
喀斯特峰丛洼地不同植被恢复阶段优势种根系构型特征
2018-04-08 02:24曾馥平宋同清彭晚霞
西北植物学报 2018年1期
苏 樑,杜 虎 ,王 华,曾馥平,宋同清*,彭晚霞,陈 莉,张 芳
(1 中国科学院亚热带农业生态研究所 亚热带农业生态过程重点实验室, 长沙410125;2 湖南农业大学 生物科学技术学院, 长沙410128; 3 中国科学院环江喀斯特生态系统观测研究站, 广西环江547100)
在陆地生态系统中,根系作为植物吸收和运输土壤养分的重要器官,对植物的生长和发育起到了非常重要的作用[1]。植物在土壤中利用其庞大的根系来固定土壤颗粒,以此来很好地固定和支持植物体,还可以有效地控制土壤侵蚀的发生,发挥了重要的水土保持功能[2]。同时,根系也在植物与土壤环境进行物质和能量交换的过程中起了最主要的作用,其分布特点反映了植物对土壤资源的利用机制及其对环境的适应性[3]。对于根系构型特征的研究主要着力于根系几何形态和拓扑结构参数,几何形态参数主要包括根长、根直径、根系生物量和分支角度等;而根系的分支状况主要是通过连接数量、根系分支率以及根系在土层中的空间分布来反映的[4]。近年来,根系拓扑结构在根系构型特征中已经受到越来越多的重视[5]。国外一些科学家提出根系拓扑结构有鱼尾形分支和叉状分支模式两种极端类型,并且应用拓扑指数来表示不同植物根系的分支模式,而且比较了不同的分支模式的营养竞争能力,发现鱼尾形分支对贫瘠的生境更适应[6-7]。也有人提出根系拓扑结构会受到自然生境条件改变的影响,进而促使根系的分支和扩展策略发生适应性变化。自然资源具有时空分布的异质性,根系的形态特征受到植物对土壤资源吸收能力影响,而根系构型会影响到植物地上部分的生长发育以及生态功能的发挥[8]。大多数研究表明,根系分支的横截面积在分支前后均相等,但在适应性环境中根系构型如何变化还没有一致的结论[9-10]。
喀斯特峰丛洼地是亚热带喀斯特地区最重要的景观类型,具有岩石裸露率和钙镁含量高、土壤瘠薄、季节性岩溶干旱严重的环境特征,其适生植物具有石生性、嗜钙性、耐瘠性和耐旱性等限制性特点[11]。而在不同的人为干扰强度下,植被形成了原生林、次生林、灌丛等3个不同的演替阶段[12-13]。喀斯特地区的石漠化和严重的生态退化一直以来是植被恢复和生态重建的难点地区,以往对喀斯特植被的研究主要集中在群落结构、生物多样性及其与土壤养分、水分、地形的相互关系等方面[14-18],而对地下根系构型的研究甚少。因此,本研究通过考察灌丛、次生林、原生林3种植被恢复阶段的优势种根系的拓扑结构特征,比较在喀斯特峰丛洼地特殊生境条件下,植物根系构型的差异性和相似性,探讨资源合理高效的利用方式,为构建喀斯特复式镶嵌群落提供理论和科学依据,促进喀斯特地区植被迅速恢复和生态重建。
1 研究地区与研究方法
1.1 研究区概况
本研究区位于广西环江毛南族自治县(107°51′~108°43′E、24°44′~25°33′N),属亚热带季风气候区,最高海拔为1 028.0 m。根据广西环江县气象局1986~2005年观测的20年的气象数据,研究区年平均气温为15.7 ℃,1月平均气温为10.1℃,7月平均气温为28 ℃,历年最低气温为-5.2 ℃,无霜期为290 d,年平均日照时数为1 451 h,年降水量为1 389.1 mm,4~9月降水量占全年降水量的70%,平均蒸发量为1 571.1 mm,平均相对湿度为70%。
喀斯特峰丛洼地集中分布在该县的西南部,土壤以碳酸盐岩发育的深色或棕色石灰土为主,土层浅薄,坡度大,水土流失严重,岩石裸露情况严重,石漠化趋势严峻。研究区主要有草丛、灌丛、次生林和原生林(顶极群落)4种植被类型。草丛主要有以斑茅(Saccharumarundinaceum)、白茅(Imperatacylindrica)、蔓生莠竹(Microstegiumfasciculatum)、纤毛鸭嘴草(Ischaemumciliare)、荩草(Arthraxonhispidus)和鬼针草(Bidenspilosa)为主要建群种的群落类型;灌丛主要有以长序苎麻(Boehmeriadolichostachya)、黄荆(Vitexnegundo)、红背山麻杆(Alchorneatrewioides)、毛桐(Mallotusbarbatus)和火棘(Pyracanthafortuneana)为主要建群种的群落类型;次生林主要有以八角枫(Alangiumchinense)、盐肤木(Rhuschinensis)和刀果鞍叶羊蹄甲(Bauhiniabrachycarpa)为主要建群种的群落类型;原生林主要有以伊桐(Itoaorientalis)、掌叶木(Handeliodendronbodinieri)、菜豆树(Radermacherasinica)、灰岩棒柄花(Cleidionbracteosum)、青冈(Cyclobalanopsisglauca)、南酸枣(Choerospondiasaxillaris)、刨花润楠(Machiluspauhoi)、伞花木(Eurycorymbuscavaleriei)、厚壳桂(Cryptocaryachinensis)、化香树(Platycaryastrobilacea)、光叶槭(Acerlaevigatum)、侧柏(Platycladusorientalis)、乌冈栎(Quercusphillyraeoides)、铁榄(Sinosideroxylonpedunculatum)、翠柏(Calocedrusmacrolepis)和罗城鹅耳枥(Carpinusluochengensis)为主要建群种的群落类型[19]。
1.2 根系挖掘方法
选取灌丛、次生林、原生林3种植被类型中4种有代表性的建群种,灌丛为红背山麻秆、黄荆、火棘、毛桐,次生林为菜豆树、野桐(Mallotustenuifolius)、灰毛浆果楝(Cipadessacinerascens)、八角枫(Alangiumchinense),原生林为圆果化香树(PlatycaryalongipesWu)、青冈(Cyclobalanopsisglauca)、枫香树(Liquidambarformosana)、栀子皮(Itoaorientalis),每种3株,测定基径和冠幅,采用全挖法,用铁铲、锄头、撬棍等工具小心挖取,清理出所有直径大于2 mm的根系(小于2 mm的根系在挖掘中易断,其根系构型的重建难以实现),尽量保持根系在土壤中的分布位置,采用50 cm×50 cm的坐标纸确定根系的分布位置(直到没有根系出现为止),按照1∶50的比例在坐标纸上画出根系的构型图。
1.3 根系参数测定与数据处理
将根系完全挖出后,用游标卡尺和卷尺等测量工具测定各级根长度、分支前后直径(测定所有根系分支,同一分支部位测量2次)、各级根数目、生物量、根系分支数目、根系内部/外部连接数目、连接长度等。实验结束后,所有的数据都采用SPSS 22.0软件的ANOVA方法进行方差分析,用Tukey HSD(α=0.01)法进行显著性检验,采用Microsoft Excel2007、OriginPro8.0进行数据计算处理和绘图。
1.4 理论分析方法
(1)拓扑指数(TI):最早提出通过拓扑指数来研究植物根系分支模式的是Fitter等,且提出鱼尾形分支和叉状形分支两种极端类型(图1)。
通过拓扑指数对根系构型进行分析,拓扑指数TI= lgA/lgM(M为根系所有外部连接总和, A为最长根系通道内部连接的总数),鱼尾形分支TI=1,TI越接近0.5根系就越接近叉状分支。由于当Fitter拓扑模型中典型叉状分支结构的拓扑指数TI接近于0.5时,大小是决定于M的[7],因此Oppelt等[20]对拓扑模型进行了修正,提出了修正的拓扑指数模型:
(1)
其中,a为拓扑长度(植物基部到根终端连接数量),b为平均拓扑长度,Ibv0=lnv0/ln2,b=Pe/v0(v0相当于Fitter拓扑模型中的M,Pe为根系基部到根系终端通道的所有连接总数)。修正后的拓扑参数qa、qb值的范围在0~1之间,其中鱼尾形分支qa=qb=1,叉状分支qa=qb=0,qa、qb值在0~1之间表示两种典型分支模式的过渡形式(表1)。
(2)根系分支率:根序依据Strahler[21]和Berntson[22]的方法确定,最末端的小根为一级根,两个一级根汇合为二级根,两个二级根汇合为三级根,依此类推,如果不同根级相遇,则取较高的作为根级。计算每个等级(i)的根系数量(Ni),以等级(i)为横坐标、lgNi为纵坐标作图,取回归直线斜率绝对值的逆对数作为根系的总分支率(Rb);逐步分支率(Ri/Ri+1)为相邻两级根系分支数之比,计算公式为:Ri/Ri+1=Ni/Ni+1。
Leonardo da Vinci法则认为植物分支前后的根系横截面积相等,即分支前的根系横截面积等于分支后的根系横截面积总和,用方程式(2)表示:
(2)
通过方程式(2),van Noordwijk和Purnomosidhi[23]提出了能够反映普遍根系分支规律的方程(3):
(3)

表1 两类典型分支的拓扑参数
其中,α为根系分支前的横截面积与分支后的面积总和之比,d前为根系分支前的直径,di为根系分支后第i级分支的直径。
2 结果与分析
2.1 各植被恢复阶段优势种根系拓扑指数与拓扑结构
由表2可知,灌丛、次生林、原生林恢复阶段优势种根系拓扑指数TI接近0.5,说明3种植被恢复阶段优势种根系的分布较为复杂,次级分支多,为典型的叉状分支;其中,拓扑指数TI又表现为次生林>原生林>灌丛,说明次生林分支结构更为复杂,次级分支相对较多。进一步通过单因素方差分析发现,3种植被恢复阶段的根系拓扑指数TI差异不显著,又说明3种植被恢复阶段植被有趋同性。
2.2 各植被恢复阶段优势种根系连接长度
3种植被恢复阶段优势种的根系平均连接长度均在34.29 cm以上,且平均值为37.01 cm。从图2可以看出,外部连接长度大于平均根系连接长度,3种植被恢复阶段表现为灌丛>原生林>次生林。较长的连接长度可以降低根系自身对土壤养分的竞争,同时也可以帮助植物拓展空间。但是,3种植被类型的根系外部根系连接长度和平均根系连接长度均不存显著性差异,说明在喀斯特峰丛洼地浅薄贫瘠的土壤中,3种植被类型均采取增加根系连接长度的策略来寻求更多的水分和养分,维持自身的生长与发育。
2.3 各植被恢复阶段优势种根系分支率
根系分支的差异反映了根系适应环境的能力。根据表3可知,3种植被类型的根系总分支率Rb和逐步分支率均不存在显著差异,但总分支率Rb表现为灌丛(3.68±0.50)>原生林(3.10±0.32)>次生林(3.43±0.40),即灌丛逐步分支率表现为增长的趋势,原生林逐步分支率表现为减少的趋势,且在次生林中最小。
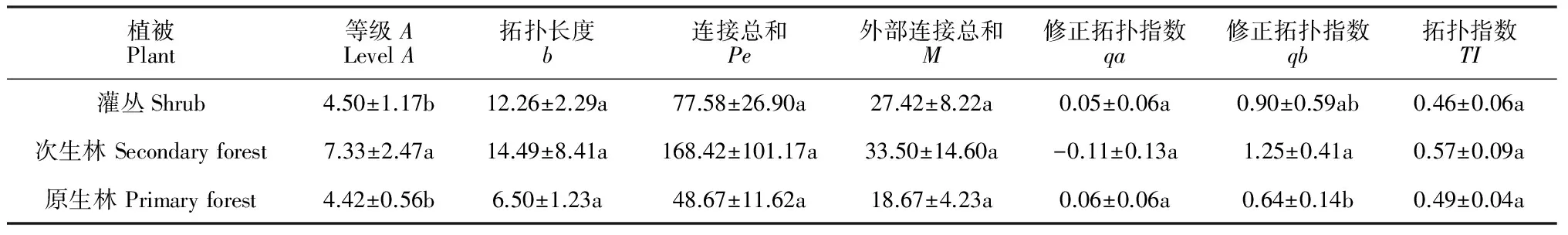
表2 3种植被恢复阶段优势种根系拓扑结构参数
注:同列不同字母表示差异显著(P<0.05). 下同
Note: Different letters in the same column meant significant difference at 0.05 level.The same as below
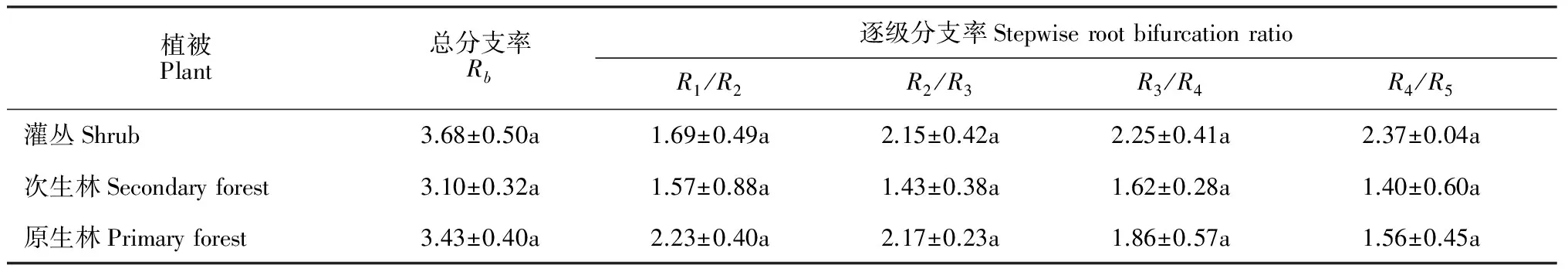
表3 3种植被恢复阶段优势种根系总分支率(Rb)和逐级分支率(Ri /Ri+1)
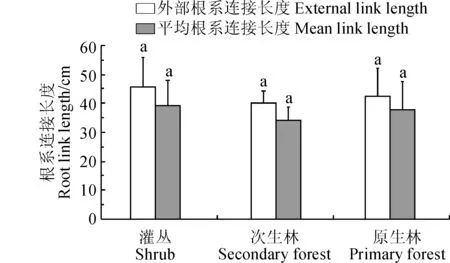
图2 3种植被恢复阶段优势种根系连接长度Fig.2 The lengths of root system of the dominant species in three vegetation restoration processes

图3 3种植被恢复阶段优势种根系直径缩减速率Fig.3 The relative tapering of the coarse root of the dominant species in three vegetation succession stages
2.4 各植被恢复阶段优势种根系直径缩减速率
根据图3可以看出,3种植被类型根系直径缩减速率与距离根茎的连接通道长度均呈一致的变化趋势,表现为随着距离主根的连接长度的增加,根系直径的缩减速率迅速减小,且在急剧减小后趋于稳定,整体呈原生林>灌丛>次生林的趋势。
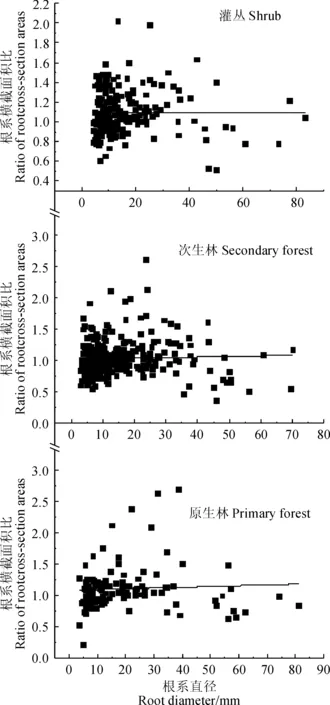
采用线性方程对根系横截面积与根系直径间的拟合图4 3种植被恢复阶段优势种根系分支横截面积比Relationships between ratio of root cross-sectional areas and root diameter were fitted with a linear model Fig.4 Ratio of root cross-sectional areas of the dominant species in three vegetation restoration processes
2.5 根系分支前后横截面积的关系
如图4所示,3种植被恢复阶段优势种的横截面积比值α分别为1.072、1.039、1.115,基本符合Leonardo da Vinci法则,说明3种植被类型分支前后的根系横截面积基本相等;同时,根系分支前后的根系横截面积之比与根系直径的相关关系不显著,又说明根系横截面积相等是一般规律,不随直径的变化而产生变化。
3 讨 论
根系统拓扑分析的主要目的是探索根系统在生境变化的条件下,是否会出现分支和扩展策略的变化[8]。拓扑结构特征是根系构型的重要组成部分,决定了根系在土壤空间中的分布,也影响根系吸收水分和养分的能力[23]。有研究者提出了根系拓扑结构的鱼尾形分支和叉状分支模式2种极端类型,并且应用拓扑指数来表示不同植物根系的分支模式[3,6-7]。在本研究中,灌丛、次生林、原生林3种植被恢复阶段优势种拓扑指数TI接近0.5,并表现为次生林>原生林>灌丛,它们的根系分支模式为典型的叉状分支。可见,在喀斯特峰丛洼地区域,无论是灌丛还是乔木,根系都呈叉状分布。这是因为喀斯特峰丛洼地土壤造壤能力差,土层浅薄且不连续,土壤养分含量处于较低的水平[24-28],导致根系分支结构复杂,次级分支较多,故而植物根系主要采取水平生长策略,能够迅速占据大量土壤空间,保障其在土壤中吸收和竞争水分和营养的能力。
植物根系的长度直接影响根在土壤中扩张空间和吸收营养的能力,根系统连接的长度越长,其空间的能力就越强[29]。在本研究喀斯特峰丛洼地3种植被恢复类型中,根系外部连接长度均大于平均根系连接长度,可知外部连接长度大于内部连接长度,说明在喀斯特峰丛洼地中灌丛与乔木的根系均强调向外扩张的生长策略,这与杨小林等[9]、郭京衡等[10]在塔克拉玛干沙漠地区研究的结果一致,证明根系外部连接长度大于内部连接长度是根系生长的普遍策略。同时,本研究中根系连接平均长度在灌丛(39.05 cm)、次生林(34.30 cm)和原生林(37.68 cm)间差异不显著,又表明在喀斯特峰丛洼地土壤养分水平较低的环境下,3种植被类型均通过增加连接长度来扩展空间,从而使自身能够得到所需的水分和养分供应,这在 “根包石”这种需要根系沿着岩石向下扩张直至土壤的小生境中体现得极为明显。
根系分支率是植物构型研究中的一个重要指标,根系根分支率的变化直接反映了根系统适应环境的能力[30-31]。总分支率相对于沙生植物[9-10]来说较小,这是因为在喀斯特地区土层薄的环境下,根系只能在水平角度通过较长连接长度和较少的分支来扩展土壤空间。同时,喀斯特峰丛洼地不同恢复阶段植被根系总分支率和逐步分支率差异均不显著,进一步说明不同植被恢复阶段优势种在喀斯特极具异质性的土壤环境中,根系的生态适应性是相同的。
根系的直径缩减速率反映了植物的觅养对策,有研究表明,植物根系的直径缩减速率与其离主根的距离呈反比[32],本研究结果与其是一致的。本研究表明,喀斯特峰丛洼地原生林的整体根系直径缩减速率最大,同时外部连接长度和其他植被类型无显著变化,说明原生林的主根较粗,这是由于原生林植物物种丰富,土壤养分竞争激烈,植物根系的生长空间受到限制,需要在地形环境更恶劣的如岩石等立地条件下生长。Leonardo da Vinci法则认为植物根系分支前后的横截面积是相等的,即分支前的根系横截面积等于分支后的根系横截面积总和[23,33],本研究发现喀斯特峰丛洼地不同植被恢复阶段灌丛、次生林、原生林的根系分支率分别为1.072、1.039、1.115,根系分支前后遵循Leonardo da Vinci法则,且根系分支前后直径之比不随着直径的变化而变化,这与van Noordwijk和Purnomosidhi及Eduardo等的研究结果一致[23,33],说明根系分支前后横截面积的相等性是植物根系构型的一般性特征,不因根系直径的变化而改变。
另外,本研究也存在一些不足之处,由于喀斯特地区岩石裸露率大,部分样株的主根在石缝中难以挖取,采用根系缩减速率进行的估算可能会出现结果与真实值的小范围偏差。由于喀斯特峰丛洼地存在许多不同的复杂小生境,而对于此类生境还不完全了解,这是以后喀斯特地区根系构型研究的重要方向。
参考文献:
[1]DANNOWSKI M, BLOCK A. Fractal geometry and root system structures of heterogeneous plant communities[J].PlantandSoil, 2005,272(1): 61-76.
[2]周本智,张守攻,傅懋毅.植物根系研究新技术Minirhizotron的起源、发展和应用[J].生态学杂志,2007,26(2):253-260.
ZHOU B Z, ZHANG S G, FU M Y. Minirhizotron, a new technique for plant root system research: its invention, development and application[J].ChineseJournalofEcology,2007,26(2):253-260.
[3]FITTER A H,STICKLAND T R,HARVEY M L,etal. Architectural analysis of plant root systems 1. Architectural correlates of exploitation efficiency[J].NewPhytologist, 1991,118(3): 375-382.
[5]HODGE A,BERTA G,DOUSSAN C,etal. Plant root growth, architecture and function[J].PlantandSoil, 2009,321(1): 153-187.
[6]FITTER A H. An Architectural approach to the comparative ecology of plant root systems[J].NewPhytologist, 1987,106(106): 61-77.
[7]FITTER AH,STICKLABD TR. Architectural analysis ofplant root systems.2. Influence of nutrient supply on architectural contrasting plant species[J].NewPhytologist, 1991, 118: 383-389.
[8]OPPELT A L,KURTH W,GODBOLD D L. Topology,scaling relations and Leonardo’s rule in root systems from African tree species[J].TreePhysiology, 2001,21(2-3): 117-128.
[9]杨小林, 张希明, 李义玲,等. 基于分形理论的塔克拉玛干沙漠腹地自然植物根系构型特征分析[J]. 干旱区资源与环境, 2015,29(8):145-150.
YANG X L, ZHANG X M, LI Y L,etal. Analysis and estimation of root architecture in the hinterland of Taklimakan desert base on the fractal theory [J].JournalofAridLandResourcesandEnvironment, 2015,29(8): 145-150.
[10]郭京衡,曾凡江,李尝君,等. 塔克拉玛干沙漠南缘三种防护林植物根系构型及其生态适应策略[J]. 植物生态学报,2014,38(1):36-44.
GUO J H, ZENG F J, LI C J,etal. Root architecture and ecological adaptation strategies in three shelterbelt plant species in the southern Taklimakan Desert[J].ChineseJournalofPlantEcology, 2014,38(1): 36-44.
[11]袁道先. 现代岩溶学和全球变化研究[J]. 地学前缘, 1997,(z1):17-25.
YUAN D X. Modern karstology and global change study[J].EarthScienceFrontiers, 1997, (z1): 17-25.
[12]曾馥平,彭晚霞,宋同清,等. 桂西北喀斯特人为干扰区植被自然恢复22年后群落特征[J]. 生态学报,2007,27(12):5 110-5 119.
ZENG F P, PENG W X , SONG T Q,etal. Changes in vegetation after 22 years’ natural restoration in the karst disturbed area in Northwest Guangxi[J].ActaEcologicaSinica, 2007,27(12): 5 110-5 119.
[13]彭晚霞,王克林,宋同清,等. 喀斯特脆弱生态系统复合退化控制与重建模式[J]. 生态学报,2008,28(2):811-820.
PENG W X, WANG K L, SONG T Q,etal. Controlling and restoration models of complex degradation of vulnerable Karst ecosystem[J].ActaEcologicaSinica, 2008,28(2): 811-820.
[14]宋同清,彭晚霞,曾馥平,等. 桂西北喀斯特人为干扰区植被的演替规律与更新策略[J]. 山地学报,2008,26(5):597-604.
SONG T Q, PENG W X, ZENG F P,etal. Vegetation Succession Rule Andregeneration Stratigies in Disturbed Karst Area, Northwest Guangxi[J].JournalofMountainScience, 2008,26(5): 597-604.
[15]宋同清,彭晚霞,曾馥平,等. 喀斯特峰丛洼地不同类型森林群落的组成与生物多样性特征[J]. 生物多样性,2010,18(4):355-364.
SONG T Q, PENG W X, ZENG F P,etal. Community composition and biodiversity characteristics of forests in Karst cluster-peak-depression region[J].BiodiversityScience, 2010,18(4): 355-364.
[16]宋同清,彭晚霞,曾馥平,等. 木论喀斯特峰丛洼地森林群落空间格局及环境解释[J]. 植物生态学报,2010,34(3):298-308.
SONG T Q, PENG W X, ZENG F P,etal. Spatial pattern of forest communities and environmental interpretation in Mulun National Nature Reserve, karst cluster-peak depression region[J].ChineseJournalofPlantEcology, 2010,34(3): 298-308.
[17]彭晚霞,宋同清,曾馥平,等. 喀斯特常绿落叶阔叶混交林植物与土壤地形因子的耦合关系[J]. 生态学报,2010,30(13):3 472-3 481.
PENG W X, SONG T Q, ZENG F P,etal. The coupling relationships between vegetation, soil, and topography factors in karst mixed evergreen and deciduous broadleaf forest[J].ActaEcologicaSinica, 2010,30(13): 3 472-3 481.
[18]杜虎,彭晚霞,宋同清,等. 桂北喀斯特峰丛洼地植物群落特征及其与土壤的耦合关系[J]. 植物生态学报,2013,37(3):197-208.
DU H, PENG W X, SONG T Q,etal. Plant community characteristics and its coupling relationships with soil in depressions between karst hills, North Guangxi, China[J].ChineseJournalofPlantEcology, 2013,37(3): 197-208.
[19]杜虎, 宋同清, 曾馥平,等. 喀斯特峰丛洼地不同植被类型碳格局变化及影响因子[J]. 生态学报, 2015,35(14):4 658-4 667.
DU H, SONG T Q, ZENG F P,etal. Carbon storage and its controlling factors under different vegetation types in depressions between karst hills, southwest China[J].ChineseJournalofPlantEcology, 2015,35(14): 4 658-4 667.
[20]OPPELT A L,KURTH W,JENTSCHKE G,etal. Contrasting rooting patterns of some arid-zone fruit tree species from Botswana - I. Fine root distribution[J].AgroforestrySystems, 2005,64(1): 1-11.
[21]STRAHLER A N. Hypsometric (area-altitude) analysis of erosional topography[J].GeologicalSocietyofAmericaBulletin, 1952,63(11): 1 117.
[22]BERNTSON G M. The Characterization of Topology: A Comparison of Four topological Indices for Rooted Binary Trees[J].JournalofTheoreticalBiology, 1995,177(3): 271-281.
[23]NOORDWIJK M V, PURNOMOSIDHI P. Root Architecture in Relation to Tree-Soil-Crop Interactions and Shoot Pruning in Agroforestry[M]// Agroforestry: Science,Policy and Practice. Springer Netherlands, 1995: 161-173.
[24]BIONDINI, MARIO E, GRYGIEL,etal. Landscape distribution of organisms and the scaling of soil resources[J].TheAmericanNaturalist, 1994,143(6): 1 026-1 054.
[25]王克林,苏以荣,曾馥平,等. 西南喀斯特典型生态系统土壤特征与植被适应性恢复研究[J]. 农业现代化研究,2008,29(6):641-645.
WANG K L, SU Y R, ZENG F P,etal. Ecological process and vegetation restoration in Karst Region of southwest China[J].ResearchofAgriculturalModernization, 2008,29(6): 641-645.
[26]彭晚霞,宋同清,曾馥平,等. 喀斯特峰丛洼地旱季土壤水分的空间变化及主要影响因子[J]. 生态学报,2010,30(24):6 787-6 797.
PENG W X, SONG T Q, ZENG F P,etal. Spatial variation of soil water and the key impact factors in dry season in Karst cluster-peak-depression region[J].ActaEcologicaSinica, 2010,30(24): 6 787-6 797.
[27]宋同清,彭晚霞,曾馥平,等. 喀斯特木论自然保护区旱季土壤水分的空间异质性[J]. 应用生态学报,2009,20(1):98-104.
SONG T Q, PENG W X, ZENG F P,etal. Spatial heterogeneity of surface soil moisture content in dry season in Mulun National Natu-ral Reserve in Karst Area[J].JournalofAppliedEcology, 2009,20(1): 98-104.
[28]刘淑娟,张伟,王克林,等. 桂西北喀斯特峰丛洼地表层土壤养分时空分异特征[J]. 生态学报,2011,31(11):3 036-3 043.
LIU S J, ZHANG W, WANG K L,etal. Spatiotemporal heterogeneity of topsoil nutrients in Karst Peak-Cluster depression area of Northwest Guangxi, China[J].ActaEcologicaSinica, 2011,31(11): 3 036-3 043.
[29]WALK T C,VAN ERP E,LYNCH J P. Modelling applicability of fractal analysis to efficiency of soil exploration by roots[J].AnnalsofBotany, 2004,94(1): 119.
[30]孙书存,陈灵芝. 辽东栎植冠的构型分析[J]. 植物生态学报,1999,23(5):433-440.
SUN S C, CHEN L Z. Architectural analysis of crown geometry inQuercusliaotungensis[J].ChineseJournalofPlantEcology, 1999,23(5): 433-440.
[31]单立山,李毅,董秋莲,等. 红砂根系构型对干旱的生态适应[J]. 中国沙漠,2012,32(5):1 283-1 290.
SHAN L S, LI Y, DONG Q L,etal. Ecological adaptation ofReaumuriasoongoricaroot system architecture to arid environment[J].JournalofDesertResearch, 2012,32(5): 1 283-1 290.
[32]KALLIOKOSKI T, NYGREN P, SIEVNEN R. Coarse root architecture of three boreal tree species growing in mixed stands[J].SilvaFennica, 2008,42(2): 189-210.
[33]SALAS E, OZIERLAFONTAINE H, NYGREN P. A fractal root model applied for estimating the root biomass and architecture in two tropical legume tree species[J].AnnalsofForestScience, 2004,61(4): 337-345.
猜你喜欢
现代农村科技(2022年5期)2022-11-18
特区文学·诗(2021年6期)2021-12-22
乡村地理(2019年2期)2019-11-16
建材发展导向(2019年5期)2019-09-09
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26
知识经济·中国直销(2018年10期)2018-11-06
现代农业研究(2017年11期)2018-01-12
现代园艺(2017年20期)2017-02-03
农业与技术(2016年6期)2016-05-14
遥感信息(2015年3期)2015-12-13
