
超级增强子在肿瘤研究中的进展
2019-01-30 06:39吴志强米泽云
遗传 2019年1期
吴志强,米泽云
超级增强子在肿瘤研究中的进展
吴志强1,2,米泽云3
1. 天津医科大学肿瘤医院,放射治疗科,天津 300060 2. 国家肿瘤临床医学研究中心,天津市“肿瘤防治”重点实验室,天津市恶性肿瘤临床医学研究中心,天津 300060 3. 天津医科大学基础医学院,生物化学与分子生物学系,天津 300070
超级增强子是由多个相邻近的普通增强子组成的、驱动调控细胞身份基因表达的一个大簇,该区域富集高密度的转录因子、辅因子及增强子相关表观修饰。超级增强子所驱动的异常转录基因对维持肿瘤细胞特性至关重要。肿瘤细胞通过组装自身超级增强子,显著促进多种癌基因表达,从而增强肿瘤细胞的增殖、侵袭和转移的能力;抑制超级增强子的活性,则显著抑制肿瘤细胞的生长和存活。本文对目前报道的肿瘤细胞中超级增强子的结构特征和功能调控,以及靶向超级增强子药物研发现状进行了总结,旨在为研发新的针对超级增强子为靶点的抗肿瘤药物提供理论基础和借鉴。
增强子;超级增强子;转录;癌症
20世纪80年代,研究发现SV40病毒的一段DNA序列对于家兔()的β-珠蛋白(β-globin,一种能够通过铁卟啉环可逆性结合氧的呼吸性蛋白质)的转录具有增强作用,因此将这一段DNA称为增强子(enhancer)[1]。随后的研究发现在哺乳动物细胞内也存在类似特性的DNA序列,可以远距离、无方向性的增强基因转录[2~4]。近30年研究证明增强子具有以下特征(图1)[2~5]:(1) 增强子DNA序列处于染色体疏松的区域,与核小体中组蛋白的修饰,转录因子的结合有关;(2) 增强子活性与其DNA序列结合的组蛋白H3的第4位赖氨酸单甲基化(H3K4me1)和第27位赖氨酸乙酰化(H3K27ac)修饰程度成正相关[6];(3) 增强子发挥功能需要增强子区域和启动子的区域的直接相互作用,形成三维环状结构(3D-loop)。增强子和启动子的相互作用由多种蛋白介导,如Mediator复合体、Cohesin等[6,7]。
随着DNA测序技术的发展,人类对基因有了进一步的认识,对于增强子的研究也越来越深入。2013年,美国Young R.A.教授(Whitehead Institute for Biomedical Research)基于当时增强子的研究首次提出超级增强子(super enhancers,SEs)这一概念。他们发现胚胎干细胞(embryonic stem cells,ESC)的主要转录因子结合在一些特殊的增强子上,这些特殊的增强子对于维持胚胎干细胞的干性至关重要,并将这些特殊的增强子定义为超级增强子[8]。超级增强子简单来说就是由多个增强子组成的一个大簇,富集高密度的转录因子、辅因子和增强子表观修饰。它和普通增强子在序列大小、转录因子的结合密度、激活转录的能力以及对转录因子抑制剂的敏感性均不同[8]。随后的研究不仅发现超级增强子存在于多种细胞类型中,也进一步明确了超级增强子区别于普通增强子的功能特性(图2)[6]:(1) 超级增强子具有高密度的H3K27ac和H3K4me1修饰,以及Mediator复合体和Bromodomain containing 4蛋白(BRD4,与组蛋白乙酰化修饰位点结合)的结合;(2) 超级增强子结合的转录因子以及与转录活性相关的染色体的标记比普通增强子高很多;(3) 超级增强子调控的基因比普通增强子调控的基因表达水平高很多;(4) 组成超级增强子的单个增强子也可以像普通增强子一样激活基因转录;(5) 超级增强子可以结合组织中特异的转录因子;(6) 与普通增强子相比,超级增强子活性对于转录因子的阻断更敏感[9,10]。这些现象支持一个假说:超级增强子发挥功能需要结合到超级增强子上的转录因子的合作协同,具有大量转录因子结合的增强子对于基因转录的调控会对转录因子浓度的改变更敏感[11](图2)。有趣的是,富集在超级增强子上的主要的转录因子也受超级增强子的调控转录,这就意味着超级增强子调控基因转录存在正反馈协同作用,也就形成了细胞中的核心转录调控环路(coretranscriptionregulatorycircuitry, CRC)[12,13]。正是由于超级增强子调控基因表达的特性和其敏感性,因而才能够协调细胞在生长、发育、分化和疾病等各种状态的过渡[9,14~16]。本文主要从超级增强子与肿瘤细胞的关系、在肿瘤细胞中的调控以及该靶点药物在肿瘤治疗中的现状这3个方面阐述超级增强子在肿瘤细胞中的作用。

图1 增强子的结构特征和功能
A:增强子的结构特征和表观修饰。增强子区域染色体比较疏松,部分DNA暴露并且富集转录因子;增强子区域H3组蛋白具有H3K4me1和H3K27ac修饰。B:转录因子结合到增强子区域之后进一步招募Mediator复合体,其主要介导增强子与PolⅡ相互作用,这样增强子与启动子之间形成三维环状结构,从而增强基因的转录水平。
1 超级增强子与肿瘤的关系
2013年,Young R.A.教授发现在多发性骨髓瘤细胞中超级增强子的区域募集了高浓度的Mediator复合体和BRD4[17],这意味着在多发性骨髓瘤细胞中超级增强子处于活化状态。功能分析实验表明,超级增强子调控的基因(、、、)对于多发性骨髓瘤的发生和发展起到关键的促进作用[17]。后续研究发现,超级增强子在多种肿瘤中均有报道,如弥漫性大B细胞淋巴瘤[18]、T细胞急性淋巴细胞白血病[19,20]、默克尔细胞癌[21]、急性髓性白血病[22]、小细胞肺癌[10]、卵巢癌[23]、上皮癌[24]、鳞状细胞癌[25]、黑色素瘤[15]、乳腺癌[26]、食管鳞状细胞癌[27]和结肠癌[28]等。在肿瘤细胞中超级增强子调控的关键癌基因在正常细胞中是不表达的,这就提示超级增强子通过调控这些基因而对肿瘤生成和肿瘤特性维持起到关键作用[9,29,30]。由于细胞在癌变过程中大多数的超级增强子是重新形成具有功能性的元件,因此超级增强子的活化可以作为细胞癌变的一种标志[9,29,30]。综上所述目前研究均表明超级增强子的激活可以促使正常细胞向肿瘤细胞的恶性转化。
超级增强子不但对蛋白编码基因具有转录激活作用,对非编码基因,如microRNA (miRNA,一种长度约22nt的小RNA)的转录及成熟也具有调控功能。美国麻省理工学院生物系Phillip A. S.教授研究组利用CRISPR/Cas9基因组编辑方法发现超级增强子不仅促进miRNA的转录,也可以通过招募Drosha/DGCR8蛋白复合体促进前体miRNA (pri- miRNA)的成熟,以此来调控细胞种类特异性miRNA的生成[31]。对18种肿瘤细胞分析发现,在有些肿瘤细胞中超级增强子活性上调,而有些肿瘤细胞中超级增强子活性下降。进一步分析表明在细胞癌变过程中激活的超级增强子往往与促癌miRNA相关,而失活的超级增强子主要调控抑癌miRNA的生成[31]。以上研究提示,调控miRNA的超级增强子活性与肿瘤发生发展密切相关。因此,超级增强子联合多个miRNA (SE-miRNA)将有潜力成为细胞癌变的生物标志物[31],对于肿瘤的早期诊断以及治疗具有重要的临床意义。除此之外,超级增强子还可以调控长链非编码RNA (long non-coding RNA, lncRNA)的转录[32]。在鳞状细胞癌组织中发现受超级增强子调控的lncRNA LINC01503明显上调。进一步研究发现,LINC01503的表达水平与鳞状细胞癌病人预后呈负相关:LINC01503高表达的病人生存率低。研究表明激活的增强子或超级增强子区域也可以被转录产生RNA,称为enhancer RNA (eRNA),eRNA可以协同超级增强子激活转录[33,34]。超级增强子发挥功能不仅依赖于和启动子之间的3D-loop的形成,也依赖于超级增强子转录的eRNA的生成。因此,在临床上可以结合lncRNA以及eRNA的水平对病人进行精准治疗。
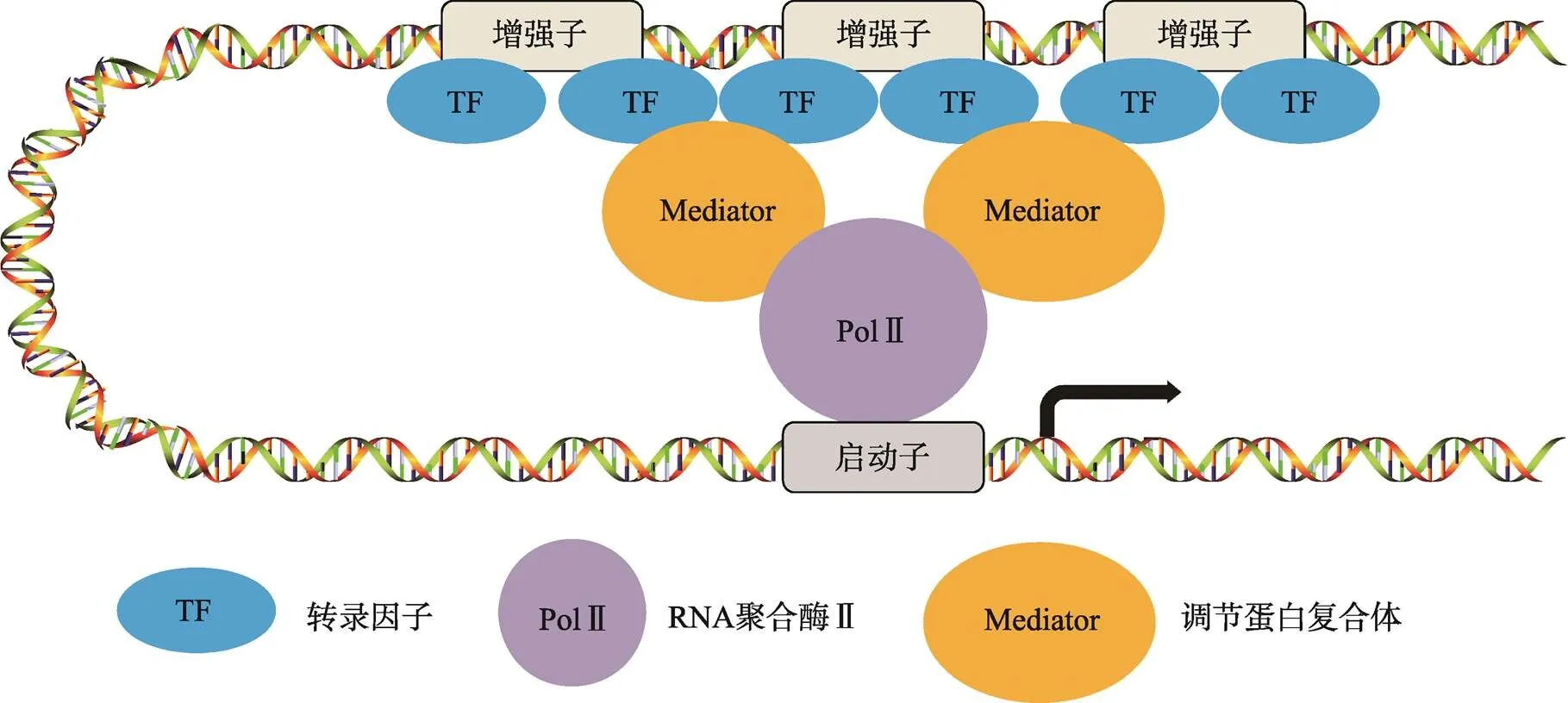
图2 超级增强子的结构
超级增强子是由多个增强子组成的一个大簇。组成超级增强子的单个增强子基因组的距离比较近,它们均可以独立的结合转录因子、Mediator复合体等辅因子,共同调控同一启动子的转录活性。
肿瘤的异质性很大一方面是由于一个肿瘤内的细胞通常可能来源于多个不同的细胞克隆,而这些不同克隆来源的肿瘤细胞其超级增强子的激活也存在差异,这就为区分肿瘤亚型或肿瘤细胞亚群提供了一种新的鉴定方法。例如,通过以往的方法对成神经管细胞瘤的生物化学和遗传学分析把其分成4个亚型。但是通过对这4个亚型的增强子图谱分析发现了一种新的亚型,这种新型的成神经管细胞瘤细胞中都具有与肿瘤异质性相关的超级增强子群[35]。更为重要的是通过分析在这类肿瘤细胞的超级增强子调控的转录因子可以明确细胞特异性的核心转录调控环路(CRC)。通过对于CRC分析确定了LIM homeobox transcription factor 1 alpha (LMX1A,一种转录因子)在第4类亚型的成神经管细胞瘤是一个主要转录因子(master transcription factor)[36]。同样,在其他基因异质性癌中也发现类似情况,如三阴性乳腺癌依靠超级增强子调控的特异性的基因群来维持细胞生长和增殖[26]。可见通过对于不同肿瘤细胞的增强子的图谱分析可以独立预测肿瘤亚型,发现之前治疗的不足以及新的潜在治疗靶点,为肿瘤治疗提供新思路、新方向[6]。
综上所述,在多种肿瘤细胞中均发现超级增强子处于异常激活状态,其对于靶基因的调控呈多样化:促进mRNA的生成、促进miRNA的转录以及成熟、促进lncRNA的转录生成以及超级增强子自身转录生成的eRNA对于其活性也起到协同作用。除此之外,通过绘制肿瘤细胞的增强子图谱可以预测肿瘤亚型,为基因异质性肿瘤提供统一的治疗 平台。
2 肿瘤细胞中超级增强子的调控
在肿瘤细胞中超级增强子的调控是如何实现的呢?早期对于小鼠胚胎干细胞发育的研究提出一个模型:组成超级增强子的每一个增强子都具有活性,而超级增强子的功能类似于一个平台,这个平台汇集了与发育相关的信号通路传递过来的信号,这些信号协同调控超级增强子活性启动基因转录(图3)[11]。同样,与癌基因相关的超级增强子也富集了肿瘤细胞依赖的信号通路的转录因子。在Wnt信号通路异常引起的结肠癌细胞中,相关的超级增强子区域富集了很多由Wnt信号通路终端的转录因子4 (transcription factor 4, TCF4),通过激活或者抑制Wnt信号通路,可以控制超级增强子调控的基因转录[11,37]。在雌激素受体(estrogen receptor, ER)阳性的乳腺癌细胞中,相关的超级增强子区域聚集了大量的ERα;而在三阴性乳腺癌细胞中缺少类固醇激素的表达,与其相关的超级增强子区域富集了完全不同的转录因子[26,37]。
在肿瘤细胞中,信号通路从多方面对超级增强子的活性进行调控。2015年美国西北大学Licht J.D. 教授团队研究发现Ras-Erk活性与超级增强子的活性密切相关:抑制Ras蛋白的活性会导致超级增强子相关的特征(如H3K27ac)消失、活性下降、降低相关基因转录;激活Ras可以增强调控癌基因的超级增强子活性[38]。另一方面,促癌信号通路可以通过操纵转录机器调节超级增强子的活性。转录暂停是激活的RNA聚合酶Ⅱ(RNA polymerase Ⅱ,Pol Ⅱ)在启动子附近停止转录的一种状态[39]。在正常的肝细胞中,Hippo信号通路可以通过限制暂停的Pol Ⅱ释放,因而抑制了增强子或超级增强子调控的基因转录[40]。然而,在肝癌细胞中Hippo信号通路的缺失导致YAP(Yes associated protein)入核,YAP蛋白结合到超级增强子上,招募Mediator复合体和细胞周期素依赖性激酶9(Cyclin-dependent kinase 9,CDK9),使暂停的Pol Ⅱ进入到延伸状态,促进癌基因转录[41]。因此,在肝癌中YAP通过激活超级增强子促进癌基因的转录。
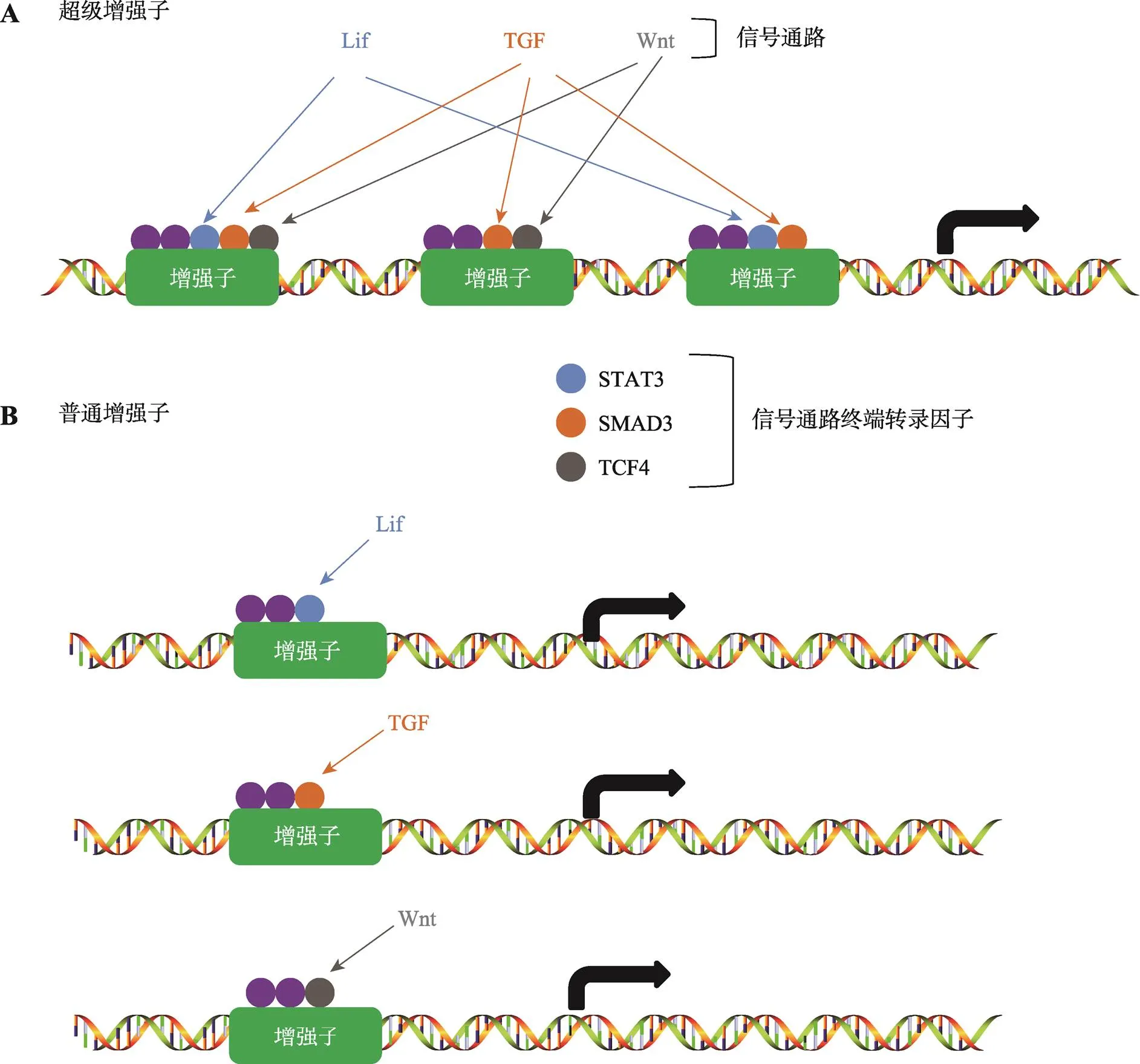
图3 信号通路对于超级增强子与普通增强子的调控
A:组成超级增强子的每一个增强子都具有活性,每个增强子富集了各种信号通路终端转录因子,这些转录因子协同调控基因转录;B:普通增强子只富集一种信号通路的终端转录因子。
以上研究表明超级增强子可以作为连接癌基因信号通路和维持肿瘤细胞特性的基因转录表达的渠道。然而进一步研究发现信号通路对于超级增强子的调控与转录因子在超级增强子区动态结合有关。例如,在NOTCH1异常导致的T细胞白血病(T-ALL)细胞中,NOTCH1在基因组上具有普遍的结合,但是只有不到10%的基因对于NOTCH1信号通路的改变有应答,而这些应答基因的NOTCH1结合在对应的超级增强子上。如果这些位点丢失NOTCH1的结合就会导致超级增强子的特征消失[42]。
3 针对超级增强子在肿瘤治疗中的作用
在肿瘤细胞中癌基因被转录激活,进而介导细胞的增殖和永生化[29,30]。因此,抑制癌基因的转录是一个潜在的治疗靶点。但是针对这一靶点面临着巨大的挑战:转录是细胞最基本的功能,对癌基因的转录抑制可能会引起细胞基因转录的广谱抑制[43]。因此,临床上用到的转录抑制剂应该特异性的抑制癌基因,对于正常细胞的转录影响不大[6,44~46]。
转录起始、暂停、延伸等过程的有序转变都是通过转录因子调控。目前研究发现超级增强子调控的转录依赖于BRD4、Mediator复合体、包含细胞周期素依赖性激酶7 (cyclin-dependent kinase 7, CDK7)的TF ⅡH复合体和包含CDK9的转录延伸复合体(P-TEFb)[39]。CDK7对Pol Ⅱ C末端区域(C terminal domain,CTD)第5位丝氨酸磷酸化起始转录[47];CDK9主要对Pol Ⅱ CTD 第2位丝氨酸磷酸化促进转录暂停的Pol Ⅱ进入转录延伸阶段,也称作Pol Ⅱ的释放[39]。另外,BRD4通过招募Mediator复合体促进超级增强子的装配进而促进暂停状态的Pol II的释放[48]。CDK12/13可以加速Pol Ⅱ的转录延伸[49,50]。因此,目前普遍认为超级增强子调控转录的关键调节点Mediator复合体、BRD4和关键的CDK有利于开发成治疗肿瘤的新靶点[6]。基于上述的转录抑制的关键节点,目前对于超级增强子这一靶点的药物主要有以下几类:(1) 针对BRD家族蛋白的抑制剂或者降解剂;(2) CDK7抑制剂;(3) 其他类型抑制剂(表1)。

表1 在肿瘤治疗中以超级增强子为靶点的小分子抑制剂研究现状
“-”代表未进入临床试验。
JQ1通过与BRD4的bromodomain结构域结 合而抑制BRD4与发生乙酰化修饰的蛋白相互作用[59,60],也就限制了BRD4和超级增强子的H3K27ac位点结合,抑制超级增强子和启动子的相互作用,进而影响癌基因的转录[17]。由于超级增强子所调控的转录对于转录因子的浓度变化特别敏感,JQ1处理可以优先阻止BRD4和超级增强子上的乙酰化修饰位点结合,进而特异性的抑制超级增强子介导的转录激活[9,17]。除此之外,还发现BRD抑制剂iBET762、OTX015、CPI0610和iBET151等,前3者已经进入临床实验阶段[22,51,61]。dBET系列化合物是基于JQ1的化学结构研发的特异性更高的BRD4抑制剂,其可以特异性的介导BRD家族蛋白的降解[62],从而阻止BRD家族蛋白识别超级增强子的乙酰化位点,影响超级增强子活性,抑制转录[63]。研究表明,BETd- 246可以靶向特异的降解BRD家族蛋白,相对于iBET- 211在三阴性乳腺癌中也表现出更好的治疗效果[52]。
THZ1是CDK7特异的抑制剂,依赖于超级增强子介导的肿瘤细胞对于THZ1高度敏感[64]。THZ1可以和CDK7的第312位半胱氨酸共价结合,抑制CDK7的激酶活性,从而抑制CDK7对于Pol Ⅱ CTD的第五位丝氨酸磷酸化,抑制转录起始,进一步阻止Pol Ⅱ在启动子近端暂停。超级增强子主要调控暂停的Pol Ⅱ释放,THZ1处理之后在启动子近端的Pol Ⅱ减少,也减少在增强子处的Pol Ⅱ结合,最终抑制转录[55,64]。THZ1处理之后超级增强子活性下降,导致多种癌基因转录抑制,从而抑制多种肿瘤细胞的生长和增殖[64]。SY-1365是Syros公司研发的CDK7的特异性抑制剂,可以选择性抑制多种实体瘤(乳腺癌、卵巢癌和小细胞肺癌等)和血癌(急性髓细胞样白血病和急性淋巴细胞白血病),目前对于晚期实体瘤的实验处于一期临床阶段。该公司通过对肿瘤细胞的基因检测分析,发现急性髓细胞白血病病人和骨髓增生异常综合征病人具有受超级增强子调控的RARA和IRF8基因的高表达,并且发现SY- 1425可以作用于维甲酸受体α(RARα),在临床上对上述两类病人具有较好的治疗效果。
CDK12是调节转录延伸的一个激酶,在T细胞白血病中THZ531可以特异性抑制CDK12/13,有效抑制超级增强子介导的基因表达[57]。在急性髓性白血病中抑制Mediator激酶(CDK8/19)活性可以上调肿瘤抑制因子相关的超级增强子活性,激活肿瘤抑制基因的表达,最终达到抗白血病的活性[22]。类似的,CDK4/6抑制剂LEE011选择性抑制CDK4,下调cyclin D1相关的超级增强子活性,有效的促进尤因肉瘤细胞的凋亡[58]。
4 结语与展望
目前关于超级增强子的研究,发现超级增强子在多种肿瘤细胞中均有激活,而激活的这些超级增强子往往促进癌基因的产生,维持癌细胞特性。通过抑制CDK、BRD4和Mediator复合体均可以干扰超级增强子的活性。超级增强子的先驱者Young R.A.与JQ1/iBET研发者Bradner J.E.这两位科学家曾预言超级增强子具有广阔的研发前景和价值,必将成为下一个药物研发的黄金靶点,因为针对这一靶点有望开发一种精确影响基因调控元件的药物,为此这两位科学家联手成立Syros公司专门研发针对超级增强子这一靶点的抗癌药。但是到目前为止对于超级增强子各个组分的研究还有欠缺:在癌细胞中组成超级增强子的每个增强子的活性是否和正常细胞中对应的增强子活性一致?信号通路活性的改变是怎样影响单个增强子组装成超级增强子的?从治疗角度考虑,也需要探索清楚超级增强子各个组分之间是怎么发挥作用的,药物是怎样抑制超级增强子各组分的活性。
[1] Banerji J, Rusconi S, Schaffner W. Expression of a beta- globin gene is enhanced by remote SV40 DNA sequences.,1981, 27(2 Pt 1): 299–308.
[2] Blackwood EM, Kadonaga JT. Going the distance: a current view of enhancer action.,1998, 281(5373): 60–63.
[3] Bulger M, Groudine M. Functional and mechanistic diversity of distal transcription enhancers.,2011, 144(3): 327–339.
[4] Sun CB, Zhang X. Advance in the research on super- enhancer.,2016, 38(12): 1056–1068.孙长斌, 张曦. 超级增强子研究进展. 遗传, 2016, 38(12): 1056–1068.
[5] Levine M. Transcriptional enhancers in animal development and evolution.,2010, 20(17): R754–763.
[6] Sengupta S, George RE. Super-enhancer-driven transcriptional dependencies in cancer.,2017, 3(4): 269– 281.
[7] Ing-Simmons E, Seitan VC, Faure AJ, Flicek P, Carroll T, Dekker J, Fisher AG, Lenhard B, Merkenschlager M. Spatial enhancer clustering and regulation of enhancer- proximal genes by cohesin.,2015, 25(4): 504–513.
[8] Whyte WA, Orlando DA, Hnisz D, Abraham BJ, Lin CY, Kagey MH, Rahl PB, Lee TI, Young RA. Master transcription factors and mediator establish super-enhancers at key cell identity genes.,2013, 153(2): 307–319.
[9] Hnisz D, Abraham BJ, Lee TI, Lau A, Saint-Andre V, Sigova AA, Hoke HA, Young RA. Super-enhancers in the control of cell identity and disease.,2013, 155(4): 934–947.
[10] Christensen CL, Kwiatkowski N, Abraham BJ, Carretero J, Al-Shahrour F, Zhang T, Chipumuro E, Herter-Sprie GS, Akbay EA, Altabef A, Zhang J, Shimamura T, Capelletti M, Reibel JB, Cavanaugh JD, Gao P, Liu Y, Michaelsen SR, Poulsen HS, Aref AR, Barbie DA, Bradner JE, George RE, Gray NS, Young RA, Wong KK. Targeting transcriptional addictions in small cell lung cancer with a covalent CDK7 inhibitor.,2014, 26(6): 909–922.
[11] Hnisz D, Schuijers J, Lin CY, Weintraub AS, Abraham BJ, Lee TI, Bradner JE, Young RA. Convergence of developmental and oncogenic signaling pathways at transcriptional super-enhancers.,2015, 58(2): 362–370.
[12] Saint-André V, Federation AJ, Lin CY, Abraham BJ, Reddy J, Lee TI, Bradner JE, Young RA. Models of human core transcriptional regulatory circuitries.,2016, 26(3): 385–396.
[13] Drier Y, Cotton MJ, Williamson KE, Gillespie SM, Ryan RJ, Kluk MJ, Carey CD, Rodig SJ, Sholl LM, Afrogheh AH, Faquin WC, Queimado L, Qi J, Wick MJ, El-Naggar AK, Bradner JE, Moskaluk CA, Aster JC, Knoechel B, Bernstein BE. An oncogenic MYB feedback loop drives alternate cell fates in adenoid cystic carcinoma.,2016, 48(3): 265–272.
[14] Adam RC, Fuchs E. The Yin and Yang of chromatin dynamics in stem cell fate selection.,2016, 32(2): 89–100.
[15] Kaufman CK, Mosimann C, Fan ZP, Yang S, Thomas AJ, Ablain J, Tan JL, Fogley RD, van Rooijen E, Hagedorn EJ, Ciarlo C, White RM, Matos DA, Puller AC, Santoriello C, Liao EC, Young RA, Zon LI. A zebrafish melanoma model reveals emergence of neural crest identity during melanoma initiation.,2016, 351(6272): aad2197.
[16] Witte S, Bradley A, Enright AJ, Muljo SA. High-density P300 enhancers control cell state transitions.,2015, 16: 903.
[17] Lovén J, Hoke HA, Lin CY, Lau A, Orlando DA, Vakoc CR, Bradner JE, Lee TI, Young RA. Selective inhibition of tumor oncogenes by disruption of super-enhancers.,2013, 153(2): 320–334.
[18] Chapuy B, McKeown MR, Lin CY, Monti S, Roemer MG, Qi J, Rahl PB, Sun HH, Yeda KT, Doench JG, Reichert E, Kung AL, Rodig SJ, Young RA, Shipp MA, Bradner JE. Discovery and characterization of super-enhancer-associated dependencies in diffuse large B cell lymphoma.,2013, 24(6): 777–790.
[19] Herranz D, Ambesi-Impiombato A, Palomero T, Schnell SA, Belver L, Wendorff AA, Xu L, Castillo-Martin M, Llobet-Navás D, Cordon-Cardo C, Clappier E, Soulier J, Ferrando AA. A NOTCH1-driven MYC enhancer promotes T cell development, transformation and acute lymphoblastic leukemia.,2014, 20(10): 1130–1137.
[20] Mansour MR, Abraham BJ, Anders L, Berezovskaya A, Gutierrez A, Durbin AD, Etchin J, Lawton L, Sallan SE, Silverman LB, Loh ML, Hunger SP, Sanda T, Young RA, Look AT. Oncogene regulation. An oncogenic super-enhancerformed through somatic mutation of a noncoding intergenic element.,2014, 346(6215): 1373–1377.
[21] Sengupta D, Kannan A, Kern M, Moreno MA, Vural E, Stack B, Jr., Suen JY, Tackett AJ, Gao L. Disruption of BRD4 at H3K27Ac-enriched enhancer region correlates with decreased c-Myc expression in Merkel cell carcinoma.,2015, 10(6): 460–466.
[22] Pelish HE, Liau BB, Nitulescu, II, Tangpeerachaikul A, Poss ZC, Da Silva DH, Caruso BT, Arefolov A, Fadeyi O, Christie AL, Du K, Banka D, Schneider EV, Jestel A, Zou G, Si C, Ebmeier CC, Bronson RT, Krivtsov AV, Myers AG, Kohl NE, Kung AL, Armstrong SA, Lemieux ME, Taatjes DJ, Shair MD. Mediator kinase inhibition further activates super-enhancer-associated genes in AML.,2015, 526(7572): 273–276.
[23] Johnatty SE, Tyrer JP, Kar S, Beesley J, Lu Y, Gao B, Fasching PA, Hein A, Ekici AB, Beckmann MW, Lambrechts D, Van Nieuwenhuysen E, Vergote I, Lambrechts S, Rossing MA, Doherty JA, Chang-Claude J, Modugno F, Ness RB, Moysich KB, Levine DA, Kiemeney LA, Massuger LF, Gronwald J, Lubinski J, Jakubowska A, Cybulski C, Brinton L, Lissowska J, Wentzensen N, Song H, Rhenius V, Campbell I, Eccles D, Sieh W, Whittemore AS, McGuire V, Rothstein JH, Sutphen R, Anton-Culver H, Ziogas A, Gayther SA, Gentry-Maharaj A, Menon U, Ramus SJ, Pearce CL, Pike MC, Stram DO, Wu AH, Kupryjanczyk J, Dansonka- Mieszkowska A, Rzepecka IK, Spiewankiewicz B, Goodman MT, Wilkens LR, Carney ME, Thompson PJ, Heitz F, du Bois A, Schwaab I, Harter P, Pisterer J, Hillemanns P, Group AGOS, Karlan BY, Walsh C, Lester J, Orsulic S, Winham SJ, Earp M, Larson MC, Fogarty ZC, Hogdall E, Jensen A, Kjaer SK, Fridley BL, Cunningham JM, Vierkant RA, Schildkraut JM, Iversen ES, Terry KL, Cramer DW, Bandera EV, Orlow I, Pejovic T, Bean Y, Hogdall C, Lundvall L, McNeish I, Paul J, Carty K, Siddiqui N, Glasspool R, Sellers T, Kennedy C, Chiew YE, Berchuck A, MacGregor S, Pharoah PD, Goode EL, deFazio A, Webb PM, Chenevix-Trench G, Australian Ovarian Cancer Study G. Genome-wide analysis identifies novel loci associated with ovarian cancer outcomes: findings from the ovarian cancer association consortium.,2015, 21(23): 5264–5276.
[24] Zhang X, Choi PS, Francis JM, Imielinski M, Watanabe H, Cherniack AD, Meyerson M. Identification of focally amplified lineage-specific super-enhancers in human epithelial cancers.,2016, 48(2): 176–182.
[25] Yang H, Schramek D, Adam RC, Keyes BE, Wang P, Zheng D, Fuchs E. ETS family transcriptional regulators drive chromatin dynamics and malignancy in squamous cell carcinomas.,2015, 4: e10870.
[26] Wang Y, Zhang T, Kwiatkowski N, Abraham BJ, Lee TI, Xie S, Yuzugullu H, Von T, Li H, Lin Z, Stover DG, Lim E, Wang ZC, Iglehart JD, Young RA, Gray NS, Zhao JJ. CDK7-dependent transcriptional addiction in triple-negative breast cancer.,2015, 163(1): 174–186.
[27] Jiang YY, Lin DC, Mayakonda A, Hazawa M, Ding LW, Chien WW, Xu L, Chen Y, Xiao JF, Senapedis W, Baloglu E, Kanojia D, Shang L, Xu X, Yang H, Tyner JW, Wang MR, Koeffler HP. Targeting super-enhancer-associated oncogenes in oesophageal squamous cell carcinoma.,2017, 66(8): 1358–1368.
[28] Togel L, Nightingale R, Chueh AC, Jayachandran A, Tran H, Phesse T, Wu R, Sieber OM, Arango D, Dhillon AS, Dawson MA, Diez-Dacal B, Gahman TC, Filippakopoulos P, Shiau AK, Mariadason JM. Dual targeting of bromodomain and extraterminal domain proteins, and WNT or MAPK signaling, inhibits c-MYC expression and proliferation of colorectal cancer cells.,2016, 15(6): 1217–1226.
[29] Hanahan D, Weinberg RA. The hallmarks of cancer.2000, 100(1): 57–70.
[30] Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation.,2011, 144(5): 646–674.
[31] Suzuki HI, Young RA, Sharp PA. Super-enhancer- mediated RNA processing revealed by integrative microRNA network analysis.,2017, 168(6): 1000–1014 e15.
[32] Xie JJ, Jiang YY, Jiang Y, Li CQ, Chee LM, An O, Mayakonda A, Ding LW, Long L, Sun C, Lin LH, Chen L, Wu JY, Wu ZY, Cao Q, Fang WK, Yang W, Meltzer SJ, Yang H, Fullwood M, Xu LY, Li EM, Lin DC, Koeffler HP. Super-enhancer-driven long non-coding RNA LINC01503, regulated by TP63, is over-expressed and oncogenic in squamous cell carcinoma.,2018, 154(8): 2137–2151.e1.
[33] Jiao W, Chen Y, Song H, Li D, Mei H, Yang F, Fang E, Wang X, Huang K, Zheng L, Tong Q. HPSE enhancer RNA promotes cancer progression through driving chromatin looping and regulating hnRNPU/p300/EGR1/HPSE axis.,2018, 37(20): 2728–2745.
[34] Cheng X, Yang Q, Tan ZD, Tan Y, Pu H, Zhao X, Zhang SH, Zhu L. The current research status of enhancer RNAs.,2017, 39(9):784–797.程霄, 杨琼, 谭镇东, 谭娅, 蒲红州, 赵雪, 张顺华, 朱砺. 增强子RNA研究现状. 遗传, 2017, 39(9): 784–797.
[35] Lin CY, Erkek S, Tong Y, Yin L, Federation AJ, Zapatka M, Haldipur P, Kawauchi D, Risch T, Warnatz HJ, Worst BC, Ju B, Orr BA, Zeid R, Polaski DR, Segura-Wang M, Waszak SM, Jones DT, Kool M, Hovestadt V, Buchhalter I, Sieber L, Johann P, Chavez L, Groschel S, Ryzhova M, Korshunov A, Chen W, Chizhikov VV, Millen KJ, Amstislavskiy V, Lehrach H, Yaspo ML, Eils R, Lichter P, Korbel JO, Pfister SM, Bradner JE, Northcott PA. Active medulloblastoma enhancers reveal subgroup-specific cellular origins.,2016, 530(7588): 57–62.
[36] Oldridge DA, Wood AC, Weichert-Leahey N, Crimmins I, Sussman R, Winter C, McDaniel LD, Diamond M, Hart LS, Zhu S, Durbin AD, Abraham BJ, Anders L, Tian L, Zhang S, Wei JS, Khan J, Bramlett K, Rahman N, Capasso M, Iolascon A, Gerhard DS, Guidry Auvil JM, Young RA, Hakonarson H, Diskin SJ, Look AT, Maris JM. Genetic predisposition to neuroblastoma mediated by a LMO1 super-enhancer polymorphism.,2015, 528(7582): 418–421.
[37] Super-enhancers facilitate gene regulation by signaling pathways.,2015, 5(5): OF11.
[38] Nabet B, P óB, Reyes JM, Shieh K, Lin CY, Will CM, Popovic R, Ezponda T, Bradner JE, Golden AA, Licht JD. Deregulation of the ras-erk signaling axis modulates the enhancer landscape.,2015, 12(8):1300–1313.
[39] Adelman K, Lis JT. Promoter-proximal pausing of RNA polymerase II: emerging roles in metazoans.,2012, 13(10): 720–731.
[40] Galli GG, Carrara M, Yuan WC, Valdes-Quezada C, Gurung B, Pepe-Mooney B, Zhang T, Geeven G, Gray NS, de Laat W, Calogero RA, Camargo FD. YAP drives growth by controlling transcriptional pause release from dynamic enhancers.,2015, 60(2): 328–337.
[41] Yimlamai D, Fowl BH, Camargo FD. Emerging evidence on the role of the Hippo/YAP pathway in liver physiology and cancer.,2015, 63(6): 1491–1501.
[42] Wang H, Zang C, Taing L, Arnett KL, Wong YJ, Pear WS, Blacklow SC, Liu XS, Aster JC. NOTCH1-RBPJ complexes drive target gene expression through dynamic interactions with superenhancers.,2014, 111(2): 705–710.
[43] Sur I, Taipale J. The role of enhancers in cancer.,2016, 16(8): 483–493.
[44] Vaharautio A, Taipale J. Cancer. Cancer by super-enhancer.,2014, 346(6215): 1291–1292.
[45] Lin CY, Loven J, Rahl PB, Paranal RM, Burge CB, Bradner JE, Lee TI, Young RA. Transcriptional amplification in tumor cells with elevated c-Myc.,2012, 151(1): 56–67.
[46] Didiasova M, Schaefer L, Wygrecka M. Targeting GLI transcription factors in cancer.,2018, 23(5): 1003.
[47] Nilson KA, Guo J, Turek ME, Brogie JE, Delaney E, Luse DS, Price DH. THZ1 reveals roles for Cdk7 in co-transcriptional capping and pausing.,2015, 59(4): 576–587.
[48] Di Micco R, Fontanals-Cirera B, Low V, Ntziachristos P, Yuen SK, Lovell CD, Dolgalev I, Yonekubo Y, Zhang G, Rusinova E, Gerona-Navarro G, Canamero M, Ohlmeyer M, Aifantis I, Zhou MM, Tsirigos A, Hernando E. Control of embryonic stem cell identity by BRD4-dependent transcriptional elongation of super-enhancer-associated pluripotency genes.,2014, 9(1): 234–247.
[49] Bartkowiak B, Liu P, Phatnani HP, Fuda NJ, Cooper JJ, Price DH, Adelman K, Lis JT, Greenleaf AL. CDK12 is a transcription elongation-associated CTD kinase, the metazoan ortholog of yeast Ctk1.,2010, 24(20):2303–2316.
[50] Liang K, Gao X, Gilmore JM, Florens L, Washburn MP, Smith E, Shilatifard A. Characterization of human cyclin- dependent kinase 12 (CDK12) and CDK13 complexes in C-terminal domain phosphorylation, gene transcription, and RNA processing.,2015, 35(6): 928–938.
[51] Shin HY. Targeting super-enhancers for disease treatment and diagnosis.,2018, 41(6): 506–514.
[52] Bai L, Zhou B, Yang CY, Ji J, McEachern D, Przybranowski S, Jiang H, Hu J, Xu F, Zhao Y, Liu L, Fernandez-Salas E, Xu J, Dou Y, Wen B, Sun D, Meagher J, Stuckey J, Hayes DF, Li S, Ellis MJ, Wang S. Targeted degradation of BET proteins in triple-negative breast cancer.,2017, 77(9): 2476–2487.
[53] Odore E, Lokiec F, Cvitkovic E, Bekradda M, Herait P, Bourdel F, Kahatt C, Raffoux E, Stathis A, Thieblemont C, Quesnel B, Cunningham D, Riveiro ME, Rezai K. Phase I population pharmacokinetic assessment of the oral bromodomain inhibitor OTX015 in patients with haematologic malignancies., 2016,3(55): 397–405.
[54] Berthon C, Raffoux E, Thomas X, Vey N, Gomez-Roca C, Yee K, Taussig DC, Rezai K, Roumier C, Herait P, Kahatt C, Quesnel B, Michallet M, Recher C, Lokiec F, Preudhomme C, Dombret H. Bromodomain inhibitor OTX015 in patients with acute leukaemia: a dose- escalation, phase 1 study., 2016, 3 (4):186–195 .
[55] Chipumuro E, Marco E, Christensen CL, Kwiatkowski N, Zhang T, Hatheway CM, Abraham BJ, Sharma B, Yeung C, Altabef A, Perez-Atayde A, Wong KK, Yuan GC, Gray NS, Young RA, George RE. CDK7 inhibition suppresses super-enhancer-linked oncogenic transcription in MYCN- driven cancer.,2014, 159(5): 1126–1139.
[56] Wong RWJ, Ngoc PCT, Leong WZ, Yam AWY, Zhang T, Asamitsu K, Iida S, Okamoto T, Ueda R, Gray NS, Ishida T, Sanda T. Enhancer profiling identifies critical cancer genes and characterizes cell identity in adult T-cell leukemia., 2017, 130(21):2326–2338.
[57] Zhang T, Kwiatkowski N, Olson CM, Dixon-Clarke SE, Abraham BJ, Greifenberg AK, Ficarro SB, Elkins JM, Liang Y, Hannett NM, Manz T, Hao M, Bartkowiak B, Greenleaf AL, Marto JA, Geyer M, Bullock AN, Young RA, Gray NS. Covalent targeting of remote cysteine residues to develop CDK12 and CDK13 inhibitors.,2016, 12(10):876–884.
[58] Kennedy AL, Vallurupalli M, Chen L, Crompton B, Cowley G, Vazquez F, Weir BA, Tsherniak A, Parasuraman S, Kim S, Alexe G, Stegmaier K. Functional, chemical genomic, and super-enhancer screening identify sensitivity to cyclin D1/CDK4 pathway inhibition in Ewing sarcoma.,2015, 6(30): 30178–30193.
[59] Delmore JE, Issa GC, Lemieux ME, Rahl PB, Shi J, Jacobs HM, Kastritis E, Gilpatrick T, Paranal RM, Qi J, Chesi M, Schinzel AC, McKeown MR, Heffernan TP, Vakoc CR, Bergsagel PL, Ghobrial IM, Richardson PG, Young RA, Hahn WC, Anderson KC, Kung AL, Bradner JE, Mitsiades CS. BET bromodomain inhibition as a therapeutic strategy to target c-Myc.,2011, 146(6): 904–917.
[60] Filippakopoulos P, Qi J, Picaud S, Shen Y, Smith WB, Fedorov O, Morse EM, Keates T, Hickman TT, Felletar I, Philpott M, Munro S, McKeown MR, Wang Y, Christie AL, West N, Cameron MJ, Schwartz B, Heightman TD, La Thangue N, French CA, Wiest O, Kung AL, Knapp S, Bradner JE. Selective inhibition of BET bromodomains.,2010, 468(7327): 1067–1073.
[61] Amorim S, Stathis A, Gleeson M, Iyengar S, Magarotto V, Leleu X, Morschhauser F, Karlin L, Broussais F, Rezai K, Herait P, Kahatt C, Lokiec F, Salles G, Facon T, Palumbo A, Cunningham D, Zucca E, Thieblemont C. Bromodomain inhibitor OTX015 in patients with lymphoma or multiple myeloma: a dose-escalation, open-label, pharmacokinetic, phase 1 study.,2016, 3(4): e196–204.
[62] Winter GE, Buckley DL, Paulk J, Roberts JM, Souza A, Dhe-Paganon S, Bradner JE. DRUG DEVELOPMENT. Phthalimide conjugation as a strategy for in vivo target protein degradation.,2015, 348(6241): 1376– 1381.
[63] Tasdemir N, Banito A, Roe JS, Alonso-Curbelo D, Camiolo M, Tschaharganeh DF, Huang CH, Aksoy O, Bolden JE, Chen CC, Fennell M, Thapar V, Chicas A, Vakoc CR, Lowe SW. BRD4 connects enhancer remodeling to senescence immune surveillance.,2016, 6(6): 612–629.
[64] Kwiatkowski N, Zhang T, Rahl PB, Abraham BJ, Reddy J, Ficarro SB, Dastur A, Amzallag A, Ramaswamy S, Tesar B, Jenkins CE, Hannett NM, McMillin D, Sanda T, Sim T, Kim ND, Look T, Mitsiades CS, Weng AP, Brown JR, Benes CH, Marto JA, Young RA, Gray NS. Targeting transcription regulation in cancer with a covalent CDK7 inhibitor.,2014, 511(7511): 616–620.
Research progress of super enhancer in cancer
Zhiqiang Wu1,2, Zeyun Mi3
Super enhancers (SEs) are composed of clusters of enhancers in close genomic proximity. They constitute a large family of regulatory elements that specify gene expression patterns and cell identity. SE regions consist of unusually strong enrichment of binding sites for transcriptional factors, cofactors, and enhancers associated with epigenetic modifications. SEs play important roles in regulating the aberrant gene expression in tumor cells.SEs, cancer cells activate the expression of various oncogenes, and promote cell proliferation, invasion and migration properties. Hence suppression of SEs activities could inhibit the growth and survival of cancer cells. In this review, we summarize the fundamental principles, functions and regulation of super enhancers and therapeutic potential in targeting SEs in cancer cells, thereby introducing and providing new conceptions for development of antineoplastic drugs.
enhancer; super enhancer; transcription; cancer
2018-07-13;
2018-09-21
国家自然科学基金项目(编号:81502660,31700144)和天津市自然科学基金项目(编号:16JCQNJC10000, 17JCQNJC10600)资助[Supported by the National Natural Science Foundation of China (Nos. 81502660,31700144) and Tianjin Municipal Science and Technology Commission (Nos. 16JCQNJC10000, 17JCQNJC10600)]
吴志强,博士,讲师,研究方向:放射生物学,肿瘤分子生物学。E-mail: zwu08@tmu.edu.cn
米泽云,博士,讲师,研究方向:肿瘤分子生物学。E-mail: mizeyun@tmu.edu.cn
10.16288/j.yczz.18-152
2018/11/6 17:30:31
URI: http://kns.cnki.net/kcms/detail/11.1913.R.20181106.1730.003.html
(责任编委: 方向东)
猜你喜欢
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
中国病理生理杂志(2018年6期)2018-01-22
中华老年口腔医学杂志(2016年3期)2017-01-15
作文教学研究(2016年1期)2016-07-05
中国男科学杂志(2016年9期)2016-03-20
原子与分子物理学报(2015年1期)2015-11-24
中国医疗美容(2015年1期)2015-07-12
医学研究杂志(2015年9期)2015-07-01
医学研究杂志(2015年8期)2015-06-22
