
不同光照强度对火力楠幼苗生长及生理特性的影响
2018-05-16 09:53韦中绵覃德文吴敏秦武明
西部林业科学 2018年2期
韦中绵,覃德文,吴敏,秦武明
(1.南宁树木园,广西 南宁530031;2.广西大学 林学院,广西 南宁 530004)
光照强度作为影响植物生长发育的重要指标,在植物生长发育过程中显著影响着植物形态形成和光合作用[1]。特别是在植物幼苗期对光照强度的要求更高,适当的光照强度可有效地提升植物幼苗叶片的生长、根系的形成,且获得高效的光合产物[2]。近年来,随着园林花卉市场的快速发展,以及珍贵树种苗木需求量增长,提升植物苗期生长、增加苗木产量成为广大科研工作者研究的热点,然而光照强度作为影响植物幼苗最直接的因素,如何控制光照强度,寻找适合的光照条件,成为了植物培育、营林生产的重要问题。
火力楠(Micheliamacclurei),学名醉香含笑,为木兰科(Magnoliaceae)含笑属(Michelia)乔木树种,是广西主要的乡土种植树种之一。在营林生产中,火力楠作为珍贵乡土树种进行培育以获取优质的家具用材[3]。在园林栽培中,火力楠由于树形笔直优美,花朵艳丽芳香,成为园林花卉设计必备树种[4]。随着其苗木需求量的逐年增长,广大园林和林业工作者对火力楠苗木开展了大量科学研究。其中,彭玉华等[5]对火力楠容器苗进行了缓释肥和复合肥试验,总结得出了在基质中添加0.6%缓释肥能促进火力楠苗木快速生长;陈宇超等[6]通过不同生长调节剂配比对火力楠苗木展开研究,得出了生长调节剂NAA浓度为100mg/L对火力楠的生长有着较强的促进作用;欧斌等[7]对赣南地区火力楠苗木展开质量分级评定,提出以苗高和地径作为该树种苗木分级的质量指标,并将其分为3级分级标准。但对于不同光照强度对火力楠苗木生长影响的研究未见报道。因此,本文设置了4个不同的光照强度梯度,对火力楠苗木进行生长和生理生化研究,探讨出适合火力楠苗木生长的最佳光照强度,为火力楠苗木栽培和营林生产提供科学的理论依据。
1 材料与方法
1.1 试验地概况及材料来源
试验地位于广西南宁市广西大学林学院苗圃实验基地内(22°51′18″N,108°17′21″E),选用广西南宁市树木园一年生优质苗木进行光照试验,试验初期以黄心土和砂土以3︰1比例作为土壤基质进行育苗,每月定期施肥,待培育3个月后,选取生长指标(苗高、地径)一致的苗木进行试验。
1.2 试验方法
1.2.1 试验方法
试验于2016年5月1日进行,对苗木进行遮荫网遮荫处理,并设置3个光照强度梯度,分别为:1层遮荫P1(42.3%NS透光率)、2层遮荫P2(12.6%NS透光率)、3层遮荫P3(5.9%NS透光率)以及全光照(对照)CK(100%NS透光率),4个光照强度试验。其中每个光照处理选取生长状况一致苗木(苗高40-45cm,地径1.5-2.5cm)做3个重复,每个重复4株,共12株。试验进行至2016年11月1日,共6个月时间,在试验期间对每个处理苗木进行相同的日常管护。
1.2.2 指标测定
(1)生长指标测定 在试验进行期间,每隔30d对苗木进行生长指标测定,运用测量尺测定苗木苗高增长量;游标卡尺测定苗木地径生长量;选取由上到下螺旋选取5片长势良好、无病害的叶片,运用叶面积仪-YMJ-A测定苗木叶面积,并计算叶面积生长量。
(2)叶片光合指数测定 待光照试验进行至2016年11月1日,选取光照充足的天气,于早晨9:00-12:00分别选取各处理的植株顶端完全展开的6片功能叶进行光合测定,采用LI-6400XT便携式光合作用测量系统(IRGA,LI-COR,Lincoln,USA)测定叶片光响应曲线、净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)等光合指标。
(3)生理生化指标测定 在光照试验结束后,于2016年11月15日每个处理选取6株苗木进行叶片生理生化指标测定,每株苗木筛选由上到下6片长势良好、健康叶片带回实验室测定叶绿素含量、丙二醛含量(MDA)、超氧化物歧化酶活性(SOD)、过氧化氢酶活性(CAT)等。其中叶绿素含量测定采用丙酮提取法[8],采用硫代巴比妥酸(TBA)显色法测定MDA[9],采用氮蓝四唑(NBT)光化还原法测定SOD[10],采用紫外吸收法测定CAT[11]。
2 结果与分析
2.1 不同光照对苗木生长指标的影响
在光照试验生长过程中,光照强度均显著影响火力楠苗木的苗高、地径的生长(P<0.01)。由图1-图2可知,火力楠苗木苗高和地径在P1处理光照强度下生长表现最强,苗高和地径在8月份生长量均达到最大值。而随着光照强度的降低,火力楠苗高生长量显著降低,P3处理条件下,10月份火力楠苗高生长量降低了0.96cm,且表现出光照强度降低抑制了苗木的生长。随着生长季节的不同,地径的生长量差异显著(P<0.01)。其中火力楠苗木在P1处理条件下,8月份地径生长量达到了0.467cm,且试验过程中苗木生长均高于其他3个光照强度。但火力楠在P2、P3处理条件下,地径生长量不随着季节的变化而呈显著差异,地径生长量平均值分别为0.170cm、0.143cm。进而表明对苗木进行一层遮荫(42.3%NS透光率)有助于火力楠苗木地径的生长。
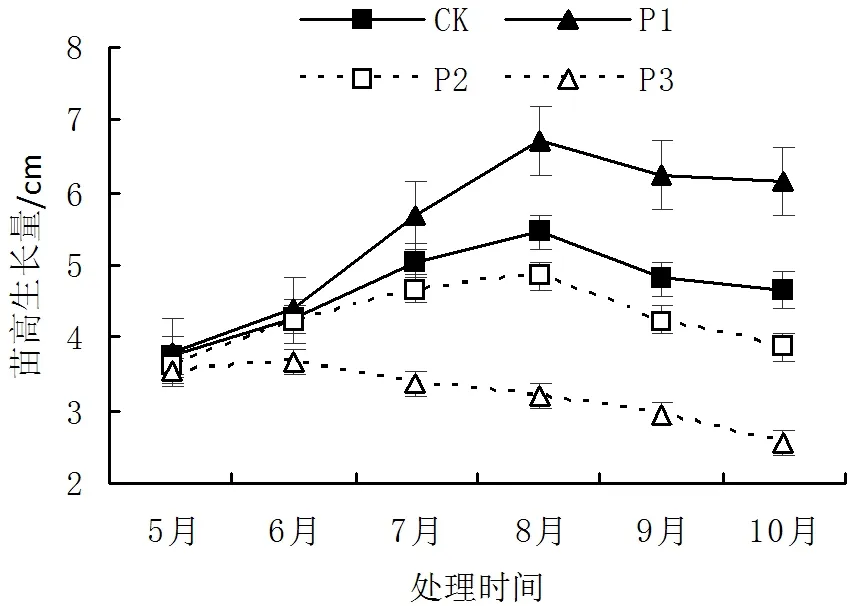
图1 不同光照强度对苗高生长影响Fig.1 Effect of height growth in different light intensities
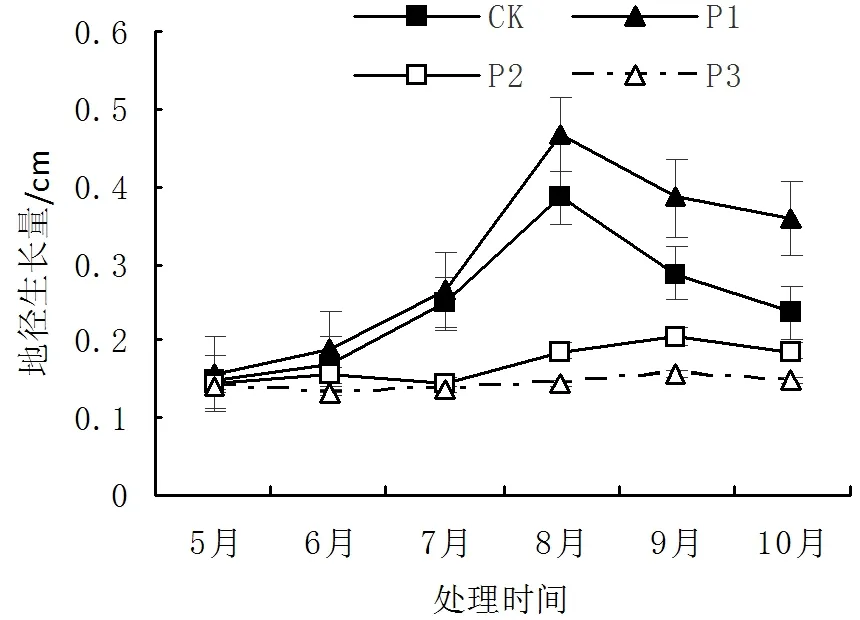
图2 不同光照对地径生长影响Fig.2 Effect of diameter growth in different light intensities
由图3可知,随着光照强度的变化,火力楠苗木叶面积变化呈显著的差异(P<0.05)。火力楠苗木在P1处理条件下,10月份苗木叶面积与CK、P2、P3处理相比,分别高出了25.49%、30.22%、69.91%。在P1条件下,火力楠叶面积8-10月份期间仍然处于不断增长的状态,而CK、P2、P3处理,在8-10月份期间,叶面积增长趋势平稳,分别稳定于27.48cm3、26.36cm3、20.34cm3。各处理火力楠苗木叶面积大小为P1>CK>P2>P3,对火力楠苗木进行适当地遮荫(42.3%NS透光率),有助于苗木叶面积增加,促进苗木光合作用。
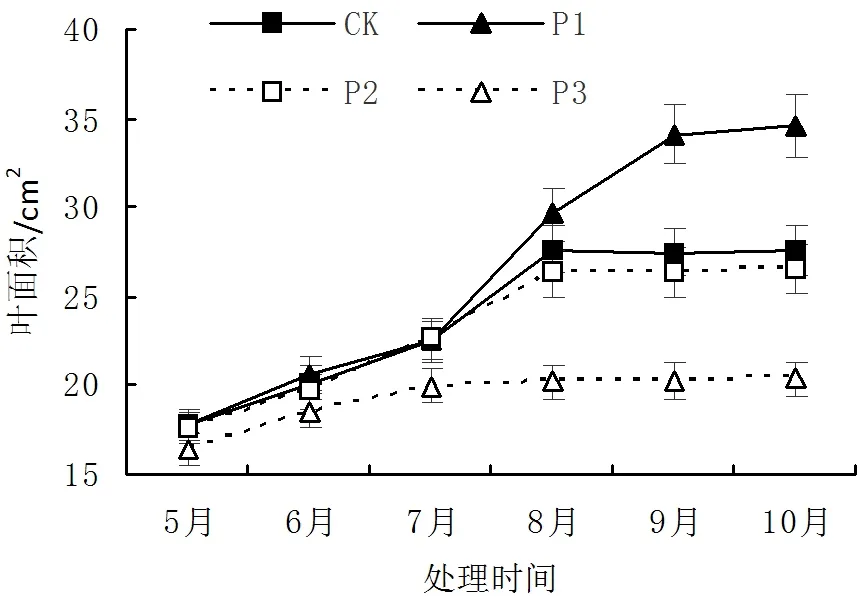
图3 不同光照对苗木叶面积影响Fig.3 Effect of leaf area in different light intensities
2.2 不同的光照强度对苗木光合指标的影响
由图4可知,火力楠苗木在光合有效辐射PAR达到776μmol/(m2·s)时,苗木的净光合速率Pn达到最大。其中,苗木在P1处理条件下,净光合速率Pn为各个处理最高,达到了9.46μmol/(m2·s)。火力楠苗木在P2条件下,光相应曲线与对照处理CK最接近。但随着光照强度的降低,苗木在P3条件下,光相应曲线最低,最大光合速率仅为3.23μmol/(m2·s)。表明了火力楠在弱光条件下,不利于光合作用的进行,而适当对苗木进行遮荫,有利于苗木光合作用,提高苗木的光合利用效率。
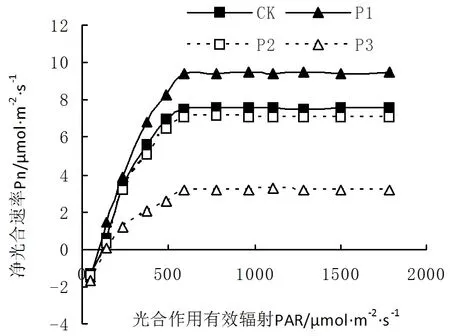
图4 不同光照条件下苗木光响应曲线变化Fig.4 Photoresponse curve of seedling under different illumination conditions
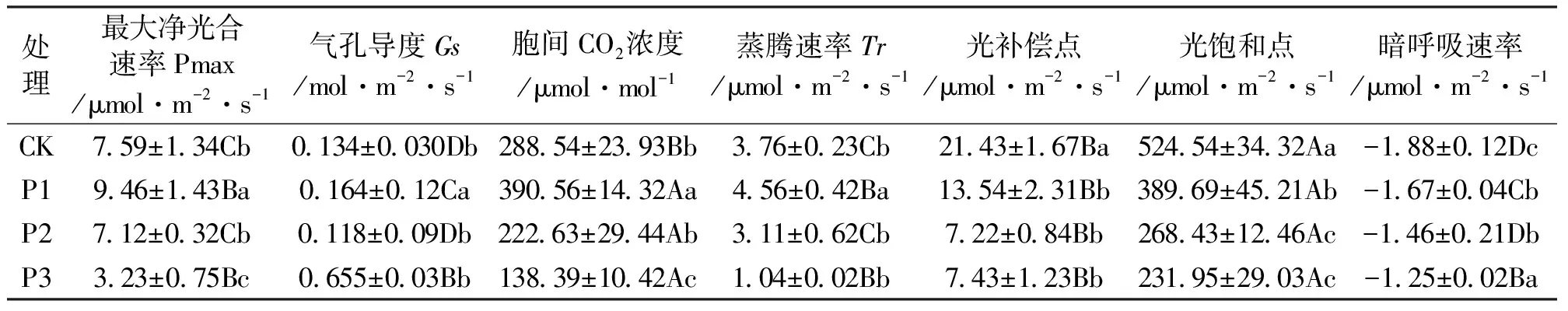
表1 不同光照苗木试验光合指标统计表Tab.1 Statistical table of photosynthetic indexes for different light seedlings
注:±数值表示每个处理3个重复的标准差值;大写字母表示不同光合指标间差异显著(P<0.05);小写字母表示不同光照处理间差异显著(P<0.05)。
在不同的光照强度条件下,火力楠各项光合指标差异显著(P<0.05)。由表1可知,火力楠在P1条件下最大净光合速率Pmax达到了最高,为9.46μmol/(m2·s),且对应的蒸腾速率也最高,达到了4.56μmol/(m2·s),且胞间CO2浓度含量最高,达到了390.56μmol/mol,但对应的暗呼吸速率仅为-1.67μmol/(m2·s)。在光照试验中,对照处理CK的Pmax与P2差异不显著,对照处理CK的光补偿点和光饱和点均为最高,分别为21.43μmol/(m2·s)和524.54μmol/(m2·s);P1光照强度下,气孔导度为4个处理最高,达到了0.164mol/(m2·s)。然而,随着光照强度逐渐减弱,火力楠苗木在P3处理条件下暗呼吸速率达到了-1.25μmol/(m2·s),火力楠苗木在P3条件下,暗呼吸速率最高,有氧呼吸最高。
2.3 不同光照对苗木生理生化指标的影响
在不同的光照强度对火力楠苗木的叶绿素含量、丙二醛含量(MDA)、超氧化物歧化酶活性(SOD)、过氧化氢酶活性(CAT)均形成了显著的影响(P<0.05),见图5。
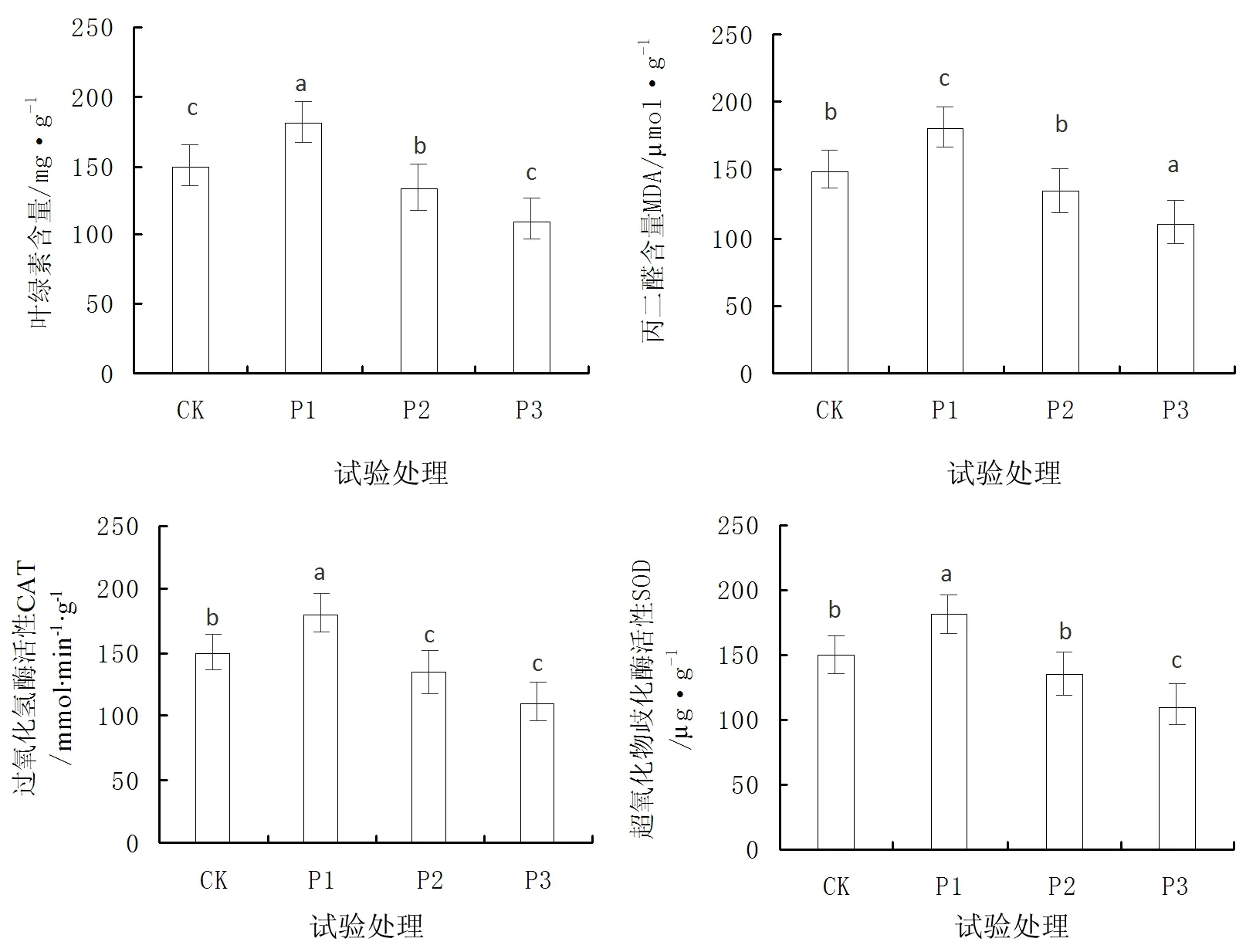
图5 不同光照强度对苗木生理生化指标的影响Fig.5 Effect of physiological and biochemical indexes in different light intensities
由图5可知,火力楠苗木在P1处理下,叶绿素含量相比对照CK处理高出了0.88mg/g,显著高于P2、P3处理。而苗木在P1处理下MDA含量仅为18.98μmol/g,为4个光照强度最低,P3处理条件下,MDA含量最高为29.32μmol/g。在火力楠苗木叶片酶的活性分析中,SOD和CAT均以P1光照条件下的苗木最高,分别达到了180.63ug/g、345.62μmol/(min·g),而P2和P3光照条件下苗木叶片SOD和CAT酶的活性均小于对照CK,且数值较为接近,表明了火力楠苗木在P2和P3光照条件下,酶的活性被抑制,降低了苗木化合物的合成与分解。
3 结论与讨论
对火力楠苗木各项生长指标分析发现,火力楠苗木的生长量随着试验的进行的变化而变化,苗木的苗高和地径在各个光照处理影响下,均在8月份达到最大值,其主要原因主要是因为该月份气温适宜,在充足的光照条件下,促进了火力楠苗木的生长[12],另外由于光照试验在进行了3个月后,苗木逐渐适应了光照环境并快速生长。其中,火力楠苗木在P1光照条件下,苗木苗高和地径显著高于同光照强度下速生秋枫(Bischofiajavanica)幼苗13.54%、6.43%,表现了火力楠苗木具有较高的生长能力[13]。在苗高生长量的比较中,P1光照处理显著高于对照CK,而P2条件下苗木苗高生长与对照CK较为接近,但苗木在P3条件下苗高生长出现了抑制情况,苗高生长量不断降低。在苗木地径的比较中,虽然苗木在P1条件下地径生长量表现最高,但仅略高于对照CK,且显著高于P2和P3处理,从而得出了对火力楠苗木进行遮荫(42.3%NS透光率),可有效地促进苗木苗高的生长,有利于苗木的枝干的生长发育。在对苗木叶面积的比较中,发现在光照试验持续进行过程中,火力楠苗木前期处于持续增长后期趋于平稳的状态,其中苗木在P1条件下,苗木叶面积比乐昌含笑(Micheliachapensis)、观光木(Tsoongiodendronodorum)平均叶面积高3.89cm2、2.16cm2[14],表现了火力楠叶片属于叶片较大树种。
不同光照强度试验对火力楠苗木光合指标造成了显著的影响,在分析苗木的光响应曲线中,发现火力楠苗木净光合速率在光合有效辐射PAR达到776μmol/(m2·s)时表现最大,火力楠苗木在P1条件下光相应曲线表现最强,而对照CK由于在全光照强度下,大量光照对苗木的光合作用造成了抑制,对照处理CK的光补偿点和光饱和点高出清香木(Pistaciaweinmannifolia)的8.53%、6.32%[15],表现了较强的光合能力;在全光照的条件下,为达到苗木光合作用最佳转换状态,降低了苗木的光合转换速率。而苗木在P1条件下,苗木可自动调整植物光合功能,提升了苗木的净光合转化速率,苗木的最大净光合值高出了合果木(Paramicheliabaillonii)1.24μmol/(m2·s)[16],且对应着高效的胞间CO2浓度、蒸腾速率;进而表明了苗木在P1条件下有利于光合物质的合成和物质间的转换。而随着光照强度的降低,火力楠苗木植物功能结合不断减弱,低的光照强度,抑制了苗木的光合作用过程,但苗木P3条件下,暗呼吸速率最高,说明了苗木在弱光条件下,可通过高效的有氧呼吸进行化合物质的合成。表明了火力楠具有一定的抗低光照的能力。
火力楠在P1条件下,叶片的叶绿素含量达到最高,高出了金露梅(Potentillafruticosa)叶片的5.35%[17],适当的对火力楠苗木进行遮荫有效的促进了苗木叶片叶绿素的合成,提升叶片光合作用效率。另外苗木在P1光照条件下酶的活性最高,SOD和CAT酶的活性相对木兰科常见树种鹅掌楸(Liriodendronchinense)生理生化活性,分别高出了10.42%和7.74%[18],适当的遮荫可有效调节酶活性,对植物体化合产物进行有效的分解和利用。但苗木在P1条件下MDA含量最低,表明苗木在该光照强度下叶片膜活性较低,未对苗木结构造成破坏,而苗木在P2和P3光照条件下对植物光合结构造成了影响,抑制了植物体酶的活性,通过提升MDA含量,增加苗木的抗逆性,进而表明火力楠可通过自身结构的变化应对逆境,具有抗遮荫性。因此,对火力楠苗木进行遮荫(42.3%NS透光率),可有效地促进火力楠生长和光合结构的形成,并获得高效的光合作用产物。
在对火力楠苗木进行不同光照试验研究中,对火力楠苗木进行遮荫(42.3%NS透光率),苗木生长和发育具有促进作用。通过植物生长指标、光合指标和生理生化指标进行验证,得出了火力楠苗木在层遮荫P1(42.3%NS透光率)条件下,生长发育最佳。在生产实践中,可通过对火力楠苗木进行适当的遮荫,提升苗木的苗高、地径和叶面积等指标,有利于获得优质、健康的火力楠苗木,提高火力楠苗木的质量。但本文仅对苗木的生理功能指标展开分析研究,未对植物细胞结构进行分析,因此在今后的研究可以对植物体解剖结构进行研究,有利于全面地分析光照胁迫对火力楠苗木生长发育的影响。
参考文献:
[1]种培芳,陈年来.光照强度对园艺植物光合作用影响的研究进展[J].甘肃农业大学学报,2008,43(5):104-109.
[2]刘璐,毛永成,王倩颖,等.木兰科植物光合作用的研究进展[J].贵州农业科学,2015,43(12):135-138.
[3]钟水红.火力楠的栽培技术[J].广西林业,2005(4):14-15.
[4]中国科学院植物志委员会.中国植物志[M].北京:科学出版社,2002.
[5]彭玉华,郝海坤,曹艳云,等.缓释肥和复合肥对火力楠容器苗的影响[J].南方农业学报,2015,46(1):47-52.
[6]陈宇超,张亨,梁逸葳,等.3种植物生长调节剂对火力楠苗木光合作用的影响[J].现代园艺,2015(15):13-15.
[7]欧斌,马小焕,赖锦良,等.赣南火力楠苗木质量分级指标研究[J].江西林业科技,2016,44(1):27-30.
[8]李松海,覃德文,韦中绵,等.广西木莲在干旱逆境下光合特性研究[J].北方园艺,2015,3(2):72-76.
[9]李付伸,覃德文,杜佩连,等.干旱胁迫对顶果木生理特性及根系形态特征的影响[J].湖北农业科学,2016,55(8):2019-2022.
[10]黄林敏,袁丛军,严令斌,等.不同遮荫处理对清香木苗木生长与光合速率的影响[J].浙江农业科学,2014,1(2):217-219.
[11]段如雁,韦小丽,孟宪帅.不同光照条件下花榈木幼苗的生理生化响应及生长效应[J].中南林业科技大学学报,2013,33(5):30-34.
[12]冯慧芳,薛立,任向荣,等.4种阔叶幼苗对PEG模拟干旱的生理响应[J].生态学报,2011,31(2):371-382.
[13]於艳萍,毛立彦,宾振钧,等.不同遮荫处理对秋枫幼苗生长的影响[J].北方园艺,2017,5(2):56-59.
[14]李超,赵广东,史作民,等.3种木兰科植物幼苗叶片功能性状及关联性分析[J].江西农业大学学报,2016,38(1):19-26.
[15]田勇,袁丛军,王加国,等.清香木对不同土壤水分胁迫的生长和光合响应[J].西部林业科学,2014,44(5):41-47.
[16]伍荣善,覃德文,秦武明.广西合果木生长及光合特性[J].西部林业科学,2015,45(3):68-72.
[17]刘行,张彦广.盐胁迫对金露梅叶片酶活性和叶绿素含量的影响[J].西部林业科学,2016,46(4):95-100.
[18]周艳威,李美平,鲁路,等.杂交鹅掌楸苗期NaCl胁迫响应研究[J].分子植物育种,2017,4(2):735-743.
猜你喜欢
江西农业(2022年8期)2022-05-13
绿色科技(2021年21期)2021-11-26
内蒙古林业调查设计(2021年3期)2021-07-06
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
政工学刊(2021年4期)2021-04-13
房地产导刊(2020年7期)2020-08-24
新疆农业科学(2020年1期)2020-02-14
现代园艺(2018年1期)2018-03-15
火力与指挥控制(2017年3期)2017-04-24
中国麻业科学(2015年5期)2015-12-28
