
蚕豆赤斑病抗性的主基因+多基因遗传分析
2019-02-10 10:50杜成章龙珏臣龚万灼朱振东宗绪晓张继君
植物保护 2019年6期
关键词:蚕豆
杜成章 龙珏臣 龚万灼 朱振东 宗绪晓 张继君


摘要 :赤斑病是世界蠶豆产区的主要病害,严重威胁蚕豆安全生产。为明确蚕豆赤斑病的抗性遗传规律,本研究用赤斑病抗性较好的蚕豆品种‘通蚕鲜8号、‘启豆2号分别与高感赤斑病蚕豆品种‘成胡10号、‘成胡14号配制杂交组合,采用主基因+多基因混合遗传模型方法对2个组合6世代(P1、P2、F1、F2、BCP1、BCP2)的赤斑病抗性进行了遗传分析。结果表明,蚕豆对赤斑病的抗性最适合遗传模型为E 0(两对加性显性上位性主基因+加性显性上位性多基因)。两对赤斑病抗性的主基因加性效应值在2个组合中分别为-40.43、2.16和-36.31、-3.86,显性效应值分别为-15.22、-15.72和-5.98、-6.48。2个组合的主基因遗传率在BCP1、BCP2、F2中分别是19.05%、51.99%、70.90%和19.29%、52.13%、77.35%,多基因遗传率分别为0、0、19.9%和0、0、21.06%。本试验条件下,蚕豆品种‘通蚕鲜8号、‘启豆2号对赤斑病抗性由2个主效基因控制,同时受多基因修饰作用,环境对其抗性影响较小。感病亲本对后代抗病性的负向影响较大,在育种实践中需适当提高感病亲本的抗病性,以提高后代的抗性水平。
关键词 :蚕豆; 赤斑病; 主基因+多基因混合遗传模型; 抗性遗传
中图分类号:
S 435.23
文献标识码: A
DOI: 10.16688/j.zwbh.2018444
Analysis of major genes plus polygenes mixed inheritance for
resistance to chocolate spot on faba bean
DU Chengzhang1, LONG Juechen1, GONG Wanzhuo1, ZHU Zhendong2, ZONG Xuxiao2, ZHANG Jijun1
(1. Research Institute of Specialty Crops, Chongqing Academy of Agricultural Sciences, Chongqing 402160, China;
2. Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China)
Abstract
Chocolate spot is one of the main diseases in the faba bean production areas in the world, which seriously threatens the production of faba bean. To confirm the inheritance of the resistance of faba bean to chocolate spot, the chocolate spot highly resistant varieties ‘Tongcanxian No.8 and ‘Qidou No.2 were crossed with the highly susceptible varieties ‘Chenghu No.10 and ‘Chenghu No.14, and the major gene plus polygene mixed genetic model was used to conduct a genetic analysis of the 2 cross combinations for 6 generations (P1, P2, F1, F2, BCP1, BCP2). The results indicated that the most suitable genetic model of faba bean chocolate spot resistance is E 0 (two addictive dominant epistatic major gene + addictive dominant epistatic polygene). Additive effect values of the two major chocolate spot resistance genes in the above two combinations were -40.43, 2.16 and -3631, -3.86, and the dominant effect values were -15.22, -15.72 and -5.98, -6.48, respectively. The inheritability of major genes in the above two combinations was 19.05%, 51.99%, 70.90% and 19.29%, 5213%, 77.35% in BCP1, BCP2 and F2, and the inheritability of polygene was 0, 0, 19.9% and 0, 0, 21.06%, respectively. Therefore, chocolate spot resistance in faba bean might be controlled by two major genes, and modified by polygenes at the same time, while the environment had less impact on it. The susceptible parents had a negative effect on their derived generations resistance. In the breeding practice, it is required to improve the resistance of susceptible parents so as to improve the next generations disease resistance.
Key words
faba bean; chocolate spot; major gene plus polygene mixed genetic model method; resistance breeding
目前蚕豆在55个国家种植,面积约220万hm2,年产量约420万t[1]。中国蚕豆生产面积和产量约占全球总量的一半[2]。世界蚕豆种植面积正在减小[3],其原因之一是病虫害引起的蚕豆产量不稳[4]。赤斑病是世界蚕豆产区的主要病害[5],在埃及、法国、马格里布等国家和地区[67]以及中国部分蚕豆产区[89],尤其是中国南方高降水地区发生特别严重。
灰葡萄孢Botrytis cinerea Pers. et Fr.和蚕豆葡萄孢Botrytis fabae Sardina是引起赤斑病的主要病原菌[10]。轮作、推迟种植期以及使用杀菌剂[11]等农艺措施对赤斑病只能起到部分防控作用,开发抗性品种才是该病害最有效的控制方法[1213]。在20世纪80年代,国际干旱地区农业研究中心和该病流行的主要蚕豆生产国将培养抗赤斑病蚕豆品种列为主要育种目标之一[14]。但是,抗性育种工作进展并不顺利,仅有1个中抗赤斑病的蚕豆新品种‘通蚕鲜8号通过鉴定并得到推广[1516]。
由于国内外对赤斑病抗性遗传基础研究报道较少,抗病遗传机制尚不明确,这在很大程度上制约了抗病育种工作的开展。近年来,在重庆市农业科学院的蚕豆赤斑病抗性育种工作中,杂交后代抗性往往低于抗性亲本,却高于感病亲本,很难选择出抗性新种质。分析蚕豆对赤斑病抗性遗传规律、鉴定抗性基因、创造新的抗性种质资源,是提高蚕豆品种抗病能力的有效途径。本文采用主基因+多基因混合遗传模型分析方法研究蚕豆对赤斑病的抗性遗传规律,探索抗性主基因与多基因的作用及其相对重要性,为拓宽抗病品种的遗传基础,培育抗病品种提供依据。
1 材料与方法
1.1 材料
2009年、2010年连续两年对来自不同遗传背景下的国内外2 136份蚕豆资源,通过田间鉴定,根据其赤斑病发病程度筛选出赤斑病抗性品种‘通蚕鲜8号和‘启豆2号以及感病品种‘成胡10号和‘成胡14号。
1.2 方法
1.2.1 试验设计
试验于重庆市渝西作物试验站进行,2006年至今该田块已经连续多年高发蚕豆赤斑病。2011年以感病品种P1(‘成胡10号)为母本(S1)和抗性品种P2(‘通蚕鲜8号)为父本(MR1)、感病品种P1(‘成胡14号)为母本(S2)和抗性品种P2(‘启豆2号)为父本(MR2)组配两个杂交组合[7],获得F1(S1×MR1)和F1(S2×MR2)种子。2012年分别用双亲回交和自交,2个组合分别得到BCP1(S1/MR1∥S1)、BCP2(S1/MR1∥MR1)、F2(S1×MR1)群体和BCP1(S2/MR2∥S2)、BCP2(S2/MR2∥MR2)、F2(S2×MR2);从而获得两个杂交组合的6个基本世代群体,即P1、P2、F1、F2、BCP1、BCP2群体。2013年种植6个世代进行抗赤斑病鉴定,采用随机区组设计,3次重复。P1、P2、F1设为10行区,BCP1、BCP2、F2设为20行区。小区行长20 m,行距40 cm,窝距25 cm,每窝留苗2株。采用田间自然发病,待鉴定材料每播种2行再播种1行‘成胡10号(HS)作为感染源。2013年10月27日播种,常规田间管理。
1.2.2 发病程度鉴定
待赤斑病充分发生时,即盛荚期进行病情调查,S1、S2、F1、BCP1、BCP2实际调查株数都为60株,F2实际调查株数为120株,每株分上部、中上部、中部、中下部、下部5个部分调查,每部分随机调查10片小叶,发病程度以病斑面积占叶面积的百分数表示。为了提高数据分析的精确性,本文将发病程度划分为11个级别,分级标准如下。L0为叶片干净无病斑;L1为叶片上仅有少数针尖状病斑,病斑占叶面积≤10%;L2为叶片上病斑散生,10%<病斑占叶面积≤20%;L3为叶片上病斑散生,20%<病斑占叶面积≤30%;L4为叶片上病斑散生,30%<病斑占叶面积≤40%,有少量落叶;L5为叶片上病斑多散生,40%<病斑占叶面积≤50%,有少量落叶;L6为叶片上病斑多散生,50%<病斑占叶面积≤60%,有少量落叶;L7为叶片和荚果上病斑多且大,60%<病斑占叶面积≤70%,有半數叶片枯死或脱落;L8为叶片和荚果上病斑多且大,70%<病斑占叶面积≤80%,有半数以上叶片枯死或脱落;L9为叶片和荚果上病斑多且大,80%<病斑占叶面积≤90%,绝大多数叶片枯死或脱落;L10为植株发黑死亡,90%<病斑占叶面积≤100%。
1.2.3 数据统计分析
利用 DPS 15.10 统计分析软件进行基本统计量、相关系数及多组数据差异显著性比较。应用主基因+多基因混合遗传模型的多世代联合分析法[17],并结合植物数量性状分离分析(SEA)[18]软件,进行数据处理和遗传模型分析。通过极大似然分析和迭代最大期望算法 (iterated expectation and conditional maximization,IECM) 对混合分布中的相关参数作出估计。采用最小赤池信息量准则(Akaikes information criterion,AIC),选取AIC 值最小及与最小AIC 值比较接近的遗传模型作为备选模型,并进行适合性检验,包括均匀性检验U21、U22、U23,Smirnov检验nW2和Kolmogorov检验Dn,根据达到显著水平统计量最少的原则,检验结果确定最佳遗传模型,并用最小二乘法估计出相应的基因效应值、方差及相关遗传参数。群体的多基因方差σ2pg=σ2p-σ2mg-σ2,主基因遗传率h2mg=σ2mg/σ2p×100%,多基因遗传率h2pg=σ2pg/σ2p×100%,其中σ2mg、σ2pg、σ2p和σ2分别表示主基因方差、多基因方差、表型方差和环境方差。
2 结果与分析
2.1 发病程度数据分析
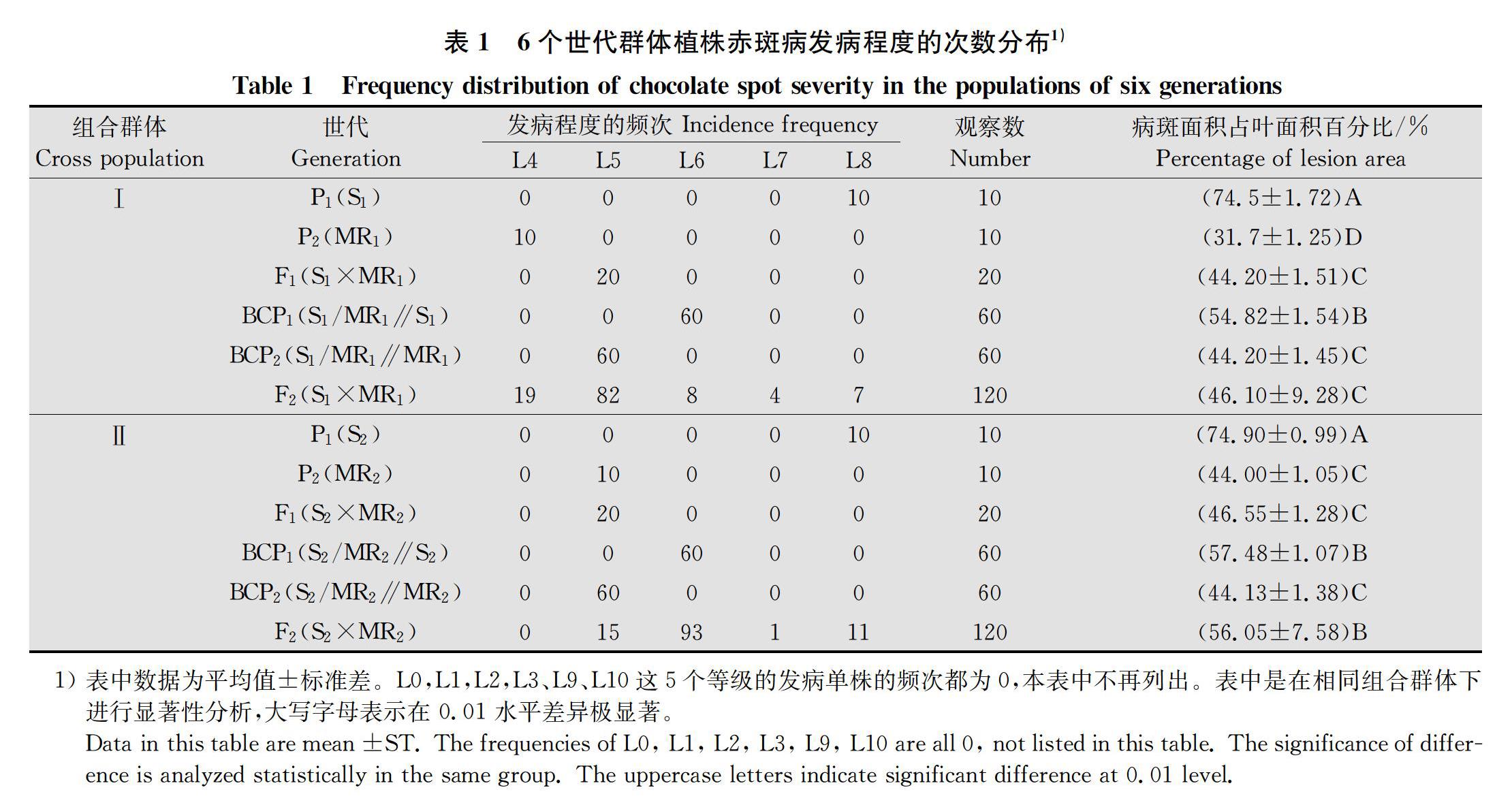
由表2可见,两个组合群体的感病亲本 P1都在本组6个世代群体中发病程度最重,极显著高于抗病品种P2。2个组合群体的2个回交组合BCP1、BCP2的平均发病程度,均介于本组合2个亲本之间,2个组合的回交群体BCP1平均发病程度均低于P1,高于P2,高于双亲的平均值,BCP2则低于双亲的平均值。2个组合的F1和F2群体平均发病等级都低于本组合双亲的平均值。2个组合的BCP1和BCP2均表现为偏态性不连续分布,BCP1偏向感病亲本P1,BCP2偏向抗病亲本P2,F2病情呈连续分布且显示多峰,说明2个抗性亲本的抗赤斑病的遗传明显地表现出主基因+多基因遗传特征。
2.2 蚕豆赤斑病抗性遗传模型的确定和检验
利用植物数量性状主基因+多基因混合遗传模型的6世代(P1、P2、F1、BCP1、BCP2、F2)联合分析方法分析,分别获得1对主基因(A类模型)、2对主基因(B类模型)、多基因(C类模型)、1对主基因+多基因(D类模型)和2对主基因+多基因(E类模型)等5类24种遗传模型的极大对数似然函数值和AIC值(表2)。根据最小AIC值原则,从各种模型中初步确定2个组合的备选遗传模型均为D 1、D 2、D 4和E 0模型。
进一步对备选模型进行适合性检验(均匀性检验、Smirnov检验和Kolmogorov检验的5个统计量U21、U22、U23、nW2和Dn),选择统计量达到显著水平个数较少的模型作为最佳模型。备选模型的适合性测验结果表明(表3),在组合Ⅰ(S1×MR1)中,E 0模型有5项统计量达到显著或极显著差异,D 1、D 2、D 4模型都有11项统计量达到显著或极显著差异,因此E 0模型是该组合的最佳模型。在组合Ⅱ(S2×MR2)中,E 0 模型有 7项达到显著或极显著差异,D 1、D 2、D 4模型都有11项统计量达到显著或极显著差异,因此也是E 0模型是该组合的最佳模型。两年份的最优遗传模型均是 E 0模型,即两对加性 显性 上位性主基因+加性 显性 上位性多基因模型。
2.3 蚕豆赤斑病抗性遗传模型的参数估计
利用最小二乘法对获得的最适遗传模型(E 0模型)估计一阶遗传参数和二阶遗传参数(表4、表5)。结果表明,2个组合,控制蚕豆抗赤斑病的2对主基因的加性效应分别为-40.43、2.16和-36.31、-3.86,只有组合Ⅰ的第2主基因的加性效应为正值,且数值相对较小,其余都为负值,这说明加性效应以负向效应为主,即感病亲本对后代抗病性的负向影响比较大;2个基因的显性效应分别为-15.22、-5.98和-15.72、-6.48,加性基因的显性效应均为负值,也验证了感病亲本对后代抗病性的负向影响较大。同时,2个组合的|ha/da︱<1,|hb/db︱>1说明控制赤斑病抗性的第1主基因以加性效应为主,第2主基因以显性效应为主。
此外,2对主基因间加性×加性效应i都为负值有利于抗病,而显性×显性效应l都为正值,不利于抗病。第1主基因的加性效应与第2主基因的显性效应间互作效应jab分别为7.85与1.89,而第1主基因的显性效应与第2主基因的加性效应间互作效应jba分别为-0.12与5.99,对抗病几乎无有利影响,且作用相对较小。可见,在赤斑病抗性的遗传中,两主基因的加性效应、显性效应起到了重要作用,上位性效应作用相对较小。
2.4 蚕豆赤斑病抗性遗传模型的抗性分析
从二阶遗传参数来看,2个组合的蚕豆赤斑病抗性BCP1、BCP2和F2分离世代群体主基因遗传率分别为19.05%、51.99%、70.90%和19.29%、52.13%、77.35%,多基因的遗传率分别为0.00%、0.00%、19.90%和0.00%、0.00%、21.06%,各世代的主基因遗传率都大于相应世代的多基因遗传率,可见在蚕豆赤斑病抗性遗传体系中,主效基因的遗传贡献占主要部分,同时受多基因部分修饰作用。环境方差仅占总表型方差的1.6%~9.2%,环境对其抗性存在影响较小。
3 讨论
制约蚕豆赤斑病抗性育种的主要因素是缺乏有效的抗性种质,已经报道的抗性种质主要来自哥伦比亚、厄瓜多尔、中国、意大利、约旦、黎巴嫩、西班牙,但大多未进入市场[3,19],本文所选用的抗性种质与感病种质都是市场化比较成熟的品种,而且常被中国的育种家当做杂交亲本利用,因此,本文所形成的结论对今后中国蚕豆抗性育种具有指导意义。
本文将蚕豆赤斑病发病程度划分为11个等级,与传统的6个等级鉴定方法[7,20]相比,该方法对蚕豆赤斑病发病程度的描述更加精准,能够将传统方法难以界定的“中间型”重新分类,进而提高了分析结果的准确性。
本文所选的两个组合BCP1和BCP2对赤斑病的抗性均表现为偏态性不连续分布,F2病情呈连续分布且显示多峰,选用主基因+多基因模型进行遗传模型和效应分析是恰当的。本研究结果表明,蚕豆赤斑病抗性遗传符合两对加性 显性 上位性主基因+加性 显性 上位性多基因(E 0)模型,主效基因的遗传贡献占主要部分,且两主基因的加性效应、显性效应起到了重要作用,多基因起到部分修饰作用。这与Noorka等[22]研究结论不一致,他们认为蚕豆赤斑病抗性具有单基因显性遗传的特征。本文的结论与Beyene等[22]的研究結果相近,他们认为蚕豆赤斑病抗性属于数量性状遗传,存在单个或多个加性效应基因控制。
Beyene等[23]的研究结论显示,蚕豆赤斑病抗性具有较高遗传力,可以利用表型选择来改进蚕豆的抗性。而本研究结果显示,两个组合的两对主基因加性和显性效应值几乎全部为负,说明感病亲本对后代抗性影响的效应为负,在抗性亲本与感病亲本的单交(single cross)后代群体中难以选择到优良的抗性种质。因此,在今后的赤斑病抗性育种实践中需适当提高亲本的抗性,还需利用复合杂交、回交等手段提高后代的抗病性。
本研究將‘成胡14用作赤斑病感病亲本,而余东梅等[24]认为‘成胡14是赤斑病抗性种质。但本研究的结果显示,‘成胡14田间发病程度高达74.90%±0.99%,且对后代抗性影响的效应为负值,说明该品种不适合用作蚕豆赤斑病抗性育种的亲本。这种结果的出现的原因可能是由于环境差异造成的。在本研究前期的基础性工作表明,余东梅等所在地区与本试验所在地区相隔约300 km,两地病原物致病性和生理小种均一致,但本试验所在地区的降水量、空气湿度、气温均高于余东梅等所在地区,更容易诱发蚕豆赤斑病,从而导致了鉴定结果的不同。
本研究结果还表明,蚕豆赤斑病抗性主基因的遗传占据主导作用,多基因的遗传率在BCP1、BCP2接近于0,两个组合的主基因和多基因遗传率都在F2表现最大。F2主基因的选择效率最高,建议在今后的蚕豆赤斑病抗性育种实践中,在该世代进行严格选择。
本研究利用主基因+多基因混合遗传模型分析了蚕豆赤斑病抗性的遗传,并检测出两对主基因的存在与效应,同时还分析了微效基因的综合表现。下一步需要将单株产量、表型性状结合分子标记进行遗传定位分析,从分子水平上阐述赤斑病抗性的基因及其遗传调控网络,发掘与赤斑病抗性及产量相关性状紧密连锁的分子标记,实现蚕豆抗赤斑病分子标记辅助育种。
4 结论
蚕豆赤斑病抗性受两对加性 显性 上位性主基因+加性 显性 上位性多基因控制(E 0模型),显性效应值和加性效应值基本同为负向,控制赤斑病抗性的第1对主基因以加性效应为主,第2对主基因以显性效应为主。各世代的主基因遗传率都大于相应世代的多基因遗传率,说明在其后代选育抗病品系,要重点考虑主基因的影响。
参考文献
[1] FAOSTAT. World statistics on faba bean [EB/OL]. Food and Agriculture Organization of the United Nations, Rome. http:∥faostat.fao.org/, 2017.
[2] 王海飞, 关建平, 马钰.中国蚕豆种质资源ISSR标记遗传多样性分析[J].作物学报, 2011, 37(4): 595602.
[3] SILLERO J C, VILLEGAS FERN NDEZ A M, THOMAS J, et al. Faba bean breeding for disease resistance[J]. Field Crops Research, 2010,115(3): 297307.
[4] STODDARD F L, NICHOLAS A H, RUBIALES D, et al. Integrated pest, disease and weed management in faba bean[J]. Field Crops Research, 2010, 115(3):308318.
[5] 王晓鸣, 朱振东, 段灿星. 蚕豆豌豆病虫害鉴别与控制技术[M]. 北京: 中国农业科学技术出版社, 2007: 15.
[6] BOUHASSAN A, SADIKI M, TIVOLI B. Evaluation of a collection of faba bean (Vicia faba L.) genotypes originating from the Maghreb for resistance to chocolate spot (Botrytis fabae) by assessment in the field and laboratory [J]. Euphytica, 2004,135(1): 5562.
[7] TIVOLI B, BARANGER A, AVILA C M, et al. Screening techniques and sources of resistance to foliar diseases caused by major necrotrophic fungi in grain legumes[J]. Euphytica, 2006,147(1/2), 223253.
[8] 王淑英, 柴琦. 甘肃省春蚕豆叶部病害病原鉴定及主要病害[J]. 植物保护学报, 2000,27(2):121125.
[9] 王淑英, 南志标, 刘福.甘肃蚕豆赤斑病及轮斑病的为害分析及经济阈值研究[J]. 植物保护学报, 1997,24(4): 371372.
[10]顾和平, 陈新, 陈华涛, 等. 不同杀菌剂对蚕豆赤斑病防治效果试验[J]. 南方农业学报, 2012, 43(3): 329331.
[11]赵晓武,严瑞飞,张永辉, 等. 赤斑病、褐斑病对蚕豆的为害分析和经济阈值的探讨[J]. 植物保护学报, 1992,19(3):270.
[12]MAALOUF F, AHMED S, SHAABAN K, et al. New faba bean germplasm with multiple resistances to Ascochyta blight, chocolate spot and rust diseases [J]. Euphytica, 2016,211(2): 157167.
[13]EL FIKI I A I. Efficiency of commercial active dry yeast for controlling the faba bean chocolate spot disease, caused by the fungus, Botrytis fabae [J]. Egyptian Journal of Biological Pest Control, 2017, 27(1): 59.
[14]梁训义,周惠静,王政逸.蚕豆种质资源对赤斑病的抗性鉴定与筛选[J]. 浙江大学学报(农业与生命科学版), 1992 (3): 4045.
[15]杜成章, 李艳花, 孟鸿菊, 等. 重庆蚕豆产业可持续发展的思考[J]. 安徽农业科学, 2015, 43(6): 337338.
[16]汪凯华, 王学军, 缪亚梅, 等. 优质鲜食大粒蚕豆通蚕鲜 8 号的选育和栽培要点[J]. 江苏农业科学, 2013, 41(11): 113115.
[17]盖钧镒, 康元明. 植物数量性状遗传体系[M]. 北京:科学出版社, 2003.
[18]曹锡文, 刘兵, 章元明. 植物数量性状分离分析 Windows 软件包 SEA 的研制[J]. 南京农业大学学报, 2013,36(6): 16.
[19]SILLERO J C, AVILA C M, RUBIALES D. Screening faba bean (Vicia faba) for resistance to aphids (Aphis fabae) [C]∥International Conference Advances in Grain Legume Breeding, Cultivations and Uses for a More Competitive Value Chain, 2017.
[20]BERNIER C C, HANOUNIK S B, HUSSEIN M M, et al. Field manual of common faba bean diseases in the Nile Vally. Information Bulletin No. 3 [M]. International Centre for Agricultural Research in the Dry Areas (ICARDA),1993.
[21]NOORKA I R, EL BRAMAWYM M A S. Inheritance assessment of chocolate spot and rust tolerance in mature faba bean (Vicia faba L.) plants [J]. Pakistan Journal of Botany,2011, 43(2): 13891402.
[22]BEYENE A,DERERA J, SIBIYA J, et al. Gene action determining grain yield and chocolate spot (Botrytis fabae) resistance in faba bean [J].Euphytica, 2016, 207(2): 293304.
[23]BEYENE A T, JOHN D, JULIA S,et al. Genetic variability of faba bean genotypes for chocolate spot (Botrytis fabae) resistance and yield [J]. Euphytica, 2018, 214(8):132.
[24]余東梅,唐海涛,吴正基,等.菜用蚕豆新品种“成胡14”的选育与应用[J].农业科技通讯,2008(12):153154.
(责任编辑:田 喆)
猜你喜欢
星火·中短篇小说(2021年3期)2021-04-28
语文世界(小学版)(2020年8期)2020-08-17
科教新报(2020年24期)2020-07-31
阅读(低年级)(2020年4期)2020-06-21
大理文化(2020年3期)2020-06-11
北京广播电视报(2019年20期)2019-07-13
现代养生·上半月(2019年2期)2019-02-24
创新作文(5-6年级)(2018年1期)2018-07-06
人生与伴侣·共同关注(2016年24期)2016-08-23
