
“云新高原”核桃及其亲本的叶绿体DNA差异分析
2018-11-30 06:46肖良俊陈少瑜陈海云宁德鲁
西南农业学报 2018年10期
吴 涛,肖良俊,陈少瑜,陈海云,宁德鲁*
(1.云南省林业科学院,云南 昆明 650201;2.云南省木本油料工程技术研究中心,云南 昆明 650201;3.云南省森林植物培育与开发利用重点实验室/国家林业局云南珍稀濒特森林植物保护和繁育实验室,云南 昆明 650201)
【研究意义】云南省是深纹核桃(Juglanssigillata)的主要起源分布中心和栽培区域,截止2015年底已种植282万hm2,产量达到85万t,产值为266亿元,面积、产量和产值均居全国之首。漾濞泡核桃是云南传统主栽的深纹核桃品种,但存在结实晚(约10 a左右)、受益慢等问题。据此,云南省林业科学院通过种间杂交途径,用漾濞泡核桃与早实核桃(J.regia)进行杂交,培育出云新高原等早实杂交新品种核桃,并于2004年通过云南省林木品种审定委员会审定[1]。云新高原核桃的母本漾濞泡核桃树体高大、树势旺,小叶9~13片、多13片,结果枝多为中长果枝,种子刻纹深。云新高原核桃的父本为早实核桃优良单株‘云林A7’树体中等、树势较弱,小叶7~9片、多7片,结果枝以中短果枝为主,种子表面光滑。云新高原树势中等,树姿开张,小叶9~11片、多9片,结果枝多为中果枝,种子刻纹浅、表面较光滑。综合来看,云新高原的树高、干径、冠幅与父本相近,在早结实(1~3年开花结果)、顶芽形状、腋芽形状、有无芽距(主副芽是否分开)等习性上偏向于父本,复叶长、小叶数、小叶形状等性状介于双亲之间,呈趋中变异。在调查云南省核桃资源中,发现一些兼具深纹核桃和核桃特点的资源。鉴于云南曾于40年代进行过核桃的大面积引种栽培,推测这些资源可能是二者的天然杂交种。为此,希望能通过研究掌握人工杂交种云新高原的叶绿体基因组的遗传规律,来对这些“疑似杂交种”的身份进行确认。【前人研究进展】叶绿体DNA(cpDNA)属于细胞质基因,为绿色植物所特有,其遗传方式有别于核基因的遗传方式,属于细胞质遗传,在被子植物中存在双亲质体遗传和单亲质体遗传两种类型,通常以单亲母系遗传方式为主,而父系质体遗传在裸子植物中较为普遍[2-4]。叶绿体基因组很小,进化较慢,不存在基因重组和结构变异,更加便于遗传进化研究[5]。近年来,以特定片段测序为代表的第三代DNA分子标记技术,因具特异性强、通量高和高效快捷的优点而受到广泛的关注和应用[6]。研究者利用小片段叶绿体DNA测序的方法进行遗传多样性的研究,取得了较好的结果[7-10]。目前,DNA分子标记已经广泛应用于核桃遗传多样性研究[11],但关于cpDNA标记在核桃遗传多样性研究中的应用较少。【本研究切入点】基于核桃叶绿体基因组DNA序列信息,设计核桃cpDNA分子标记引物,并对云新核桃及其亲本的叶绿体DNA进行扩增产物的序列检测、分析和比较。【拟解决的关键问题】一是检测核桃叶绿体基因的遗传特性,二是开发出可应用于核桃起源及遗传多样性研究的分子标记。
1 材料与方法
1.1 试验材料
云新高原核桃(Juglanssigillata×J.reigacv. ‘Yunxingaoyuan’)、母本漾濞泡核桃(J.sigillatacv. ‘Yangpao’)、父本早实核桃(J.reigacv. ‘Yunlin A7’)的树龄分别为10年、20年、25年,均种植在云南省林业科学院核桃资源圃内。2017年5月采集健康新鲜叶片,即时移入实验室进行DNA提取。
1.2 研究方法
1.2.1 总DNA提取 总DNA用CTAB法提取。获得的总DNA经0.8 %的琼脂糖凝胶电泳进行质量检测,用超微量紫外分光光度计ND2000测定浓度和纯度,然后稀释为40 ng/μl,于-20 ℃保存备用。
1.2.2 引物设计 Hu等[12]获得了核桃属5个种的叶绿体基因组全长。本试验以其中的深纹核桃和核桃的cpDNA全长序列为基础(NCBI登录号分别为KX424843和KT963008),首先使用DNAMAN对二者的全长序列进行差异分析,然后使用Primer 5.0软件针对差异位点分别设计引物序列(表1),引物均由昆明硕擎生物科技有限公司合成。
1.2.3 PCR扩增与测序 PCR反应体系总体积20 μl,其中PCR Mixer 10 μl,引物(浓度10 μmol/L)各0.5 μl, DNA模板1 μl,ddH2O 8 μl。PCR扩增反应在扩增仪(Biometra TGradient,Germany)中进行,反应程序:94 ℃预变性4 min;94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸60 s,35个循环;72 ℃延伸7 min。PCR产物用1 %琼脂糖胶电泳检测,在凝胶成像仪上观察并切取目的条带,使用凝胶回收试剂盒(天根生化科技有限公司,北京)回收,回收产物送昆明硕擎生物科技有限公司采用ABI 3730XL自动测序仪(Applied Biosystems,USA)进行双向测序。
1.2.4 数据处理与分析 测序结果使用ContigExpress软件观察峰图并进行序列拼接及人工校对,手动删除序列两端50~60 bp不可靠的位点,拼接好的序列用 DNAMAN 9.0进行比对和差异位点的分析。
2 结果与分析
2.1 深纹核桃与核桃的叶绿体基因组序列差异分析
深纹核桃和核桃的叶绿体基因组全序列于2017年成功测序[12],全序列长度分别为160 350和160 367 bp。经序列对比分析,其序列变异信息见

表1 核桃叶绿体DNA扩增引物序列信息
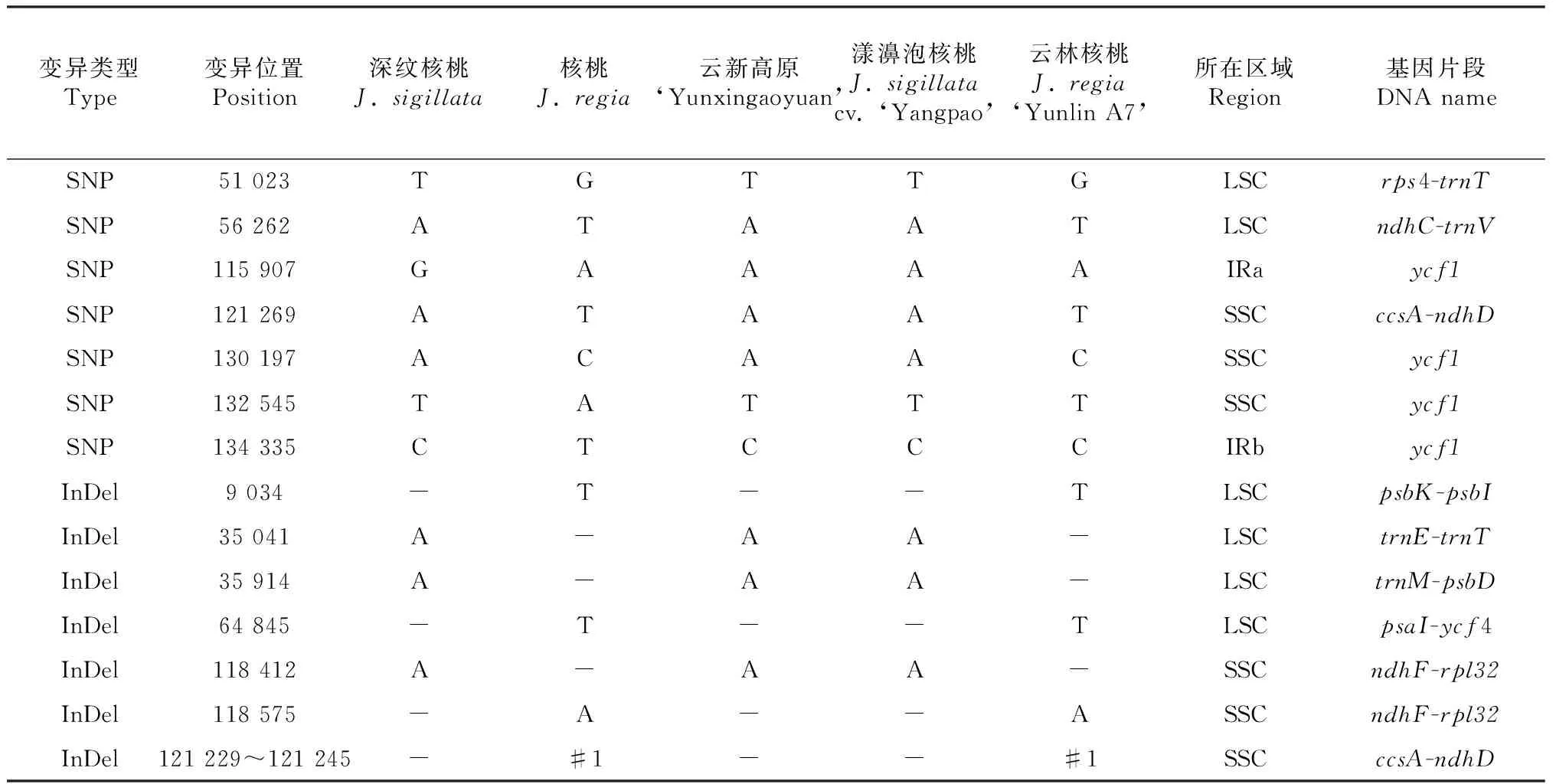
表2。深纹核桃和核桃的叶绿体基因组间仅有14个差异位点,总变异序列长度为30 bp,变异率为0.019 %。其中,单核苷酸多态性位点数(SNP数)有7个,2个为碱基转换,5个为碱基颠换;插入/缺失位点数(InDel数)有7个,6个为单碱基的InDel,1个为17碱基的InDel。分析结果表明,深纹核桃与核桃叶绿体基因组存在明显的差异,但二者不存在大范围的结构变异。
2.2 云新高原核桃与其亲本的叶绿体基因组扩增片段多态性分析
根据深纹核桃与核桃的叶绿体基因组间存在的14个变异位点,分别对云新高原及其父本(云林A7)、母本(漾濞泡核桃)的14个叶绿体基因片段进行测序及分析。结果表明,云新高原与母本在14个变异稳点上,碱基完全一致,但二者均与深纹核桃叶绿体模式基因组序列(KX424843)在115 907位点存在1个SNP(G/A碱基转换),变异之后与核桃叶绿体模式基因组序列(KT963008)保持一致;云新高原的父本与核桃叶绿体模式基因组序列(KT963008)存在2个SNP(132 545位点存在1个A/T碱基颠换,134 335位点存在1个T/C碱基转换),而云新高原与其父本在所检测的14个位点中,仅有11个位点存在差异。综合表2可以看出,14个检测位点中,云新高原与母本完全一致,而与父本存在11个位点的差异(占79 %),且差异位点仅有1个发生于基因编码区(占9 %),其余10个位点均位于非编码的基因间隔区(占91 %)。
另外,对9034(psbK-psbI基因间隔区)和118 575(ndhF-rpl32基因间隔区)变异位点检测的过程中发现,这2个扩增片段还分别含有8和14个变异位点,具体信息见图1。由图1可以看出,在这22个变异位点中,云新高原与其母本在其中10个位点上具有相同的SNP或InDel,余下的12个位点与其父本的相同。
2.3 云新高原核桃与其亲本的叶绿体基因单倍型差异分析
对云新高原核桃和其亲本的11个基因或基因间隔区片段进行单倍型分析,共发现有11种单倍型(表3),其中10个单倍型均表现为母系遗传,而剩余的ndhF-rpl32片段的单倍型较为复杂,存在7种亚单倍型,其中4种表现为母系遗传,3种表现为父系遗传,即云新高原核桃的ndhF-rpl32片段的单倍型表现为双亲遗传。
表2深纹核桃、核桃、种间杂交种云新高原及其亲本的叶绿体基因序列差异比较
Table 2 Comparisons of chloroplast DNA difference and polymorphism amongJ.sigillatacv. ‘Yangpao’,J.regia‘Yunlin A7’ and interspecific hybrid ‘Yunxingaoyuan’
变异类型Type变异位置Position深纹核桃J. sigillata核桃J. regia云新高原‘Yunxingaoyuan’漾濞泡核桃J. sigillata cv. ‘Yangpao’云林核桃J. regia ‘Yunlin A7’所在区域Region基因片段DNA nameSNP51 023TGTTGLSCrps4-trnTSNP56 262ATAATLSCndhC-trnVSNP115 907GAAAAIRaycf1SNP121 269ATAATSSCccsA-ndhDSNP130 197ACAACSSCycf1SNP132 545TATTTSSCycf1SNP134 335CTCCCIRbycf1InDel9 034-T--TLSCpsbK-psbIInDel35 041A-AA-LSCtrnE-trnTInDel35 914A-AA-LSCtrnM-psbDInDel64 845-T--TLSCpsaI-ycf4InDel118 412A-AA-SSCndhF-rpl32InDel118 575-A--ASSCndhF-rpl32InDel121 229~121 245-#1--#1SSCccsA-ndhD
注:“-”表示碱基缺失(InDel);“#1”表示序列TATTTTTAACTTAAGTT;“LSC”表示大单拷贝区域;“SSC”表示小单拷贝区域;“IR”表示反向重复序列。
Note:‘-’: Base deletion (indels); ‘#1’: TATTTTTAACTTAAGTT; ‘LSC’: Large single-copy; ‘SSC’: Small single-copy; ‘IR’: Inverted repeat.

*:碱基缺失*: single nucleotide deletion图1 云新高原核桃及其亲本与深纹核桃叶绿体模式基因组序列的2个片段的多态性比较Fig.1 Comparisons of two chloroplast DNA fragments polymorphism among ‘Yunxingaoyuan’, its parents and Juglans sigillata model sequence

编号No.基因片段DNA name变异位置Position单倍型 Haplotype漾濞泡核桃J. sigillata cv. ‘Yangpao’云林A7核桃J. regia ‘Yunlin A7’云新高原‘Yunxingaoyuan’1psbK-psbI9034A1A2A12psbI-trnS9256,9258,9279,9285,9312B1B2B13trnS9325,9329,9340C1C2C14trnE-trnT35 041D1D2D15trnM-psbD35 914E1E2E16rps4-trnT51 023F1F2F17ndhC-trnV56 262G1G2G18psaI-ycf464 845H1H2H19ndhF-rpl32118 412I1I2I1118 471I3I4I3118 473, 118 475, 118 481, 118 494, 118 503, 118 521, 118 533, 118 537I5I6I6118 575I7I8I7118 576, 118 594, 118 598I9I10I10118 600I11I12I11118 604I13I14I1410ccsA-ndhD121 229~121 245, 121 269J1J2J111ycf1130 197K1K2K1
注:粗体显示云新高原叶绿体DNA的单倍型与其父本相同。
Note: Bold characters showed that the chloroplast haplotype of ‘Yunxingaoyuan’ was identical to that of its farther parent.
3 讨 论
本项研究基于网络数据库中已公布的序列,从叶绿体全基因组层面分析,发现深纹核桃与核桃的叶绿体基因组存在14个变异位点、30个碱基的差异。总体上,93 %的差异类型(13/14)为单碱基多态性变异,其中7个为SNP,6个为单碱基InDel;7 %的差异类型(1/14)为17个碱基的短片段InDel,不存在大范围的结构变异,这与水稻[13]、大麻[14]和其它植物[15]关于叶绿体基因组结构的研究相同。
针对云新高原核桃及其父母本的这14个变异位点进行多态性检测,发现仅11个位点在三者间存在多态性,另外3个位点的碱基之间则完全相同,不具有多态性。这11个多态性位点中,云新高原与其母本完全一致,而与其父本完全不一致。并发现有1个位于基因编码区,其余10个都位于非编码的基因间隔区。在psbI-trnS和ndhF-rpl32两个片段序列中,另发现存在22个变异位点,其中19个位点位于基因间隔区,云新高原与其母本在这22个位点中有10个保持一致,而与其父本在余下的12个位点保持一致。综合而言,云新高原与其亲本三者间共检测发现33个变异位点,云新高原与其母本在其中21个位点上保持相同碱基,占全部变异位点数的64 %;云新高原与其父本在余下12个位点上保持相同碱基,占全部变异位点数的36 %。这33个变异位点分属11个基因或基因间隔区片段,可以分为11种单倍型,云新高原的这11个单倍型中有10个与母本相同,1个为双亲的嵌合体。据此推断,云新高原的叶绿体DNA为以母系遗传为主的双亲遗传。这与草本植物绿绒蒿(Meconopsis)相类似,其种间杂交种与其亲本在trnL-trnF片段中存在25个变异位点,其中84 %的位点与母本一致,4 %的位点与父本一致,其余12 %的位点与亲本均不相同[16]。另外,ndhF-rpl32DNA片段具有多个变异位点,是多态性高的叶绿体基因片段,有望广泛应用于核桃群体遗传分析、谱系地理和种内品种鉴定等研究。
大多数被子植物的叶绿体DNA通过雌配子以母系遗传的方式传递[2-3],父系遗传仅见于紫花苜蓿(Medicagosativa)[17]、甘草(Glycyrrhizaspp.)[18]和牵牛花(Pharbitisnil)[3]等少数种类中;裸子植物中父系遗传较为普遍[3-4];天竺葵属(Pelargonium)和月见草属(Oenothera)植物是双亲遗传方式的代表[3]。此外,随着研究手段和研究材料的更新和丰富,之前认为猕猴桃(Actinidia)是严格的父系遗传,但后来发现还具有母系及双亲遗传的特性,与杂交亲本的基因型有一定关系[19]。总之,植物叶绿体DNA的遗传方式具有多样性和复杂性。而胡桃科植物的叶绿体DNA遗传方式的研究尚无相关报道,现有的相关叶绿体DNA研究都是针对系统进化和植物分类开展的。本项研究发现,核桃叶绿体DNA的遗传方式为以母系遗传为主的双亲遗传,这为今后核桃种间杂交和遗传变异研究提供了新的资料。
4 结 论
深纹核桃与核桃的叶绿体基因组存在14个变异位点、30个碱基的差异,其中7个为核苷酸多态性变异(SNP),7个为碱基插入/缺失变异(InDel)。对云新高原及其亲本的这14个位点及其附近片段进行检测,发现三者存在33个变异位点,云新高原与其母本在其中21个位点上保持相同碱基,占全部变异位点数的64 %;云新高原与其父本在余下12个位点上保持相同碱基,占全部变异位点数的36 %。这33个变异位点分属11个基因或基因间隔区片段,可以分为11种单倍型,云新高原的这11个单倍型中有10个与母本相同,1个为双亲的嵌合体。根据质体遗传的特性,推断核桃叶绿体DNA的遗传方式为以母系遗传为主的双亲遗传。另外,核桃ndhF-rpl32DNA片段存在多个变异位点,多态性高,该叶绿体基因片段可广泛应用于核桃系统发育、群体遗传和种内品种鉴定等研究。
猜你喜欢
中国甜菜糖业(2022年4期)2023-01-21
教学考试(高考生物)(2020年6期)2020-11-23
家教世界·创新阅读(2020年4期)2020-06-03
当代工人·精品C(2020年1期)2020-05-20
家教世界·创新阅读(2020年1期)2020-05-11
家教世界·创新阅读(2020年2期)2020-04-07
食品与生物技术学报(2020年8期)2020-01-06
课程教育研究·学法教法研究(2019年18期)2019-10-08
少儿科技(2019年9期)2019-09-10
学苑创造·B版(2019年5期)2019-06-14
