便逐渐降低,日变化曲线整体呈“单峰型”;12∶ 00 前 Tr 逐渐上升,WUE 逐渐下降,12∶ 00 后 Tr 一直下降,WUE 先升高,至16∶ 00 后又下降;Gs 在 12∶ 00 前逐渐上升,之后逐渐下降,Ci 在 14∶ 00 前逐渐下降后迅速上升。
2.4 龟甲竹净光合速率与部分生理生态因子的相关性分析
由龟甲竹 Pn 与各生理生态因子的简单相关分析可知(见表 4),Pn 与各生理生态因子均存在显著或极显著相关性。但由于环境因子对 Pn 的影响复杂多变,简单相关并不能准确地反映各因子与 Pn 的相关关系,因此,本研究分析了各因子与Pn 的偏相关性(见表 5),结果表明龟甲竹的 Pn与各环境因子的偏相关性与简单相关性不完全一致。利用逐步多元回归确定影响龟甲竹 Pn 的主要因子,以 Gs(X1)、Ci(X2)、Tr(X3)、Ta(X4)、Ca(X5)、RH(X6)和 PAR(X7)为自变量,龟甲竹的 Pn 为因变量 Y,得到了龟甲竹的最优回归方程为:

结果表明 PAR、Gs、Ta、Ci 和 Tr 等是影响龟甲竹净光合速率的主要因素。
通径分析能反映出各因子对 Pn 和各因子间的复杂相互作用关系。由通径分析结果可以看出(见表 6),所有因子对龟甲竹净光合速率产生的直接效应均为正效应;而由于因子间的间接效应相对复杂,综合直接与间接效应,PAR 对龟甲竹 Pn的影响效应最大,其次依次为 Gs、Tr、Ta 和 Ci。
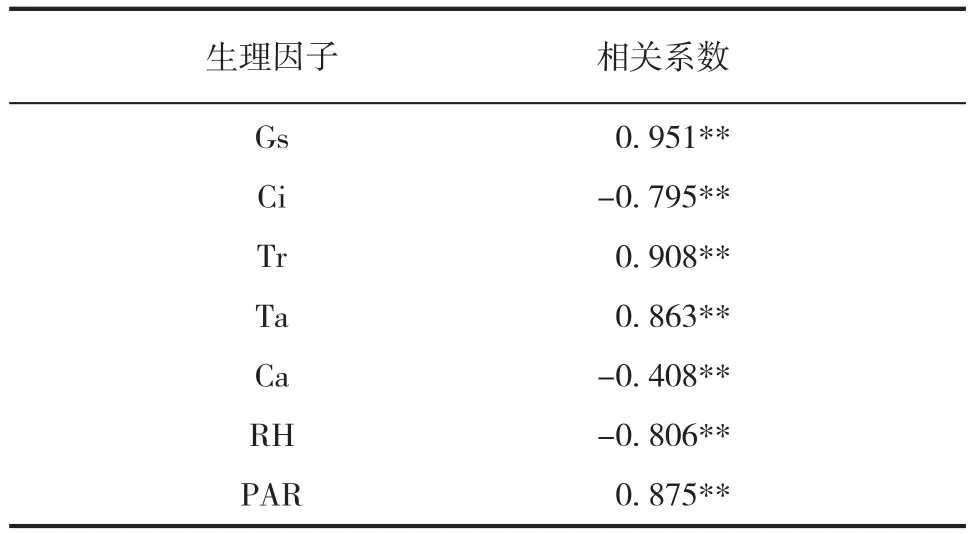
表4 龟甲竹净光合速率与生理生态因子间的简单相关性Tab.4Correlation of net photosynthesis rate with some physio-ecological factors in Phyllostachys

表5 龟甲竹净光合速率与生理生态因子间的偏相关性Tab.5Partial correlation analysis of the net photosynthesis rate with the factors in Phyllostachys heterocycla
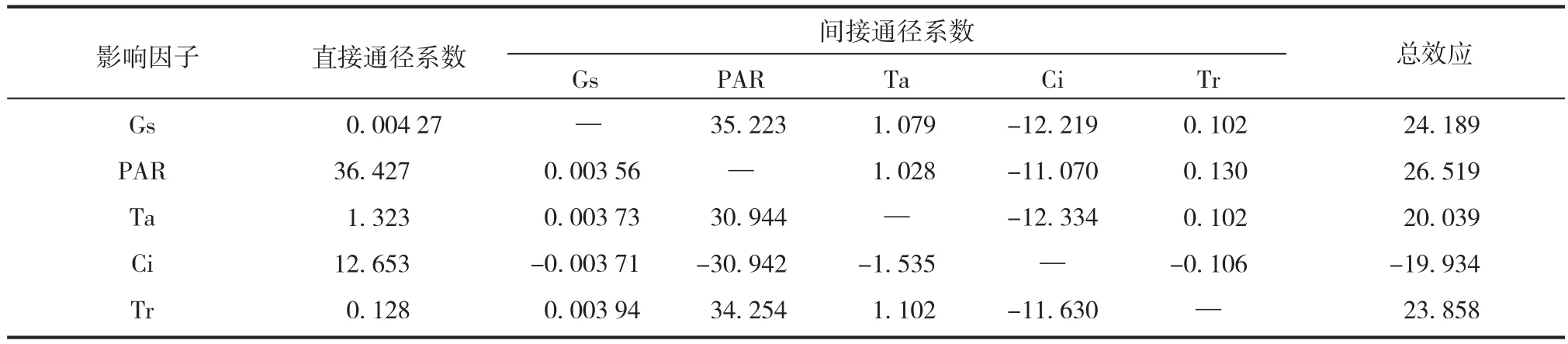
表6 影响龟甲竹净光合速率的因子的通径分析Tab.6 Path analysis of the net photosynthesis rate with main influence factors in Phyllostachys heterocycla
3 结论与讨论
本研究中龟甲竹从3 月12 日开始发笋,4月13 日终止发笋,发笋期为 33 天。许多竹类[5-6]的研究结果表明,温度和降水量是影响竹发笋的主要影响因子,因此造成二者初始发笋时间不同的原因可能是因为研究时间和地点不同,使得发笋期间温度与降水量产生了较大差异而引起的。与大多数竹类的研究结果一致[4,7,8,10],龟甲竹发笋数变化也明显分为 3 个不同时期,且差异极显著。另外,本研究中共有 5 支退笋,其中 2 支退笋均为发笋末期萌发笋,营养供应不足是造成退笋的主要原因;由于 3 月中旬到 4 月中旬,湖南森林植物园举办世界名花生态文化节,游客较多,由于人为的机械损伤导致了 3 支退笋。
龟甲竹幼竹高生长过程呈“慢—快—慢”的“S”型生长曲线,本研究利用 Logistic 方程很好地拟合了龟甲竹幼竹的高生长过程,为制定龟甲竹抚育管理措施提供了重要的理论基础。黄纹竹[3]、圣音竹[4]和篌竹[7]均可以利用 Logistic 模型较好的模拟其高生长过程。与黄纹竹[3]和筇竹[9]的昼夜生长规律研究结果一致,龟甲竹白天生长量也大于夜间生长量,该结果与撑绿竹[8]的研究结果相反。温度和湿度会影响竹子高生长过程,龟甲竹在春季发笋,黄纹竹、筇竹也是春季发笋,而撑绿竹是夏秋季发笋,春季发笋由于白天温度更适宜竹笋的高生长,因此白天生长量相对更高;夏秋季发笋,白天温度过高会加速竹子的蒸腾作用,而夜间温度和湿度更利于幼竹的高生长,夜间生长量更高。
植物的光合特性是植物生产中重要的指导理论,主要取决于其自身的遗传特征,环境因子也同时对植物的光合作用产生影响[19]。本研究与李迎春等[12]的研究一致表明,龟甲竹的净光合速率在一天中仅有一次峰值,利用弱光的能力强,但光合能力整体较弱。另外,本研究还拟合了龟甲竹的 CO2响应曲线,表明龟甲竹同化 CO2的能力不强。
由于生理生态因子与植物净光合速率间,以及生理生态因子间均存在着复杂的相互作用关系[20-22],因此,本研究与李迎春等[12]的研究结果并不完全一致。比如,净光合速率虽然都为“单峰型”,但峰值出现的时间相差较大,本研究中峰值出现在正午 12∶ 00,而后者出现在上午 9∶ 00;采用通径分析的方法,本研究认为在长沙地区,光合有效辐射对龟甲竹的净光合速率影响效应最大,其次依次为气孔导度、蒸腾速率、气温和胞间 CO2浓度,而后者认为影响龟甲竹的因子,按影响效应大小依次为大气湿度、叶温、气孔导度和光合有效辐射。二者研究结果的差异,可能是由于龟甲竹在不同地区生长,为适应环境,光合特性发生了相应的变化。
因此,在实际生产中应该按照“因种制宜,因地制宜”的原则,确定龟甲竹的光合特性和主要的影响因子,制定合理的栽培措施,有规划地指导生产管理工作,增强其观赏性并得到广泛应用。
[1]马乃训,张文燕.中国珍稀竹类[M].杭州:浙江科学技术出版社,2007.
[2]刘 玮,黄程前,黄 滔,等.长沙地区观赏竹引种筛选研究[J].经济林研究,2014,31(4):132-138.
[3]楼 扬,孙 祥,江津凡.黄纹竹出笋及幼竹高生长规律研究[J].世界竹藤通讯,2010,8(4):7-10.
[4]唐 红,刘 玮,黄 滔,等.圣音竹发笋及幼竹高生长规律研究[J].中南林业科技大学学报:自然科学版,2015,35(8):27-31.
[5]毕红玉,李宜文,李晓英,等.淡竹发笋与幼竹高生长规律的研究[J].山东农业大学学报:自然科学版,2010,41(3):340-343.
[6]潘寅辉,高立旦,虞敏之,等.四季竹发笋及幼竹高生长规律研究[J].竹子研究汇刊,2006,25(1):27-29,36.
[7]陈玉华,宋丁全.篌竹出笋成竹生长规律研究[J].南京林业大学学报:自然科学版,2002,26(3):43-47.
[8]付建生,董文渊,韩 梅,等.撑绿竹出笋规律探析[J].林业科技开发,2007,21(3):42-45.
[9]董文渊,黄宝龙,谢泽轩,等.筇竹生长发育规律的研究[J].南京林业大学学报:自然科学版,2002,26(3):43-47.
[10]黄 滔,唐 红,刘 玮,等.长沙地区 3 种优良观赏竹发笋及幼竹高生长规律[J].经济林研究,2016,34(2):114-119.
[11]余红兵,曾 磬,肖润林,等.花叶芦竹光合特性日动态及其影响因素分析[J].湖南农业大学学报:自然科学版,2013,39(2):200-203.
[12]李迎春,杨清平,陈百林,等.龟甲竹光合生理特性及其与主要影响因子关系[J].林业科技开发,2011,25(2):35-39.
[13]徐振国,郭起荣,冯 云,等.淡竹林分结构与林分因子相关研究[J].中南林业科技大学学报:自然科学版,2014,34(2):72-76.
[14]陈 容,张 丽,曹 颖,等.低温胁迫下梁山慈竹再生植株叶绿素荧光特性和耐寒转录因子的表达[J].南京林业大学学报:自然科学版,2014,38(4):39-44.
[15]蒋 瑶.慈竹 CBF1 基因克隆及其耐寒性研究[D].成都:四川农业大学,2012.
[16]李在留,辉朝茂.珍惜竹种巨龙竹组织培养研究[J].林业科学,2006,42(2):43-48.
[17]杨海芸,王晓芹,张 宁,等.日本花叶矢竹组织培养与叶色变异研究[J].竹子研究汇刊,2010,29(4):15-20.
[18]叶子飘,高 峻.光响应和 CO2响应新模型在丹参中的应用[J].西北农林科技大学学报:自然科学版,2009,37(1):128-134.
[19]李丽芳,吴晓敏,王立峰.植物光合生理生态学研究进展[J].陕西师范大学学报:自然科学版,2007,21(3):71-75.
[20]Coste S,Roggy J C,Imbert P,et al.Leaf photosynthetic traits of 14 tropical rain forest s pecies in relation to leaf nitrogen concentration and shade tolerance[J].Tree physiology,2005,25(9):1127-1137.
[21]邹学校,马艳青,张竹青,等.环境因子对辣椒光合与蒸腾特性的影响[J].植物资源与环境学报,2005,14(4):15-20.
[22]李永华,张开明,于红芳.10 个秋菊品种的光合特性及净光合速率与部分生理生态因子的相关性分析[J].植物资源与环境学报,2012,21(1):70-76.
(文字编校:张 珉)
Young bamboo height growth and photosynthetic characteristics of Phyllostachys heterocycla in Changsha,Hunan
ZHONG Xinke1,HUANG Tao2,TANG Hong2
(1.Ziyang District Bureau of Foresty,Yiyang City,Yiyang 413000,China;2.Hunan Forest Botanical Garden,Changsha 410116,China)
In this paper, the regularity of shooting,height-growth and photosynthetic characteristic of Phyllostachys heterocycla were studied in Hunan Forest Botanical Garden.The results of shooting and the regularity of young bamboo height-growth indicated that the shooting time was from mid-March to mid-April,which lasting about 33 days,and could be divided into three phases,including shooting initial phase,abundant phase and final phase.The total shooting number was 48,the total shoot degradation was 5,and the reason was artificial mechanical damage and short of nutrition.The height growth rate of young bamboo was slow,then was fast,and then was slow,and the logistic equation of height growth fitted with the h eight growth data.During the high growth period,the height growth at a d ay amounted to 17.65 cm,while the daytime height growth was higher than that in the night.The results of photosynthetic characteristic indicated that Phyllostachys heterocycla had a stronger ability to utilize the weak light,but its photosynthetic ability was weak.The abilities of CO2assimilation of Phyllostachys heterocycla were weak and it had a narrow range for adapting to the CO2concentration.The daily change of net photosynthetic rate was a one-peak curve,and the peak value was at 12∶ 00.The correlation results pointed out main influence factors for Pn of Phyllostachys heterocycla and ranked the factors by the effects from the highest to the lowest as follow:PAR,Gs,Tr,Ta and Ci.
Phyllostachys heterocycla;shooting regularity;young bamboo height growth regularity;photosynthet ic characteristi-cs;physio-ecological factors
S 795.7
A
1003-5710(2017)03-0015 -06
10.3969/j.issn.1003-5710.2017.03.003
2017-03-27
中央财政林业科技推广示范科研项目([2013]XT03)
钟新科(1980-),男,湖南益阳人,工程师,主要从事园林绿化及园林植物培育研究
猜你喜欢
江西农业(2022年8期)2022-05-13
绿色科技(2021年21期)2021-11-26
内蒙古林业调查设计(2021年3期)2021-07-06
家教世界·创新阅读(2020年6期)2020-07-09
———在龟甲、兽骨上刻字的朝代
家教世界(2020年16期)2020-07-08
中成药(2018年10期)2018-10-26
现代园艺(2018年1期)2018-03-15
新课程·下旬(2017年7期)2017-08-14
西北林学院学报(2015年5期)2015-01-03
植物营养与肥料学报(2011年2期)2011-10-26
