
DC靶向适配体修饰的载铜绿假单胞菌DNA疫苗递送系统的构建及体外评价
2019-04-27 01:57黄仕琴何颖娜岳瀚勋
中国药科大学学报 2019年6期
黄仕琴,石 敏,何颖娜,岳瀚勋,余 娴*
(1重庆医科大学附属第二医院I期临床试验研究室,重庆 400010;2河北中医学院药学院,石家庄 050200;3重庆医科大学药学院生物化学与分子药理学重点实验室,重庆 400010)
铜绿假单胞菌(Pseudomonasaeruginosa,PA)是引起医源性感染的常见病原体之一,主要引起烧伤、囊性纤维化、免疫缺陷病人急性或慢性感染[1]。PA引起的感染通常难以治愈,原因在于其对抗生素类药的多重耐药性[2]。多重耐药的PA感染引起的死亡率可达40.6%[3],研究有效的PA疫苗成为预防PA感染的选择之一。PA的外膜蛋白(outer membrane protein,Opr)F或I对小鼠感染模型有明确的免疫保护作用[3-4],且Opr基因在不同血清型高度保守[5]。临床试验结果表明,以OprF/I为候选抗原的疫苗有良好的应用前景[6]。细胞穿膜肽(cell penetrating peptides,CPPs)是促进树突状细胞(dendritic cells,DC)摄取抗原的有效方法,CPP携带抗原易于进入DC,继而促进MHC Ⅰ和MHC Ⅱ类途径提呈抗原,最终增强体液及细胞免疫[7]。1型单纯疱疹病毒的间层蛋白VP22是重要的CPP之一,其能从表达细胞进入周围未表达细胞[8]。前期已成功构建pVAX1-OprF-VP22 DNA疫苗,然而DNA疫苗在体内易被核酸酶降解且非抗原提呈细胞优先摄取导致其免疫效力严重受限是限制其临床应用的最大难题。
近年来,药物靶向递送系统是研究热点,而针对DNA疫苗的缺点,其理想的递送系统需满足两个条件[9]:保护DNA疫苗不被核酸酶降解;靶向递送DNA疫苗至抗原提呈细胞(antigen presenting cell,APC)。因此,筛选一个与靶细胞受体特异性和亲和力高的抗体或配体十分必要。基于核酸的适配体是抗体的非蛋白替代品,其除了具有与抗体相似的高特异性和高亲和力之外,还具有许多优于抗体的特性:相对分子质量小,无毒性及免疫原性,易于体外筛选及合成等[10]。
适配体靶向递送抗原至DC已有相关研究报道[11]。其中,DEC-205是研究最多的DC特异性受体之一,在人、鼠间序列保守[12],主要在皮肤朗格汉斯细胞、脾脏及淋巴结的CD8+DC表面高度表达。DEC-205介导DC对抗原的摄取、加工及提呈[13],靶向DEC-205可使MHC Ⅰ及MHC Ⅱ类途径抗原提呈分别提高1 000及300倍[11]。Wengerter等[14]筛选出鼠DEC-205的RNA适配体min.2,该适配体能显著增强CD8+T细胞的增殖,促进IFN-γ、IL-2的分泌,并抑制表达OVA B16肿瘤细胞的增殖。然而,min.2用于靶向递送DNA疫苗的研究还未见报道,本研究通过制备一种适配体修饰的DC靶向载pVAX1-OprF-VP22 DNA 疫苗的阳离子脂质体并对其进行体外评价,为PA感染的预防提供新的思路。
1 材 料
1.1 药品与试剂
RNA适配体5Cy3-min.2(min.2序列5′标记Cy3)、RNA适配体5S-min.2(min.2 5′标记-SH)(上海生工生物有限公司),序列详述于表1;DOTAP、胆固醇、MPEG-2000-DSPE、DSPE-PEG2000-MAL(上海艾维特有限公司);聚乙烯亚胺(polyethyleninime,PEI,美国Polysciences公司);pEGFP-N2及pVAX1-OprF-VP22由本实验室保存。小鼠骨髓来源的树突状细胞系DC2.4、中国仓鼠卵巢细胞CHO(华拓生物有限公司);MEM细胞培养基、NEAA(美国Gibco公司);DMEM高糖细胞培养基、RPMI 1640细胞培养基、胎牛血清(FBS)、胰蛋白酶及CCK-8试剂盒(赛米克生物有限公司);重组小鼠粒细胞-巨噬细胞集落刺激因子(recombinant mouse granulocyte macrophagecolony stimulating factor,rmGM-CSF)、重组小鼠IL-4(recombinant mouse IL-4,rmIL-4,美国PeproTec公司);RIPA裂解液、PMSF、ECL发光液、β-Actin兔单克隆抗体(碧云天生物有限公司);脂多糖(lipopolysaccharide,LPS)、Alexa Fluor488标记的羊抗兔IgG(北京索莱宝公司);荧光单克隆抗体FITC-CD11c、PE-MHC Ⅱ、PE-CD86、PE-CD80(美国Bio Legend公司);OprF抗原特异性兔多克隆抗体(上海生工生物有限公司)。
Table 1 Aptamer sequences

RNA AptamerSequence5Cy3-min.25′Cy3-GGGAGGUGUGUUAGCACACGAUUCAUAAUCAGCUACCCUCCC-inverted dT 3′5S-min.25′SH-GGGAGGUGUGUUAGCACACGAUUCAUAAUCAGCUACCCUCCC-inverted dT 3′
1.2 仪 器
Malvern激光粒径测量仪、Malvern表面电位测量仪(美国Zetasizer公司);酶标仪(美国Thermo公司);荧光显微镜(日本Nikon公司);CytoFLEX流式细胞仪(美国贝克曼公司);UV凝胶成像仪(美国Bio-Rad公司)。
2 方 法
2.1 脂质体的制备
采用乙醇注入法制备阳离子脂质体,乙醇溶解物质的量比1∶1的胆固醇、DOTAP,55 ℃磁力搅拌下将脂质溶液滴加到生理盐水中搅拌1 h,冰浴下200 W超声10 min。
2.2 Lip-pOprF-VP22、Lip-pEGFP、Apt-Lip-pOprF-VP22的制备
静电吸附法制备负载pVAX1-OprF-VP22的阳离子脂质体(Lip-pOprF-VP22),按照不同的DOTAP与质粒质量比(DOTAP/pDNA分别为0∶0,2∶1,4∶1,5∶1,6∶1,8∶1,10∶1,pDNA质量保持不变,仅改变DOTAP质量),将pVAX1-OprF-VP22与阳离子脂质体涡旋混合后室温反应1 h,加入MPEG-2000-DSPE(DOTAP/MPEG-2000-DSPE,3∶1)60 ℃水浴15 min,冷却,即制得Lip-pOprF-VP22。同法制备负载pEGFP-N2的阳离子脂质体(Lip-pEGFP)。采用后插法[15]制备适配体5S-min.2修饰的Lip-pOprF-VP22(Apt-Lip-pOprF-VP22),即将上述反应中的MPEG-2000-DSPE替换为DSPE-PEG2000-MAL得到MAL基团修饰的Lip-pOprF-VP22(MAL-Lip-pOprF-VP22)后,加入适配体5S-min.2(5S-min.2预先在70 ℃变性3 min 后在室温下复性15 min),N2保护下20 ℃反应过夜。
2.3 琼脂糖凝胶阻滞分析
将Lip-pOprF-VP22进行1%琼脂糖凝胶电泳(120 V,30 min),UV凝胶成像仪中观察DNA迁移情况。
2.4 CCK-8法检测不同DOTAP/pDNA质量比Lip-pOprF-VP22的细胞毒性
使用EMEM完全培养基培养DC2.4细胞,于96孔板中,37 ℃、5%CO2培养24 h,以不加Lip-pOprF-VP22为空白对照,其余各组中加入不同DOTAP/pDNA质量比的Lip-pOprF-VP22,每组均设3个复孔,分别培养24 h、48 h后每孔加入CCK-8溶液10 μL,37 ℃继续孵育4 h,测定450 nm处的吸收度。
2.5 不同DOTAP/pDNA质量比的Lip-pEGFP体外转染效果
将DC2.4细胞接种于24孔板中,以PBS为空白对照,PEI为阳性对照(PEI与pDNA质量比为3∶1,后续实验均采用该比例),其余各组中加入不同DOTAP/pDNA质量比的Lip-pEGFP 100 μL,每组均设2个复孔。37 ℃、5%CO2培养,荧光显微镜观察24 h、48 h各组的荧光强度。
2.6 Lip-pOprF-VP22的粒径、电位及多分散性检测
将适量的DOTAP/pDNA质量比为5∶1的复合物稀释在无菌去离子水1 mL中,测定其粒径及电位。
2.7 适配体的靶向性验证
以CHO细胞为DEC-205阴性表达细胞,DC2.4细胞为DEC-205阳性表达细胞。细胞接种于6孔板,37 ℃、5%CO2培养6 h至细胞贴壁,各组分别加入5Cy3-min.2(适配体预先在70 ℃变性3 min,室温下复性15 min)5 μL,每组均设2个复孔。37 ℃、5%CO2孵育0.5、2、4 h,PBS润洗孔板3次,荧光显微镜下观察荧光强度。
2.8 免疫荧光法检测OprF-VP22抗原蛋白的表达
将DC2.4细胞接种于24孔板中培养24 h,以PBS为空白对照,PEI为阳性对照,实验组分别加入 DOTAP/pDNA质量比为5∶1的Lip-pOprF-VP22、Apt-Lip-pOprF-VP22 100 μL,每组均设2个复孔,37 ℃、5%CO2培养。4组细胞分别于转染后48 h进行免疫荧光染色,具体步骤为:吸去培养基,4%多聚甲醛固定20 min,PBS冲洗3次,5% BSA 37 ℃封闭30 min,用OprF抗原特异性抗体兔多克隆抗体(1∶1 000)4 ℃孵育过夜,次日用PBS冲洗3次,Alexa Fluor488标记的羊抗兔IgG抗体 37 ℃避光孵育1 h,PBS冲洗3次,DAPI染核8 min,PBS冲洗3次,荧光显微镜下观察荧光强度。
2.9 Western blot检测OprF-VP22抗原蛋白的表达
将DC2.4细胞接种于6孔板中培养24 h,按方法“2.8”项分组。4组细胞分别于转染后48 h用RIPA裂解液裂解,收集各组细胞裂解液,加入上样缓冲液煮沸10 min,采用BCA法测定蛋白浓度,常规方法进行Western blot实验,最后使用HRP化学发光试剂(ECL)将清洗好的PVDF膜显影,检测蛋白条带。
2.10 体外Apt-Lip-pOprF-VP22对小鼠BMDCs成熟的影响
2.10.1 BMDCs的诱导培养及鉴定 颈椎脱臼处死6~8周龄的C57BL雄性小鼠,按文献所述方法收集BMDCs[16],37 ℃、5% CO2培养7 d后收集到的疏松贴壁细胞即为未成熟小鼠骨髓来源树突状细胞(bone marrow-derived dendritic cells,BMDCs)。含2% FBS的PBS洗涤并重悬细胞,调整细胞浓度为每毫升1.0×106,加入FITC-CD11c 0.5 μL,4 ℃避光孵育30 min后用含2% FBS的PBS洗涤 3次,预冷PBS重悬,进行流式细胞仪检测。
2.10.2 Apt-Lip-pOprF-VP22对BMDCs成熟的影响 未成熟BMDCs(每毫升1.0×106个)接种于6孔板中,每孔加入含添加了20 ng/mL rmGM-CSF和20 ng/mL rmIL-4的RPMI 1640 完全培养基2 mL。分别加入Apt-lip(不载pDNA)、Lip-pOprF-VP22、Apt-Lip-pOprF-VP22到培养孔,另设置PBS空白对照与LPS(1 μg/mL)阳性对照,每组均设3个复孔,将各组细胞放入37 ℃、5% CO2培养48 h。将每孔细胞分8组:阴性对照、FITC-CD11c、PE-CD80、PE-CD86、PE-MHC Ⅱ、FITC-CD11c+ PE-CD80、FITC-CD11c+PE-CD86、FITC-CD11c+PE-MHC Ⅱ。按前述方法标记抗体,并送流式细胞仪检测。
2.11 统计学处理
3 结 果
3.1 琼脂糖凝胶阻滞分析
采用琼脂糖凝胶阻滞实验观察不同DOTAP/pDNA质量比的Lip-pOprF-VP22对pVAX1-OprF-VP22的包封效果。图1可见,随着DOTAP/pDNA质量比的增加,脂质体抑制DNA流动的性能增强,即脂质体对pVAX1-OprF-VP22的包封效果增强,当DOTAP/pDNA质量比为5∶1时即可完全抑制pVAX1-OprF-VP22的流动,达到最佳的包封效果。

Figure 1 Encapsulation effect of Lip-pOprF-VP22 with different mass ratios of DOTAP/pDNA on pVAX1-OprF-VP22
3.2 不同DOTAP/pDNA质量比Lip-pOprF-VP22的细胞毒性
采用CCK-8法检测不同DOTAP/pDNA质量比的Lip-pOprF-VP22对DC2.4的细胞毒性。图2可见,随着DOTAP/pDNA质量比的增加,不同复合物处理24、48 h后,各复合物对细胞存活率几乎无影响(P>0.05),细胞存活率均在80%以上,说明该脂质体复合物细胞毒性低,是一种生物相容性较好的基因递送载体。
3.3 不同DOTAP/pDNA质量比的Lip-pEGFP体外转染效果
使用荧光显微镜观察不同DOTAP/pDNA质量比Lip-pEGFP对DC2.4细胞的转染效果。图3结果表明,当DOTAP/pDNA质量比从2∶1增加至 10∶1,Lip-pEGFP对DC2.4细胞的转染率先增加后降低,当DOTAP/pDNA质量比为4∶1、5∶1时,Lip-pEGFP 的荧光强度较高,说明转染率相对较高;而转染48 h较转染24 h的荧光更强,转染效率更好。
3.4 Lip-pOprF-VP22的粒径、电位及多分散性检测
上述实验结果表明DOTAP/pDNA质量比为5∶1时,Lip-pOprF-VP22对pVAX1-OprF-VP22的包封效果最佳且细胞毒性低、转染率较高。因此对该比例的DOTAP/pDNA复合物进行表征,载或不载pVAX1-OprF-VP22阳离子脂质体的平均粒径、Zeta电位及PDI测定结果见表2和图4,符合基因递送条件[17]。
3.5 适配体的靶向性验证
为了验证实验所选的适配体对DC具有特异的靶向性,采用荧光显微镜观察min.2与DC2.4的结合能力。图5表明,当5Cy3-min.2适配体与不同细胞孵育2 或4 h时,DC2.4组能观察到更多的红色荧光,而CHO组几乎不能观察到红色荧光,说明RNA适配体min.2具有较好的DC靶向性。



Figure 3 Transfection effects of Lip-pEGFP with different mass ratios of DOTAP/pDNA to DC2.4 for 24 hours (A) and for 48 hours (B)
Table 2 Particle size,PDI and Zeta potential of different liposome complexes (the mass ratio of DOTAP/pDNA is 5∶1)

GroupSize/nmPDIZeta/mVLiposome184.4±5.300.36161.00±1.68Lip-pOprF-VP22171.7±1.270.11811.30±0.570

Figure 4 Particle size (A) and Zeta potential (B) distribution of Lip-pOprF-VP22 when the mass ratio of DOTAP/pDNA was 5∶1

Figure 5 Verification the targeting properties of min.2 (×100)
3.6 OprF-VP22抗原蛋白的表达
采用免疫荧光及Western blot检测不同复合物转染DC2.4后OprF-VP22蛋白的表达。图6-A表明,与PBS组相比,Lip-pOprF-VP22组和Apt-Lip-pOprF-VP22组均有较强的绿色荧光,但Apt-Lip-pOprF-VP22组荧光强度更大。这表明通过适配体对DC的靶向作用,Apt-Lip-pOprF-VP22组中更多OprF-VP22基因进入胞内并成功表达OprF-VP22蛋白。同样,Western blot实验也进一步印证了免疫荧光结果(图6-B)。

Figure 6 Immunofluorescence assay (×200) (A) and Western blot assay (B) for expression of OprF-VP22 antigen protein
3.7 体外Apt-Lip-pOprF-VP22对小鼠BMDCs成熟的影响
3.7.1 BMDCs的诱导培养及鉴定 CD11c是小鼠BMDCs的特征性表面分子,通过观察细胞形态及检测CD11c的表达量,可以反映出骨髓中诱导的BMDCs的数量。由图7-A、7-B可知,未成熟的树突状细胞(immature DCs,imDCs)聚集成簇,贴壁不牢;成熟的树突状细胞(mature DCs,mDCs)悬浮生长,单细胞成毛刺状、出现典型的树突样突起。图7-C、7-D流式细胞仪分析出的表达CD11c的细胞数量约占70%,即表明通过骨髓造血干细胞分化出的DC数量约占70%。
Figure 7 photomicrograph of immature dendritic cells (imDCs) (×200) (A),photomicrograph of mature dendrtitic cells (mDCs) (×200) (B),collecting the living cell population by flow cytometry method (C) and number of cells expressing CD11c in the imDCs cell population (D)
3.7.2 Apt-Lip-pOprF-VP22对BMDCs成熟的影响 成熟的DC表面表达大量的MHC Ⅱ类分子和共刺激分子如CD80、CD86,而未成熟DC只能少量表达这些分子。MHC Ⅱ类分子、CD80和CD86是DC识别和摄取抗原必要的信号转导分子,也是评价DC免疫功能的最佳指标。实验中采用流式细胞术检测上述分子在BMDCs的表达(图8-A)。图8-B表明,与PBS空白对照组相比,Lip-pOprF-VP22组及Apt-Lip组均不能增加CD80分子的表达;与Lip-pOprF-VP22组相比,Apt-Lip-pOprF-VP22组及LPS阳性对照组均能显著增加CD80分子的表达,且Apt-Lip-pOprF-VP22及LPS两组间效果无差异。图8-C表明,与PBS组相比,Apt-Lip组不能增强CD86的表达,Lip-pOprF-VP22、Apt-Lip-pOprF-VP22及LPS各组均能增强CD86的表达;虽然LPS组与Lip-pOprF-VP22组或Apt-Lip-pOprF-VP22组相比,不能增强CD86表达,但Apt-Lip-pOprF-VP22组较Lip-pOprF-VP22组能显著增强CD86的表达。图8-D表明,与PBS组相比,Apt-Lip组不能增强MHC Ⅱ的表达,而Lip-pOprF-VP22组、Apt-Lip-pOprF-VP22组及LPS组均能增强MHC Ⅱ的表达;而与Lip-pOprF-VP22组相比,Apt-Lip-pOprF-VP22组及LPS阳性对照组均能显著增加BMDCs表面MHC Ⅱ的表达,且Apt-Lip-pOprF-VP22及LPS两组间效果无差异。上述结果说明Apt-Lip-pOprF-VP22对BMDCs的成熟效果比Lip-pOprF-VP22更显著,其与阳性对照LPS具有相似的显著促进作用。
4 讨 论
PA是临床上最为常见的病原体之一,研发PA疫苗是预防其感染的重要措施。课题组前期利用FDA批准的pVAX1载体构建了pVAX1-OprF-VP22融合DNA疫苗,研究证实:与OprF DNA疫苗相比,OprF/VP22融合DNA疫苗显著增强体液、细胞免疫及免疫保护作用[4]。尽管如此,DNA疫苗在体内易被核酸酶降解且外周DC数量少导致其特异性摄取量很低,使得DNA疫苗在体内免疫原性低。因此,目前尚无DNA疫苗应用于临床。
阳离子脂质体为最常用的基因递送载体,其可通过静电作用与细胞膜融合而携带DNA分子进入胞内,体内外转染效果良好。研究表明,使用阳离子脂质体递送DNA疫苗还可促进APC对其的摄取,激活TLR并产生IFN,刺激适应性免疫应答[17]。本实验中,采用操作简单的乙醇注入法制备阳离子脂质体,从而避免了有机溶剂的使用,然后通过简单的静电吸附法成功制备Lip-pOprF-VP22。有研究者指出胶体脂质微粒携带的电荷越多、空间位阻越大,稳定性越好[18],因此,为了增加Lip-pOprF-VP22的动力学稳定性,本研究选用亲水性的长链PEG修饰脂质体,增加Lip-pOprF-VP22的空间位阻,阻止脂质体的融合与聚集[19];另外,通过协调阳离子脂质与MPEG-2000-DSPE的比例,使得制备的脂质体带上合适的正电荷,依靠同种电荷相互排斥作用增强Lip-pOprF-VP22的动力学稳定性。

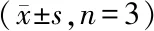
*P<0.05vsPBS group;#P<0.05vsLip-pOprF-VP22 group
本实验中制备的Lip-pOprF-VP22随着DOTAP/pDNA质量比增加,包封效果增强,当质量比为5∶1时,达到最佳的包封效果(图1):这说明该比例下形成的复合物具有很强的静电相互作用,因此阳离子脂质体便可以更好地保护DNA疫苗,使得DNA疫苗在酶解前被APC内吞。此外,在实验中发现,随着DOTAP/pDNA质量比增加,Lip-pEGFP对DC的转染率先增加后降低(图3),这可能是由于Lip-pEGFP表面电荷的改变使得转染率发生变化:即MPEG-2000-DSPE的加入使得载pDNA的Lip的表面电荷被屏蔽,随着DOTAP/pDNA质量比增加,MPEG-2000-DSPE的量也成比例增加,当MPEG-2000-DSPE增大到一定量时,足以屏蔽掉增加的DOTAP所携带的正电荷,使得表面电荷又出现降低的趋势。在DOTAP/pDNA质量比为4∶1、5∶1时,制备的脂质体复合物转染率高,具有作为基因载体的优良潜质。除具有高的转染率外,理想的基因递送载体还应具有良好的生物安全性[20]。其中,载体的细胞毒性是一个最重要的生物安全性指标,CCK-8实验结果表明(图2),本研究制备的Lip-pOprF-VP22几乎是安全无毒的。
通过对包封效果、转染率、细胞毒性的评估,选取最佳DOTAP/pDNA质量比为5∶1的Lip-pOprF-VP22进行表征并用于连接适配体。粒径在100~200 nm带正电荷的微粒应用于基因递送是最合适的[21],在DOTAP/pDNA质量比为5∶1时,制备的Lip-pOprF-VP22粒径约170 nm,Zeta电位约12 mV(图4),符合基因递送条件。进一步通过后插法在Lip-pOprF-VP22表面修饰以DEC-205特异性的RNA适配体min.2,构建了DC靶向 PA DNA疫苗递送系统。后插法可避免一步法(即将靶向配体修饰的PEG磷脂与脂质体磷脂和胆固醇共同旋转成膜,按照薄膜分散的方法制备靶向脂质体)制备Apt-Lip-pOprF-VP22存在的问题:成膜不均;靶头可存在于脂质体内外两侧造成靶头的浪费。此外,后插法还可避免成膜中所用有机溶剂对配体活性的影响[22]。故而选用后插法连接适配体的方案显得更为优越。
在基因治疗中有效的细胞内吞和靶向能力是获得有效治疗结果的必要条件[23],而在DNA疫苗中,抗原特异性的被APC捕获并且内吞进入细胞是将抗原递呈给细胞的关键步骤。本研究首先证实了适配体min.2能与DC特异性结合(图5),具有特异性的DC靶向性能;随后,通过免疫荧光实验验证了DOTAP/pDNA质量比为5∶1时Lip-pOprF-VP22及Apt-Lip-pOprF-VP22转染DC后OprF蛋白的表达,结果也表明Apt-Lip-pOprF-VP22有一定的DC靶向性,其能靶向DC并表达出更多OprF蛋白(图6);接着又探讨了Lip-pOprF-VP22和min.2修饰的递送系统Apt-Lip-pOprF-VP22对BMDCs成熟的影响,旨在通过对BMDCs表面成熟相关分子的表达差异来证实Apt-Lip-pOprF-VP22靶向DC后能表达出的抗原蛋白并刺激DC成熟。和预期结果一致,Lip-pOprF-VP22和Apt-Lip-pOprF-VP22均能促进DC成熟,但Apt-Lip-pOprF-VP22与LPS阳性对照相似,可更加显著的促进DC成熟(图8)。究其原因,DC对外来物质的内吞主要受特异的细胞膜受体和胞内信号通路的调控[24],min.2可通过与DC表面DEC-205受体结合后,在受体介导的内吞下进入胞内,抗原基因表达出相应的抗原蛋白促进DC成熟并活化初始T淋巴细胞,进而引发体液及细胞免疫。因此,制备的Apt-Lip-pOprF-VP22在PA DNA疫苗的研究中可能具有较好的应用潜力。
本实验通过后插法成功制备了安全性好、DC靶向性好、促进DC成熟的PA DNA疫苗递送载体,将进一步优化适配体的连接比例以增强递送载体的DC靶向性,并研究该适配体修饰的脂质体DNA复合物的体内靶向DC能力,通过对体内免疫效力及免疫小鼠的PA感染率的评价来观察该递送载体能否增强PA DNA疫苗的体内免疫原性,为PA DNA疫苗的研发提供新的思路。
猜你喜欢
昆明医科大学学报(2022年3期)2022-04-19
中国化妆品(2022年2期)2022-03-04
昆明医科大学学报(2021年3期)2021-07-22
中国药学药品知识仓库(2021年18期)2021-02-28
昆明医科大学学报(2021年1期)2021-02-07
信息技术时代·上旬刊(2019年4期)2019-09-10
数码世界(2018年1期)2018-12-23
生物学教学(2018年10期)2018-11-29
商情(2017年30期)2017-09-18
江苏农业科学(2016年1期)2017-05-17
