
乌头属植物ITS 条形码的序列及结构特征研究
2019-05-17 05:40张丹丹吴智华张婧捷
西南民族大学学报(自然科学版) 2019年2期
覃 瑞,张丹丹,刘 虹,吴智华,张婧捷,韩 迪,王 靖
(1.中南民族大学武陵山区特色资源植物种质保护与利用湖北重点实验室,湖北 武汉 430074;2 农业部食物与营养发展研究所,北京 100081)
毛茛科( Ranunculaceae)乌头属(Aconitum L.)在我国记载分布200 多种[1],分为露蕊乌头亚属(Subgen. Gymnaconitum )、乌头亚属(Subgen. Aconitum)、牛扁亚属(Subgen. Paraconitum),3 个亚属中约170种植物主要分布于青藏高原及其临近地区;除海南岛外,其他各省区亦均有分布[2-3].乌头属植物含有乌头碱等生物碱,在民族地区广泛用于治疗跌打损伤、风湿骨痛以及流感等[4]. 在《本草纲目》、《中华草本》、《伤寒论》中乌头类药物及其使用方法均有记载[5].中国药典1977 版收录了雪上一枝蒿(短柄乌头的块根、叶、花蕾、花、幼苗),中国药典2015 版收载了草乌(北乌头的干燥块根),川乌(乌头的干燥母根),附子(乌头的干燥子根)[6]. 目前报道的药用成分主要是其中含有的二萜生物碱,具有镇痛、消炎、局麻等效果.近年来从高乌头根中提取的拉巴乌头碱(lappaconine)制成的氢溴酸盐[7],由于其良好的镇痛不成瘾等特性使高乌头药材具有广泛的应用前景.然而由于乌头属植物形态上具有高度相似性,在使用中同名异物或同物异名的现象普遍存在[8],比如紫乌头在我国民间就常做草乌入药,伏毛铁棒锤的块根在藏族有作“雪上一枝蒿”药用[9]. 仅从形态学上鉴定乌头属植物容易导致药材的混用[10-11],因此根据分子特征来鉴定乌头属植物更为准确,可为道地药材的使用与推广提供分子学鉴定依据.
DNA 条形码是利用若干DNA 片段作为标准片段,对物种进行快速准确的自动化标记,将DNA 序列与生物物种相对应[12].2009 年国际生命条形码联盟植物工作组建议将rbcL + matK 组合作为陆地植物的核心DNA 条形码[13];2011 年中国条形码研究团队建议将ITS(Internal Transcribed Space) 或ITS2(Internal Transcribed Space 2)作为陆地植物核心条形码的补充条形码[14].ITS 是指18S rDNA 和28S rDNA 基因的间隔序列,由ITS1 区、5.8S 区、ITS2 区组成,这段序列进化速度较快,且近缘种的进化速度比较一致,不仅广泛用于近缘属、种间、属内的分子系统学及鉴别,而且也广泛用于种内居群间的差异性研究[15].采用ITS 序列来分析不同乌头属植物,为乌头属道地药材分类和使用提供分子学证据,为后续青藏地区相关研究提供基础.
另外根据ITS 的二级结构与补偿性碱基配对变化(CBCs,compensatory base pair changes)位点来探究乌头属种间杂交情况.目前在与ITS 二级结构相关的研究中,CBCs 开始越来越多的应用到种间和种内关系的研究中[16].Muller 等人用大量的数据对CBC criterion 假设进行了验证,对30 万个ITS2 二级结构综合分析表明,两个个体之间存在CBC,它们是两个不同的种的概率约为93%;两个个体之间不存在CBC,它们是同一种的概率约为76%[17].因此,以此来分析不同乌头属植物之间的亲缘关系与遗传距离,对青藏地区种质资源的研究具有重要意义.
1 材料与方法
1.1 实验材料
本研究材料以甘肃遵义地区乌头属植物为主,在湖北、陕西、四川、河南、甘肃采集形态相似的乌头属植物,并与新疆地区形态相似的乌头属植物相比较.收集9 种乌头属材料,共11 个样本.经实验室专家依形态学鉴定,ZY(遵义)乌头为高乌头或白喉乌头或花葶乌头,在当地又称贵州穿心莲、麻布袋、毛茛等,神农架、太白山、青城山、宣恩、利川、老界岭的为高乌头,甘肃白水江为赣皖乌头,新疆为空茎乌头. 另外,从GenBank 下载了已测序的国内乌头属植物共75 条序列.详细材料信息见表1.
1.2 材料与方法
1.2.1 DNA 提取9 种乌头属植物采集于不同产地,对其硅胶干燥的叶片,采用改良的CTAB 法提取总DNA.
1.2.2 扩增与测序
使用引物ITS5、ITS4[17],扩增全长ITS(包括5.8S) 区 域. 引 物 序 列: ITS4: 5-TCCTCCGCTTATTGATATGC-3; ITS5: 5-GGAAGTAAAAGTCGTAACAAGG-3 .PCR 扩增程序:70 ℃4 min;94 ℃1 min,52 ℃20 s,72 ℃50 s;94 ℃20 s,52 ℃20 s,72 ℃50 s(36 cycle);72 ℃4 min;4 ℃∞.PCR 反应体系为50 μl:ITS4 与ITS5 各2.5 μl(10 μm/L),扩增Mix 25 μl,DNA 2.5 μl(10 ng/μl),H2O 17. 5 μl.1%琼脂糖凝胶电泳检测,试剂盒切胶回收,送至武汉擎科伟业生物科技有限公司进行ABI3730 测序.
2 数据处理
实验所得 DNA 序列在 NCBI 数据库进行BLASTN 比对,依相似性最高得分排序,与乌头属植物相似分值达到99 以上,从GenBank 下载国内乌头属植物的序列.将实验所得序列与GenBank 下载序列一起在MEGA X 进行比对(Alignment),去除两端的序列,得到可分析的ITS 总长,基于K2P(Kimura 2-parameter)模型计算9 种乌头属植物的遗传距离,用邻接法(Neighbor-Joining)构建距离树,拓扑结构的可靠性用1000 次重复的自展检验(Bootstrap analysis)来评估,分支支持率低于50% 的合并. 从GenBank 下载ITS1 与ITS2 序列,利用MEGA X 将下载的ITS1 和ITS2 序列与实验所得ITS 序列比对,得到可分析的ITS1 和ITS2 序列,运用Freiburg RNA Tools[18]预测ITS1 以及ITS2 一致二级结构,ITS1 序列利用mfold web serve[19]预测二级结构,参数设置为默认的37℃(http:/ /unafold.rna. albany. edu/);ITS2 序列在ITS2 Database (http: / /its2. Bioapps. Biozentrum. uniwuerzburg.de/)上预测二级结构,将DNA 序列与二级结构导入4SALE[20-21]统计软件中,比对ITS2 二级结构和序列,统计CBCs[22]位点.
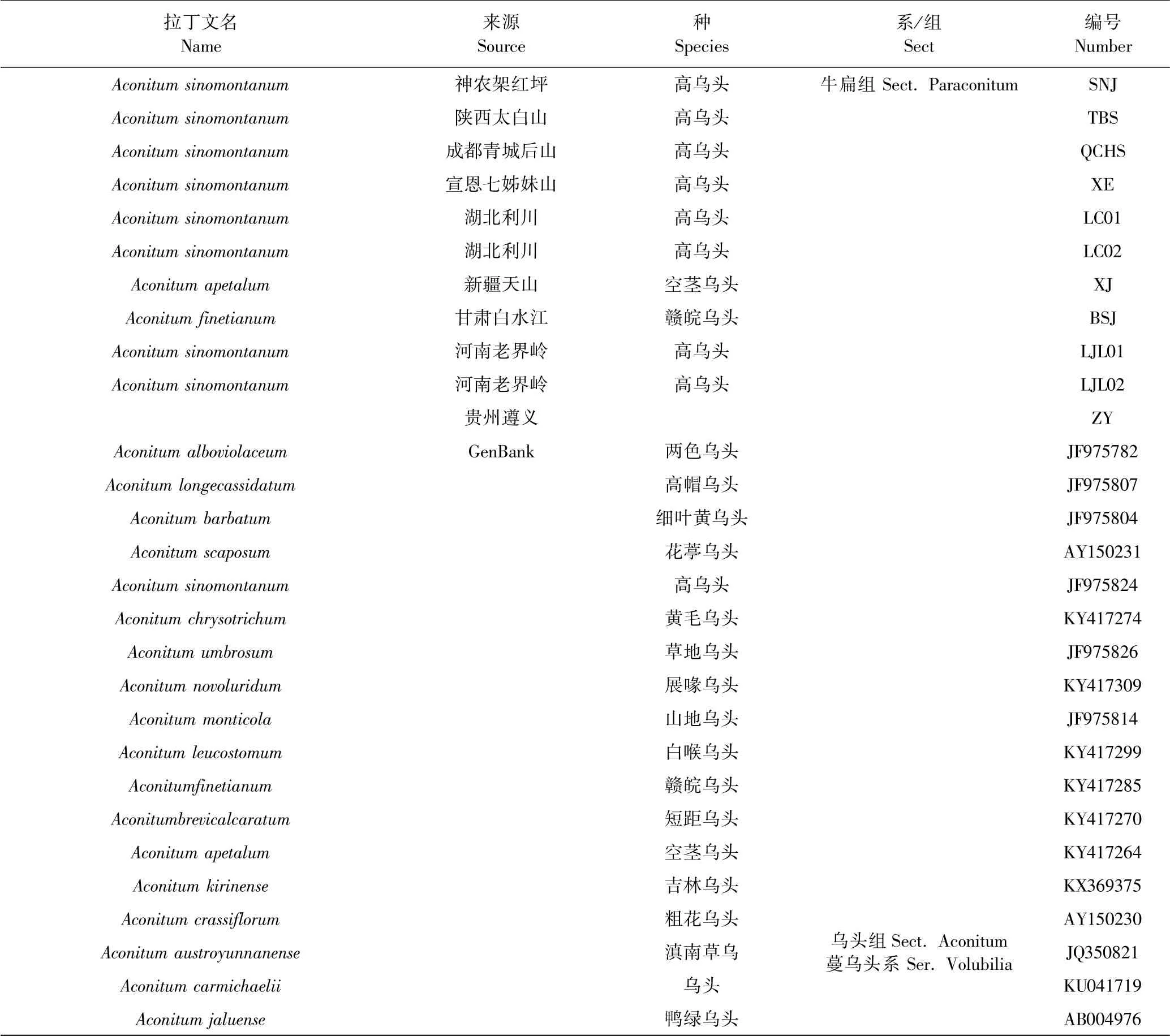
表1 材料来源Table 1 Material source
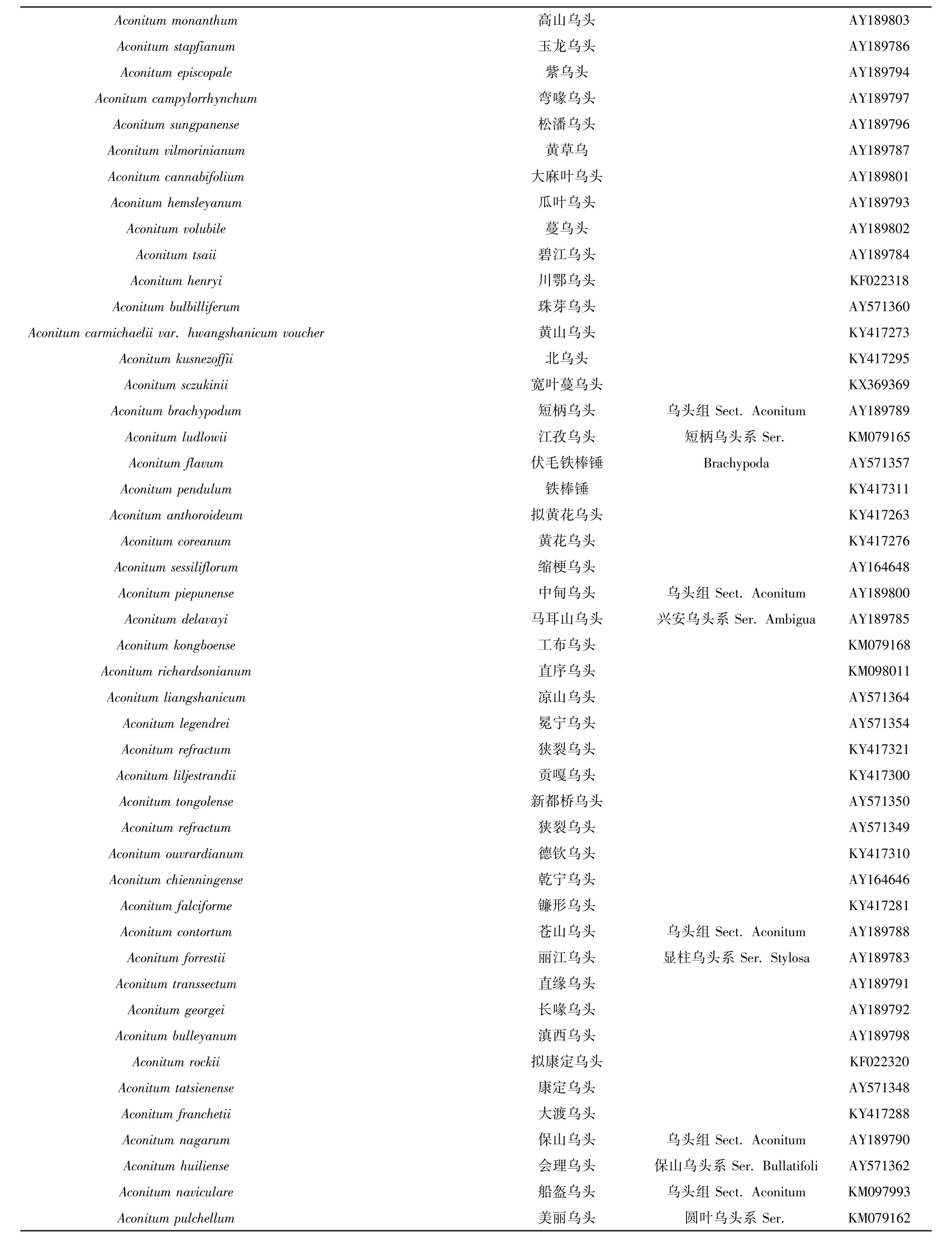
Aconitum monanthum 高山乌头 AY189803 Aconitum stapfianum 玉龙乌头 AY189786 Aconitum episcopale 紫乌头 AY189794 Aconitum campylorrhynchum 弯喙乌头 AY189797 Aconitum sungpanense 松潘乌头 AY189796 Aconitum vilmorinianum 黄草乌 AY189787 Aconitum cannabifolium 大麻叶乌头 AY189801 Aconitum hemsleyanum 瓜叶乌头 AY189793 Aconitum volubile 蔓乌头 AY189802 Aconitum tsaii 碧江乌头 AY189784 Aconitum henryi 川鄂乌头 KF022318 Aconitum bulbilliferum 珠芽乌头 AY571360 Aconitum carmichaelii var. hwangshanicum voucher 黄山乌头 KY417273 Aconitum kusnezoffii 北乌头 KY417295 Aconitum sczukinii 宽叶蔓乌头 KX369369 Aconitum brachypodum 短柄乌头 乌头组Sect. Aconitum AY189789 Aconitum ludlowii 江孜乌头 短柄乌头系Ser. KM079165 Aconitum flavum 伏毛铁棒锤 Brachypoda AY571357 Aconitum pendulum 铁棒锤 KY417311 Aconitum anthoroideum 拟黄花乌头 KY417263 Aconitum coreanum 黄花乌头 KY417276 Aconitum sessiliflorum 缩梗乌头 AY164648 Aconitum piepunense 中甸乌头 乌头组Sect. Aconitum AY189800 Aconitum delavayi 马耳山乌头 兴安乌头系Ser. Ambigua AY189785 Aconitum kongboense 工布乌头 KM079168 Aconitum richardsonianum 直序乌头 KM098011 Aconitum liangshanicum 凉山乌头 AY571364 Aconitum legendrei 冕宁乌头 AY571354 Aconitum refractum 狭裂乌头 KY417321 Aconitum liljestrandii 贡嘎乌头 KY417300 Aconitum tongolense 新都桥乌头 AY571350 Aconitum refractum 狭裂乌头 AY571349 Aconitum ouvrardianum 德钦乌头 KY417310 Aconitum chienningense 乾宁乌头 AY164646 Aconitum falciforme 镰形乌头 KY417281 Aconitum contortum 苍山乌头 乌头组Sect. Aconitum AY189788 Aconitum forrestii 丽江乌头 显柱乌头系Ser. Stylosa AY189783 Aconitum transsectum 直缘乌头 AY189791 Aconitum georgei 长喙乌头 AY189792 Aconitum bulleyanum 滇西乌头 AY189798 Aconitum rockii 拟康定乌头 KF022320 Aconitum tatsienense 康定乌头 AY571348 Aconitum franchetii 大渡乌头 KY417288 Aconitum nagarum 保山乌头 乌头组Sect. Aconitum AY189790 Aconitum huiliense 会理乌头 保山乌头系Ser. Bullatifoli AY571362 Aconitum naviculare 船盔乌头 乌头组Sect. Aconitum KM097993 Aconitum pulchellum 美丽乌头 圆叶乌头系Ser. KM079162
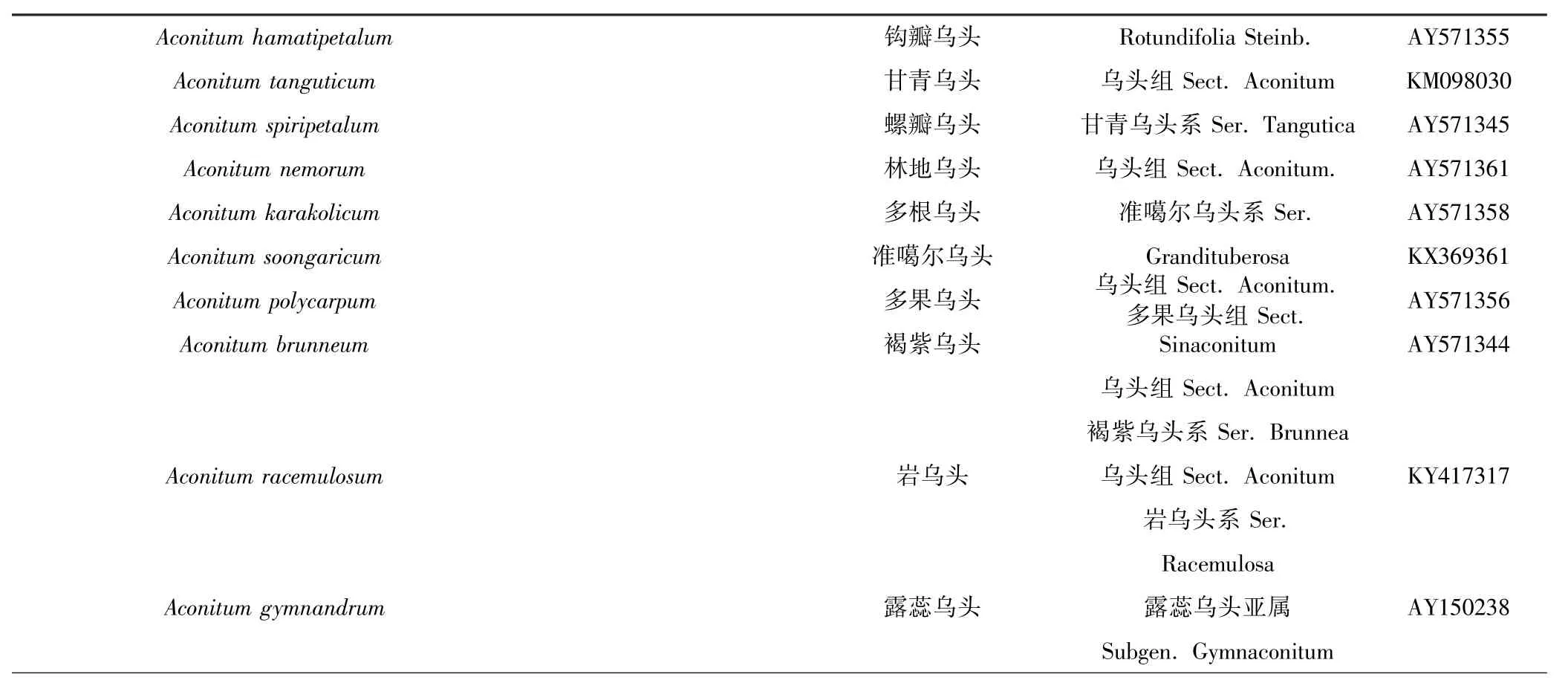
Aconitum hamatipetalum 钩瓣乌头 Rotundifolia Steinb. AY571355 Aconitum tanguticum 甘青乌头 乌头组Sect. Aconitum KM098030 Aconitum spiripetalum 螺瓣乌头 甘青乌头系Ser. Tangutica AY571345 Aconitum nemorum 林地乌头 乌头组Sect. Aconitum. AY571361 Aconitum karakolicum 多根乌头 准噶尔乌头系Ser. AY571358 Aconitum soongaricum 准噶尔乌头 Grandituberosa KX369361 Aconitum polycarpum 多果乌头 乌头组Sect. Aconitum.多果乌头组Sect. AY571356 Aconitum brunneum 褐紫乌头 Sinaconitum AY571344乌头组Sect. Aconitum褐紫乌头系Ser. Brunnea Aconitum racemulosum 岩乌头 乌头组Sect. Aconitum KY417317岩乌头系Ser.Racemulosa Aconitum gymnandrum 露蕊乌头 露蕊乌头亚属 AY150238 Subgen. Gymnaconitum
3 结果
3.1 种内种间遗传变异分析
实验所得rDNA 序列与从GenBank 下载序列,以露蕊乌头(Aconitum gymnandrum) 为外类群,利用MEGA X 进行比对,得到可分析的ITS1(表2)长度在244 ~250bp,平均GC 含量60.5%,检测到保守位点(conserved sites)158 个、变异位点(variable sites)89个、简约信息位点(parsimony-informative sites)31 个.ITS2 可分析全长217 ~220bp,平均GC 含量61.2%,检测到保守位点(conserved sites)167 个、变异位点(variable sites)53 个、简约信息位点(parsimony-informative sites)20 个.10 个种ITS1 的K2P(Kimura 2-parameter)遗传距离表(表3)显示分布于0 至0.331之间,平均遗传距离为0.110.其中XE(宣恩)与LC01(利川)遗传距离为0,露蕊乌头与其余几种遗传距离均大于0.2,其次是ZY 与各种的遗传距离偏大,表现出明显的差异,露蕊乌头为露蕊乌头亚属,ZY 为乌头亚属,其余均为牛扁亚属且遗传距离偏小.ITS2 遗传距离表(表4)显示种间与种内水平上,ITS2 的遗传距离小于ITS1,在这几种乌头属植物中,ITS1 的变异程度大于ITS2.
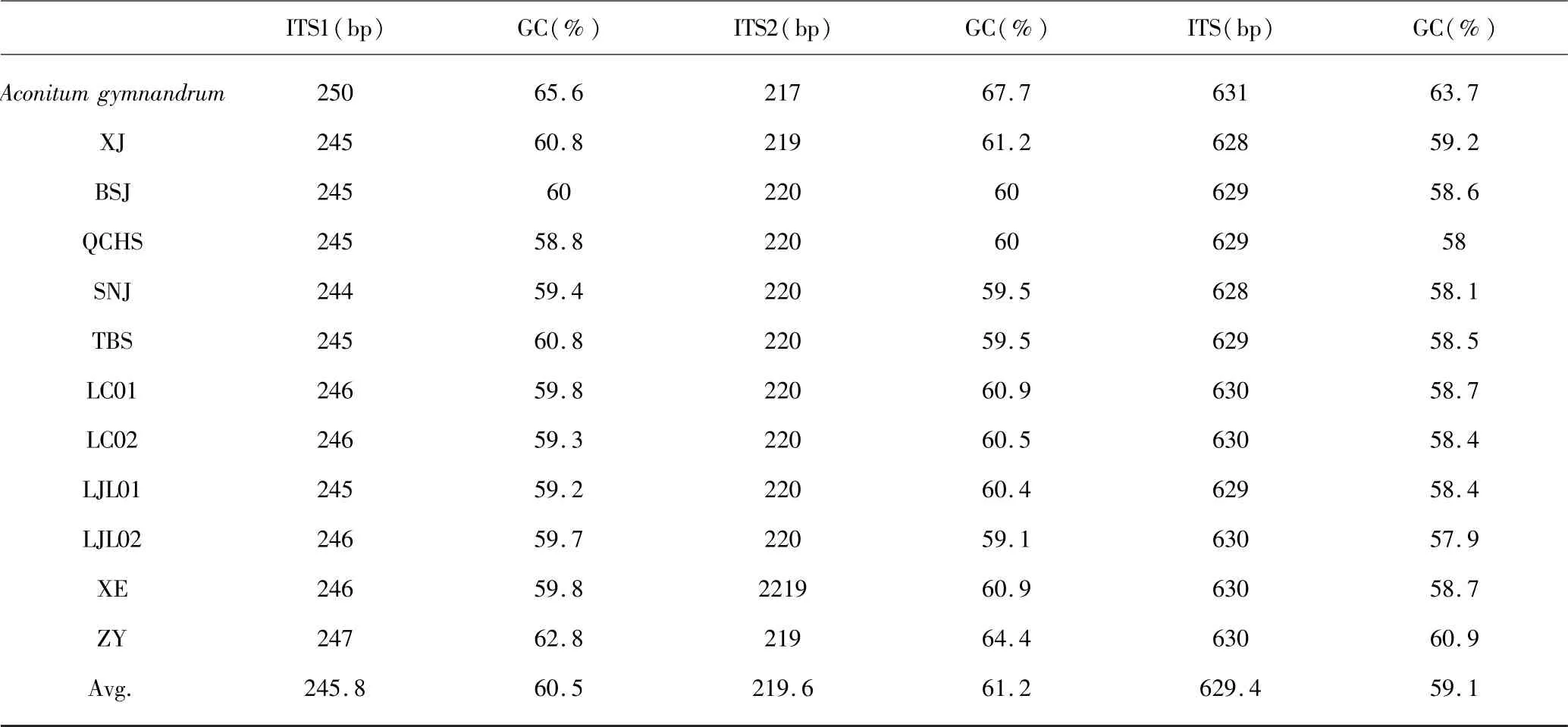
表2 ITS 的长度及GC 含量统计Table 2 ITS length and GC content statistics
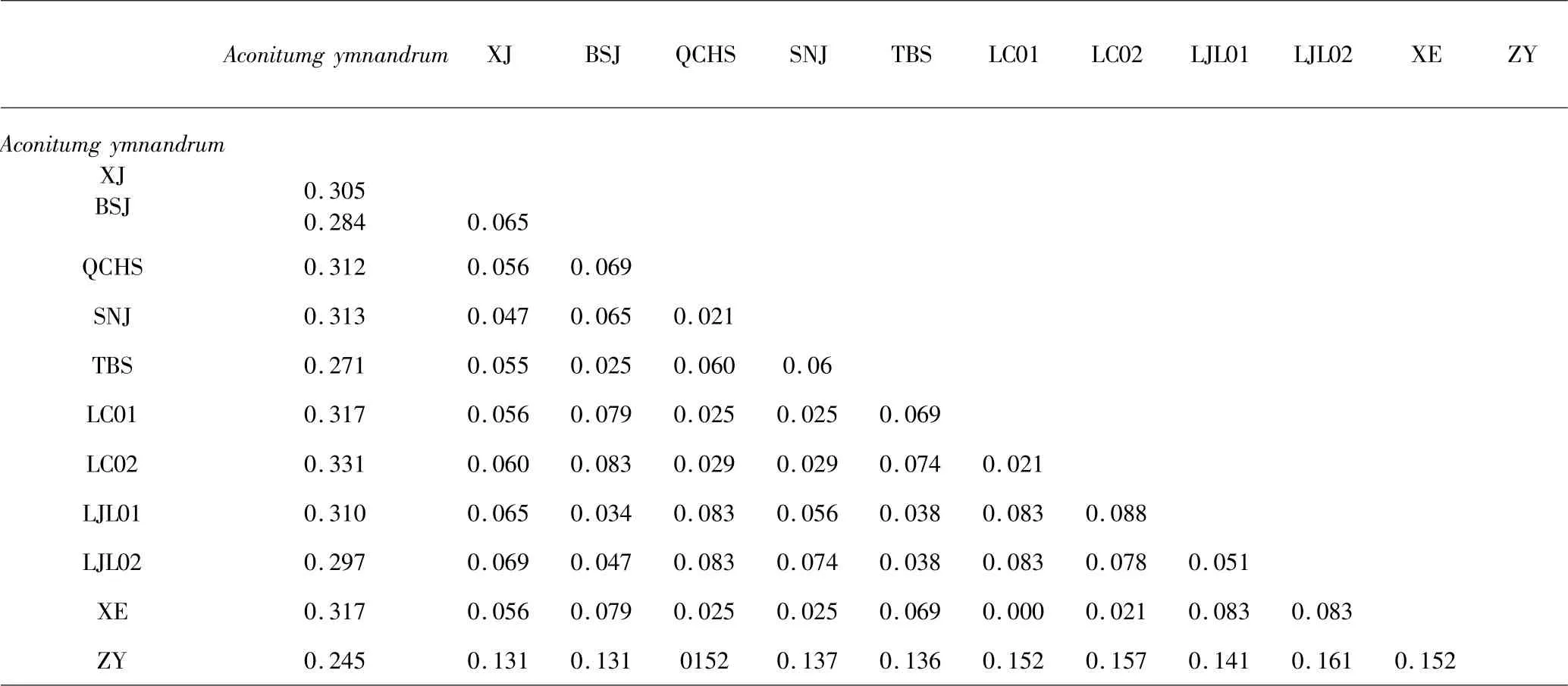
表3 乌头属植物ITS1 的遗传距离Table 3 Genetic distance of ITS1
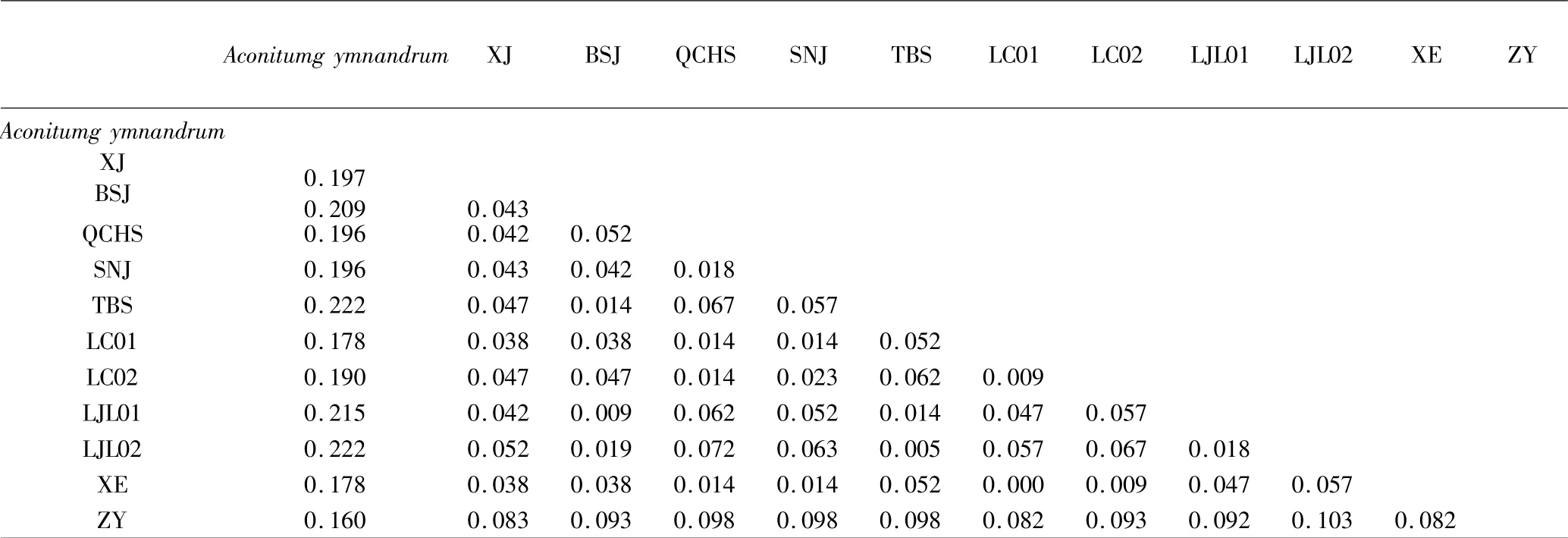
表4 乌头属植物ITS2 的遗传距离Table 4 Genetic distance of ITS2
3.2 ITS 一级结构聚类分析
将实验所得序列与GenBank 下载序列一起在MEGA X 进行比对(Alignment),去除两端的序列,基于K2P(Kimura 2-parameter)用邻接法(Neighbor-Joining)构建距离树(图1). 结果显示,在与GenBank 序列聚类的邻接树中,以露蕊乌头(A. gymnandrum)为外类群,在下属分支中,牛扁亚属与乌头亚属能完全区分开,ZY 与其他8 种乌头属植物属于不同亚属,为乌头亚属植物,其他8 种为牛扁亚属植物.ZY(遵义)与北乌头(A. kusnezoffii)聚为一支,XJ(新疆)与空茎乌头(A. apetalum)bootstrap value 为99,花葶乌头(A.scaposum)与LC01(利川)、LC02(利川)、QCHS(青城后山)、XE(宣恩)、SNJ(神农架)的bootstrap value 为80,TBS(太白山)、LJL02(利川)与高乌头(A. sinomontanum)聚为一支,BSJ(白水江)乌头与赣皖乌头(A. finetianum)的bootstrap value 为83. 亚属水平上的发育树分类与形态学分类一致,XJ(新疆)、BSJ(白水江)、TBS(太白山)与LJL(老界岭)种水平的聚类与形态学一致.
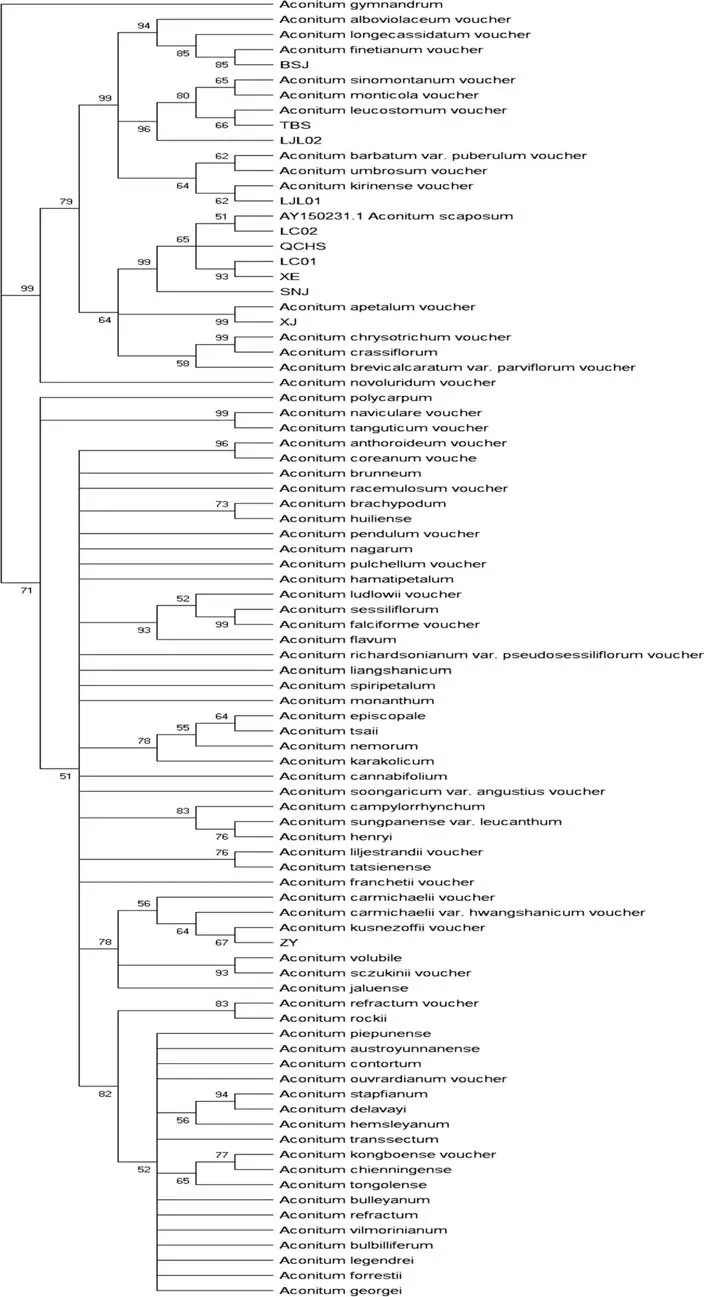
图1 基于ITS 序列构建邻接树,Bootstrap =1000Fig.1 Building neighbor adjacency tree based on ITS sequence, Bootstrap =1000
将9 种乌头聚类,观察9 种乌头的亲缘关系. 基于ITS K2P(Kimura 2-parameter)模型构建的邻接树(图2)显示ZY(遵义)乌头为外类群,BSJ(白水江)、TBS(太白山)、LJL01(老界岭)与LJL02(老界岭)聚为一支,具有较近的亲缘关系;XJ(新疆)乌头与QCHS(青城后山)、LC01(利川)、XE(宣恩)、LC02(利川)、SNJ(神农架)位于二级分支,bootstrap value 为59,LC01(利川)与XE(宣恩)的距离最近.验证了ZY(遵义)的亲缘关系最远,其次是XJ(新疆),BSJ(白水江)、TBS(太白山)、LJL(老界岭)为一类,SNJ(神农架)、QCHS(青城后山)、LC(利川)和XE(宣恩)为一类.
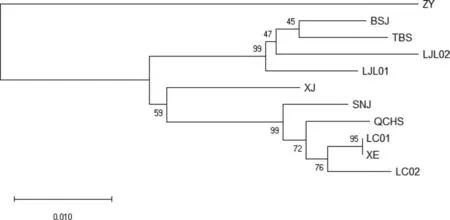
图2 9 种不同产地乌头属植物ITS 邻接树,Bootstrap =1000Fig.2 9 species of Aconitum ITS neighbor tree, Bootstrap =1000
3.3 ITS1、ITS2 一致性二级结构
二级结构不同颜色的碱基配对代表此处有不同数量的配对类型,而颜色的深浅则代表此配对在所有组内种中的保守性,颜色越浅则此配对在组内的差异越大(图3). ITS1 与ITS2 的二级结构上都是一个多分枝环,周围有4 条螺旋臂(Helix).从图3 中的结果显示ITS1 比ITS2 的配对稳定性小,9 种乌头属植物ITS1 变异位点有89 个,占35%,ITS2 变异位点有53个,占23%,表明ITS1 比ITS2 区域有更高的进化速率.
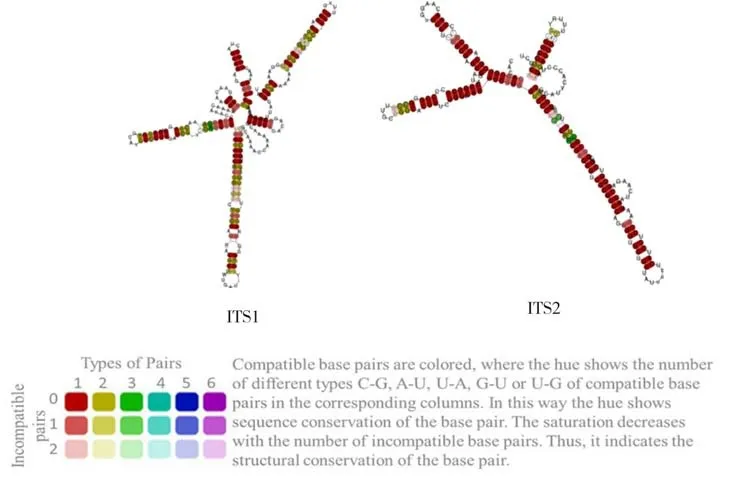
图3 最小自由能预测的ITS1、ITS2 一致性二级结构Fig.3 ITS1, ITS2 consistent secondary structure with minimum free energy prediction
3.4 ITS1 二级结构分析
所有种都有1 ~2 个多分枝环结构,四个主要的螺旋臂,三个较长的螺旋臂,1 ~4 个较短的螺旋臂(图4).ZY(遵义)的有两个明显的主环,三个较长的螺旋臂,三个较短的螺旋臂.XJ(新疆)的为中心两个较小的分枝环,三个主要的长螺旋臂,三个短螺旋臂.BSJ(白水江)的两个分枝环都在靠近臂I 与臂IV 处,同样有3 条长螺旋臂,短螺旋臂较其他的多. 所有的都在臂II 处中间有两个小环,臂长也大致相同,暗示此段可能为乌头属的特有结构.
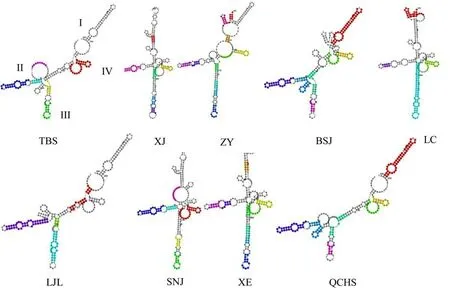
图4 ITS1 的二级结构Fig.4 Secondary structure of ITS1
3.5 ITS2 二级结构分析
在ITS2 Database 上预测二级结构,得到9 个序列的二级结构图(图5).结构符合被子植物一环(主环)四臂(4 个螺旋区)的特征,在臂I 和臂IV 之间有一段比较保守的、富含嘌呤的单链结构. XE(宣恩)和LC01(利川)的臂IV 上有2 个环,其他为1 个环;XJ(新疆)的差异最为明显,主环呈现分裂状,臂II 上为4 个环,臂III 中部有一个较大的环;QCHS(青城后山)的臂I 和臂IV 之间单链长度最短.从整体结构上来看,SNJ(神农架)、TBS(太白山)、LC01(利川)、LJL01(利川)具有高度相似性,在臂III 上都是一个凸出环与5 个小环,在臂IV 末端都为一个环且大小一致,从二级结构上可以把这4 种归为一类;因此可从二级结构的环数与长度来区分这几种乌头.
将ITS2 序列与二级结构信息导入4SALE 软件后,比对后,统计CBCs 位点信息. TBS(太白山)与QCHS(青城后山)和SNJ(神农架)的CBCs 位点都为2,从ITS 一级结构来看,TBS(太白山)与QCHS(青城后山)和SNJ(神农架)位于牛扁亚属的不同分支,亲缘关系较远.在ITS 一级结构中,ZY(遵义)与其他各地乌头亲缘关系都比较远,但在二级结构与BSJ(白水江)、LC01(利川)、LC02(利川)、LJL01(老界岭)、XE(宣恩)存在CBCs 位点. LJL02(老界岭)的CBCs位点较LJL01 都多1 个,除了与TBS 的为0.详细信息见表5.

图5 ITS2 二级结构Fig.5 Secondary structure of ITS2
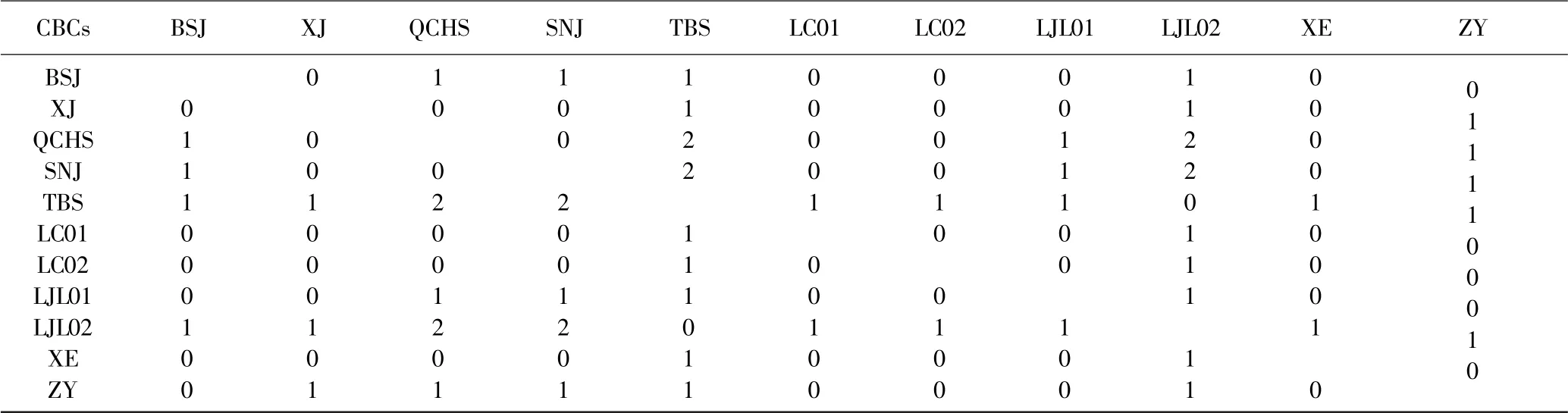
表5 CBCs 位点统计Table 5 CBCs site statistics
4 讨论
在生物系统进化的过程中,高级结构与功能有更加密切的联系,通过ITS1 一级结构与ITS2 二级结构预测来阐明它们的亲缘关系是十分必要的. 同时,已有研究表明ITS2 对系统进化树的误差进行更精确的修正重构是可行有效的[23],在许多以二级结构分析为工具来进行系统进化的研究中,RNA 的折叠结构常用来改善多序列的比对.虽然序列比对的结果还不够完全正确,但这种方法确实能够提高序列比对的准确性,加强系统发育的重建.
本研究中的9 个材料显示,ITS1 与ITS2 一级结构显示GC 含量都高于60%,ITS1 的变异位点数高于ITS2,ITS1 简约信息位点占12.4%,ITS2 简约信息位点9%. 同时ITS1 与ITS2 的一致性二级结构表明,ITS2 的碱基配对稳定性更高,ITS1 的结构变化更大.综合一级结构和二级结构的比对情况,认为ITS1 的结构变异程度高于ITS2 的结构变化,ITS2 序列具有更高进化保守性.
ITS 一级结构的聚类及遗传距离显示,ZY 乌头与其他8 种的聚类亲缘关系都较远,遗传距离最大,从亚属水平上即区分开,BSJ 乌头与XJ 乌头在一级结构上也明显区分开,与形态学分类一致. 通过对9 种乌头属植物的ITS 序列观察可以发现:种间的遗传距离都较小,低于0.1 反映了种间的遗传相似性,LJL01与LJL02 序列之间微小的差异可能是由于种内遗传的多态性所致.6 个产地的高乌头,分为两大类,从ITS 一级结构来看,BSJ(白水江)、TBS(太白山)、LJL(老界岭)的聚为一支,相互之间遗传距离小于0.04,这3 种位于秦岭区域,QCHS(青城后山)、LC(利川)、XE(宣恩)、SNJ(神农架)聚为一支,两两之间遗传距离小于0.03,分布地位于武陵山区,推测高乌头植物在进化过程中,受地理环境的影响产生明显的遗传差异;同时从ITS1 二级结构来看,BSJ(白水江)、TBS(太白山)、LJL(老界岭)具有高度的相似性,LC(利川)、XE(宣恩)、SNJ(神农架)的二级结构形态上能明显区分开;CBCs 位点结果表明这两支的高乌头之间存在多个CBCs 位点,种间杂交率极低,进而验证了这一推测.此前根据ITS 来区分不同种乌头属植物,虽然能相互区分,但是对于种间的区分有限,因此在利用ITS 区分乌头属植物时有必要考虑不同地区的ITS 特征,即可以用于道地药材的鉴定.
通过对ITS1 与ITS2 的二级结构分析比对,不同种具有不同的二级结构特点. ITS1 的螺旋臂II 环数与臂长类似,主要的差异在于分枝环的大小和位置.ITS2 的二级结构差异主要在螺旋臂I 与IV 之间的单链结构,螺旋臂II 和III 类似.因此认为可以用二级结构不同特点来区分不同乌头属植物.
根据Coleman 和Vacquier(2002)提出的CBCcriterion 假说,当两个种间有足够的进化距离累积产生即使一个CBC 在ITS2 二级结构的保守区域上时,它们无法杂交产生后代.9 种乌头两两组合时,54.5%的组合无CBCs 位点,即种间杂交概率高,38%的组合存在1 个CBCs 位点,即无法杂交产生后代,这个结果表明本文中用到的材料存在一定的种间杂交.同时根据以往的乌头属植物研究表明,乌头属植物在形态上类似,并且受环境影响大,即使是同一个种在草地上生长则为直立茎状,在林地下则为缠绕藤蔓状,仅从形态上鉴定容易出错,特别是炮制后的中药材更易产生掺假现象,因此从分子水平上鉴定,具有更加准确的效果.
ZY 的形态学鉴定为白喉乌头,依据中国植物志中记载,白喉乌头在乌头属分类中属于牛扁亚属. 而在本研究中9 个地区的亲缘关系比较中,无论是一级结构比对,还是二级结构分析,ZY 的亲缘关系最远.在一级结构中ZY 与乌头亚属植物聚为一类,其形态上又与牛扁亚属植物接近,推测可能是地理环境导致了此种形态学上的变化;或者从演化进程上来看,牛扁亚属位于较原始的类群,其次是乌头亚属,最后是露蕊乌头亚属,ZY 形态上与牛扁亚属相似,序列上又与乌头亚属的聚为一类,因此推测此种可能是从牛扁亚属到乌头亚属的过渡,但这个推测还需进一步验证.后续将从乌头碱含量来鉴定遵义乌头,为ZY 乌头的道地性提供分子证据与成分含量依据.
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
中成药(2018年11期)2018-11-24
蔬菜(2018年5期)2018-05-17
中成药(2018年2期)2018-05-09
湖北畜牧兽医(2017年9期)2017-09-21
湖北畜牧兽医(2016年4期)2016-07-02
中成药(2016年4期)2016-05-17
湖北畜牧兽医(2016年1期)2016-03-26
杂草学报(2015年2期)2016-01-04
当代党员(2015年9期)2015-09-18
