
四川桤木杂交子代苗期性状遗传分析及选择
2018-03-09 06:29杨汉波郭洪英王泽亮李佳蔓肖兴翠
四川林业科技 2018年1期
杨汉波,郭洪英,陈 炙,王泽亮,黄 振,李佳蔓,肖兴翠
(四川省林业科学研究院,四川 成都 610081)
四川桤木(AlnuscremastogyneBurk.)是我国桤木属11个种中最重要的一个特有种,极具开发潜力[1-3]。四川桤木生长快,木材用途广,是重要的速生优质用材树种;根部具根瘤,能固定空气中游离的氮,对土壤改良效果明显,是理想的生态防护和混交造林树种[2]。20世纪60年代以来,在浙江、湖南和湖北等长江中下游地区广泛引种栽培,现已成为我国长江流域水网地区防护林、丘陵山区水土保持林和短周期工业原料林基地建设的重要阔叶树种[4]。国内对四川桤木遗传变异的研究较少。陈益泰等[4]发现四川桤木存在极其丰富的地理变异和个体变异。王军辉等[2,5]分析了四川桤木优良种源/家系生长适应性和遗传稳定性,指出树高、材积、木材基本密度和纤维长度的种源差异显著,而家系间差异不显著。卓仁英等[6]建立了四川桤木DNA提取和PCR条件。任保青等[7]和杨汉波等[8]对四川桤木开展了染色体核型分析,均表明四川桤木的体细胞染色体数为56,为多倍体木本植物。
随着生物技术的不断发展,新的育种途径也不断出现[9]。但杂交育种仍为最有效的育种方法,将是目前及今后相当长时间培育四川桤木新品种的重要手段[10,11]。而杂交育种作为一种重要的育种手段在四川桤木遗传改良方面的应用未见相关报道。为此,以12个四川桤木优树和1个欧洲桤木优良品种为亲本,按照NCⅡ交配设计进行控制授粉,根据1 a生杂种苗的苗期生长性状测定结果,对各遗传参数进行估算,以期为速生、丰产四川桤木优良品种的选育提供依据。
1 材料与方法
1.1 试验地概况
试验地位于四川省林业科学研究院唐昌基地,地理坐标为东经103°49′18″,北纬39°55′13″,海拔600 m,年均气温16℃,年日照1 014 h,年均降雨量979.4 mm,土层深厚,含水层一般厚15 m~25 m。
1.2 材料
以收集的12个四川桤木优树及引种的1个欧洲桤木优良品种作为杂交亲本,采用NCⅡ交配设计组成42个组合(家系)(见表1)。于2016年3月进行控制授粉,12月采集种子,4℃贮藏。
2017年2月进行营养杯内播种育苗,试验采用完全随机区组设计,每个小区100杯,重复3次,定期浇灌。8月调查杂交子代的苗高和地径。
表1供试四川桤木的6×7 NCⅡ交配设计
Tab. 1 6×7 NCⅡ cross design for excellent breeding inAlnuscremastogyneBurk.

母本父本YO2TJ9LD8YO7TT4JT2XH5JG1F1F7F13F19F25F31F37BZ6F2F8F14F20F26F32F38SW2F3F9F15F21F27F33F39PC8F4F10F16F22F28F34F40JG6F5F11F17F23F29F35F41JT4F6F12F18F24F30F36F42
1.3 数据分析
采用SPSS 20.0和excel软件对获得的试验数据进行处理,利用方差分析和多重比较进行家系间的差异分析。一般配合力(GCA)、特殊配合力(SCA)、广义遗传力和狭义遗传力等遗传参数的估算借助DPS软件进行。
2 结果与分析
2.1 杂交子代家系间及家系内各性状的差异性
家系间的苗高和地径变异大小见表2。苗高家系间变幅为5.85 cm~27.53 cm,种间杂交组合JT4×YO2子代苗高最高;地径家系间变幅为1.21 mm~3.05 mm,JT4×JT2子代地径最大,种间杂交组合JT4×YO2子代地径也处于较高的水平,为2.58 mm。苗高和地径在家系间存在显著差异,具有较大的选择潜力。苗高家系内变异系数在9.24%~43.48%之间,平均为20.82%;地径家系内变异系数在12.85%~39.51%之间,平均为21.72%,表明家系内也存在较大的变异,有必要进行家系内选择。由表3可知,四川桤木杂交种苗苗高和地径在家系间的差异均达到极显著水平,说明不同家系的苗高和地径存在真实的遗传差异。母本的苗高和地径方差均达到极显著水平,而父本的苗高和地径方差均不显著。母本×父本在苗高上的方差达到显著水平,而在地径上的方差不显著。
表2四川桤木杂交子代各性状在家系间和家系内的变异
Tab. 2 Variation of hybrid offspring traits among and within families ofA.cremastogyne

家系苗高地径均值/cm0.05水平CV/%均值/cm0.05水平CV/%F17.70opqrs21.271.67fghijklmn34.42F28.72mnop43.481.54ijklmno39.51F36.73rst18.811.33mno20.50F412.84fg16.351.86efghijk18.26F510.61ijk19.042.03efg13.14F627.53a26.342.58bcd23.95F77.73opqrs19.631.76efghijklm19.87F811.22hij19.871.87efghijk20.05F97.48opqrs28.471.40lmno27.43F108.13nopqrs21.661.66fghijklmn23.21F1117.71c17.422.47cd12.85F1211.77ghi16.961.36lmno19.04F139.49klmn18.741.58ghijklmno19.58F1412.27gh17.521.90efghij21.38F156.77rst20.311.38lmno21.19F168.10nopqrs19.331.49jklmno22.65F1713.86ef21.872.19de19.50F1817.63c15.012.58bcd15.78F198.54mnopq30.191.57hijklmno29.20F2015.39d19.522.07ef17.22F218.77lmnop13.751.69fghijklmn20.70F228.49mnopq22.191.88efghijk18.75F2310.82ijk23.861.81efghijkl19.72F2410.26jkl19.511.53ijklmno21.33F255.85t9.241.28no20.49F269.45klmn21.231.68fghijklmn18.10F277.34pqrs18.441.56hijklmno27.31F289.33klmn15.051.79efghijkl15.89F298.71mnop21.961.78efghijkl19.43F309.81jklm24.531.95efghi22.50F318.28mnopqr40.681.50ijklmno37.75F3215.09de16.852.59bc16.86F337.04qrst23.681.21o36.20F349.82jklm18.881.78efghijklm23.32F358.25mnopqr18.091.65fghijklmn19.22F3615.40d27.603.05a121.78F379.06lmno14.572.00efgh22.25F387.63opqrs22.301.75fghijklm17.87F396.62st16.701.39lmno17.94F407.59opqrs15.641.44klmno22.99F4112.39gh14.902.50cd13.66F4223.59b22.972.94ab19.60
注:“0.05水平”一列下标相同字母表示差异不显著,标不同字母表示差异显著(P<0.05)。
表3四川桤木杂交子代各性状的方差分析结果
Tab. 3 Variance analysis of hybrid offspring traits ofA.cremastogyne

变异来源区组家系母本父本母本×父本苗高df2415630M.S.0.15587.19∗∗79.87∗∗8.7037.63∗地径df2415630M.S.0.265.80∗∗0.69∗∗0.080.33
注:**表示P<0.01极显著水平,*表示P<0.05显著水平。
2.2 配合力效应分析
对13个亲本估算一般配合力效应值(GCA),结果见表4。GCA效应值在同一亲本不同性状间及同一性状不同亲本间存在明显的差异,如YO2的苗高GCA效应值为15.36,而地径GCA效应值为负值,JT2的苗高GCA效应值为负值,而地径GCA效应值为正,其值为7.14。母本JT4、JG6、BZ6和父本LD8、XH5的苗高和地径GCA效应值均为正值,其中母本JT4的苗高和地径GCA效应值均最大,分别为54.72和24.51,其次为母本JG6,苗高和地径GCA效应值分别为9.83和12.37。
对24个家系各性状的特殊配合力(SCA)效应分析结果表明(见表5),不同家系在各个性状上的差别较大,苗高和地径的SCA效应值变幅分别为-55.85~86.99和-45.84~34.43。由表5还可知,F6在苗高上具有最大特殊SCA效应值,其次为F42和F11;F36在地径上具有最大SCA效应值,其次为F32和F42。F42和F32在苗高和地径上的SCA效应值均较高,它们的杂种优势较为明显。
表4四川桤木13个亲本的一般配合力效应值

Tab. 4 Estimated values of GCA of 13 A.cremastogyne parents
2.3 各性状遗传力及性状间的相关性
从图1可以看出,四川桤木杂交苗在1 a生时苗高的广义遗传力较高,为77.36%,地径的广义遗传力为29.97%,表明该群体各性状的遗传潜力较大,从进行选择不但是可行的,而且从苗高性状能够获得较大的遗传增益。
对1 a生四川桤木杂交苗的苗高和地径进行相关分析结果表明(见图2),苗高与地径间存在显著正相关,相关系数为0.6798,说明苗高与地径密切相关。因此,如果选择生长量高的优良家系,可以根据苗高和地径进行联合选择。
表5四川桤木不同家系各性状的特殊配合力效应值
Tab. 5 Estimated values of SCA in the traits of the families ofA.cremastogyne
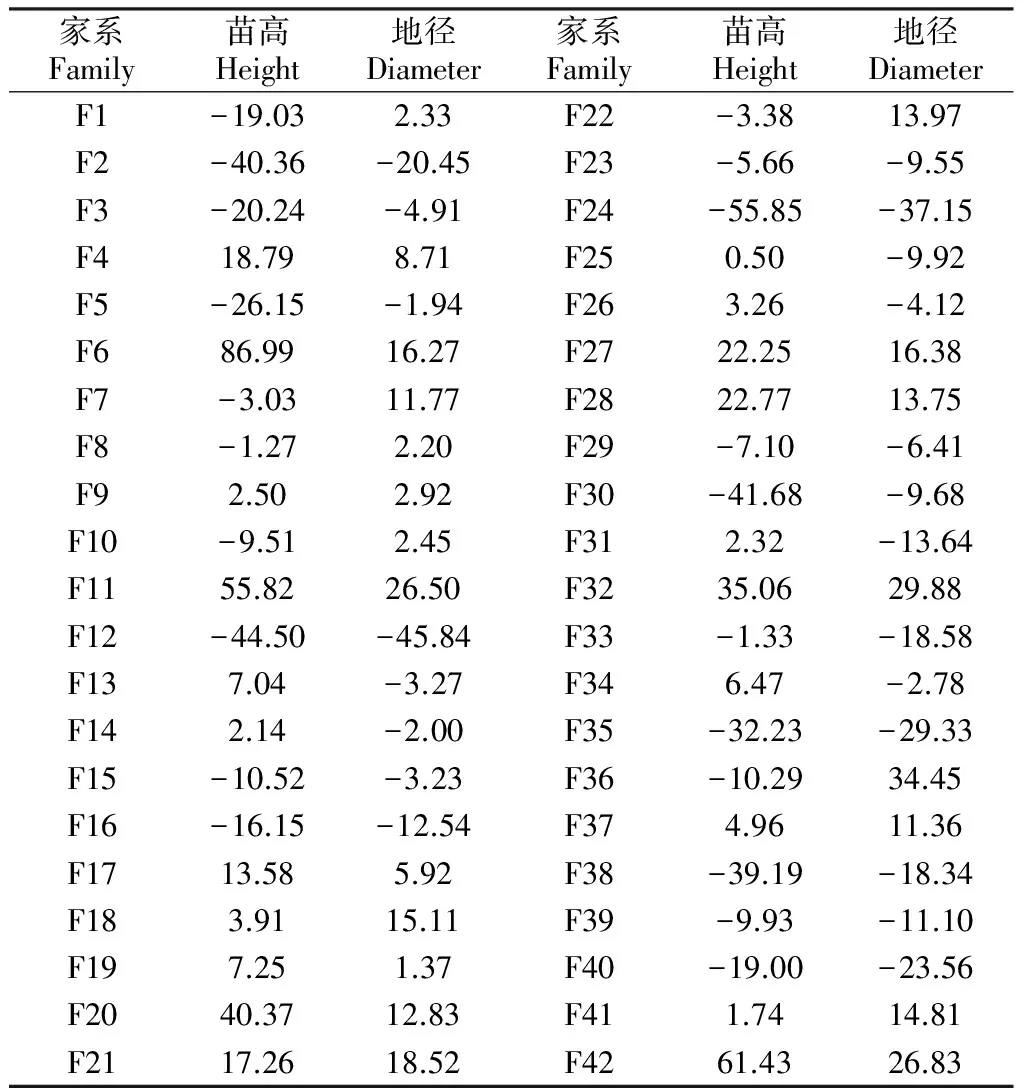
家系Family苗高Height地径Diameter家系Family苗高Height地径DiameterF1-19.032.33F22-3.3813.97F2-40.36-20.45F23-5.66-9.55F3-20.24-4.91F24-55.85-37.15F418.798.71F250.50-9.92F5-26.15-1.94F263.26-4.12F686.9916.27F2722.2516.38F7-3.0311.77F2822.7713.75F8-1.272.20F29-7.10-6.41F92.502.92F30-41.68-9.68F10-9.512.45F312.32-13.64F1155.8226.50F3235.0629.88F12-44.50-45.84F33-1.33-18.58F137.04-3.27F346.47-2.78F142.14-2.00F35-32.23-29.33F15-10.52-3.23F36-10.2934.45F16-16.15-12.54F374.9611.36F1713.585.92F38-39.19-18.34F183.9115.11F39-9.93-11.10F197.251.37F40-19.00-23.56F2040.3712.83F411.7414.81F2117.2618.52F4261.4326.83

图1 四川桤木杂交子代各性状的遗传力Fig.1 Heritability of hybrid offspring traits in A.cremastogyne Burk.

图2 四川桤木杂交子代苗高和地径之间的相关分析Fig.2 Correlation analysis between height and diameter of hybrid offspring of A.cremastogyne
3 结论与讨论
测定1 a生四川桤木杂交苗的苗高和地径两个性状并进行了遗传参数的估算,目的是为了研究四川桤木杂交子代的遗传规律,为优良家系/无性系的选育创造条件[12]。四川桤木杂交子代的苗高和地径在家系间存在极显著差异,家系内存在显著差异,可以进行家系间和家系内的初步选择,为进一步开展四川桤木遗传改良提供种质材料。母本×父本在苗高上的方差达到显著水平,而地径的方差不显著,表明母本×父本互作对苗高的影响更为重要[13]。四川桤木同一亲本不同性状及同一性状不同亲本间,一般配合力(GCA)效应存在明显差异,表明这些亲本的遗传特性均存在差异,即加性效应大小是不同的[14]。JT4、JG6和BZ6作母本的苗高和地径一般配合力(GCA)均为正,LD8和XH5作父本的苗高和地径一般配合力(GCA)均为正。多重比较结果表明,F6、F32、F36和F42等以JT4、BZ6和JG6为母本,XH5和LD8为父本的杂交子代均具有较高的苗高和地径生长量,因此,初步认为这5个亲本为优良的杂交育种亲本资源。不同家系不同性状的特殊配合力(SCA)效应值差异显著,其中家系F6(JT4×YO2)、F32(BZ6×JT2)、F36(JT4×JT2)和F42(JT4×XH5)的苗高和地径特殊配合力相对较高,利用这4个组合进行杂交,可获得优良杂交子代。
遗传力是表型变异受遗传因素影响的比例,是杂交后代选择的一个重要指标[14]。魏永成等[15]对杜仲杂交子代进行遗传分析发现,2 a生时各性状均具有较高的广义遗传力,苗期选择具有较大的遗传潜力。杜克兵等[11]对黑杨派杨树杂交子代进行遗传分析发现,苗高和地径具有较高的广义遗传力,与本研究结果类似,说明在林木遗传改良中,通过苗期选择,可以使杂交子代获得显著的遗传增益。遗传力分析结果显示,四川桤木杂交子代的苗高和地径均具有较高的广义遗传力,其中苗高的遗传力在50%以上,表明亲本基因型对子代有很大的影响。林木表型性状间的相关系数反映了性状间的相关程度,可为林木遗传育种方案和改良策略的制定提供理论参考[16]。四川桤木杂交子代苗高和地径呈显著正相关,可根据苗高和地径开展联合选择,以选育优良家系/无性系,提高育种效率。长期以来,育种工作者都将F1代的选择作为育种主体,利用多群体、多世代改良提高杂交育种的预见性和效率[17]。根据NCⅡ交配设计获得的杂交子代一般配合力和特殊配合力较高,可以在大范围内进行选择,在以后的工作中,可进一步扩大群体,进行F1代的选择。
由于只是基于1 a生四川桤木杂交子代苗期表型性状的调查分析,考虑到早期性状的可靠性问题,因此,在四川桤木后续遗传改良研究中,应进一步结合分子标记和连续多年观测数据进行分析,为四川桤木杂种优势的研究与利用提供更加详细、可靠的依据。
[1] 杨志成.优良阔叶树种—桤木的分布、生长和利用[J].林业科学研究,1991,4(6):643~648.
[2] 王军辉,顾万春,李斌,等.桤木优良种源/家系的选择研究—生长的适应性和遗传稳定性分析[J].林业科学,2000,36(3):59~66.
[3] 周永丽,刘福云,万军,等.四川桤木木材材性初步研究[J].四川林业科技,2003,24(1):75~78.
[4] 陈益泰,李桂英,王惠雄.桤木自然分布区内表型变异的研究[J].林业科学研究,1999,12(4):379~385.
[5] 王军辉,顾万春,夏良放,等.桤木种源(群体)/家系材性性状的遗传变异[J].林业科学研究,2001,14(4):362~368.
[6] 卓仁英,孟现东,陈益泰.桤木群体遗传分化研究Ⅰ.DNA提取和PCR条件的建立[J].林业科学研究,2003,16(1):117~122.
[7] 任保青,刘军.中国桤木属植物的细胞学研究(Ⅰ)[J].广西植物,2006,26(4):356~359.
[8] 杨汉波,饶龙兵,郭洪英,等.5种桤木属植物的核型分析[J].植物遗传资源学报,2013,14(6):1203~1207.
[9] 杜克兵,许林,沈宝仙,等.黑杨派杨树杂交子代的遗传分析及苗期选择[J].华中农业大学学报,2009,28(5):624~630.
[10] 王庆菊,李晓磊,王磊,等.紫叶稠李叶片花色苷及其合成相关酶动态[J].林业科学,2008,44(3):45~49.
[11] Chung Y C,Kai W Y.Differential expression of MYB gene (OgMYB1) determines color patterning in floral tissue of Oncidium G ower Ramsey [J].Plant Mol Biol,2008,66:379~388.
[12] 周素华,李周岐,李煜.杜仲优树自由授粉家系遗传变异及苗期初选[J].西北林学院学报,2010,25(1):57~60.
[13] 何旭东.桉树杂种优势及其分子标记辅助选择研究[D].南京:南京林业大学,2010.
[14] 李周岐,王章荣.鹅掌楸属种间杂种苗期生长性状的遗传变异与优良遗传型选择[J].西北林学院学报,2001,16(2):5~9.
[15] 魏永成,李周岐,李煜.杜仲杂交子代苗期表型性状的遗传分析[J].西北农林科技大学学报(自然科学版),2012,40(8):137~143.
[16] 李守勇,赵永军,孙明高.11个黑杨无性系叶片性状变异研究[J].山东林业科技,2002,2:5~7.
[17] 栾启福.几种松树杂交选育及其遗传分析[D].北京:中国林业科学研究院,2010.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
农技服务(2020年1期)2020-12-17
贵州林业科技(2020年2期)2020-09-07
山东农业科学(2019年11期)2019-12-24
现代农业科技(2019年1期)2019-07-11
中国种业(2018年11期)2018-11-16
猪业科学(2018年5期)2018-07-17
现代农业科技(2017年24期)2018-01-22
中南林业科技大学学报(2016年1期)2016-12-20
广西林业科学(2016年3期)2016-03-16
