
藏药金腰属ITS 二级结构的特点及其系统发育研究
2019-05-17 05:40刘秋宇兰德庆易丽莎
西南民族大学学报(自然科学版) 2019年2期
刘 虹,黄 文,刘秋宇,兰德庆,易丽莎,覃 瑞
(中南民族大学武陵山区特色资源植物种质保护与利用湖北重点实验室,湖北 武汉 430074)
目前金腰属在全世界范围约有90 种(The Plant List),亚、欧、非、美四洲均有分布,但主要分布于亚洲温带.生于海拔450 ~4 800 米的林下、高山灌丛、高山草甸和高生碎石隙.我国约40 种,南北均有分布[1-5].Franchet 依据叶子的对生与互生叶的存在为依据,将该属分为Sect. Oppositifolia 和Sect. Alternifolia 两组[6];在此基础上,1957 年Hara 根据茎、叶及萼片等形态特征将金腰属分为了17 个系[7]. 潘锦堂也将金腰属分为金腰亚属(Subgen. Chrysosplenium)和互叶亚属(Subgen. Gamosplenium Maxim.)两大亚属[8],但作为分类学上最困难的群体之一,目前本属内的分类还存在较大问题,需要辅以分子手段来解决.
Nakazawa 和Soltis 使用叶绿体matK 基因对金腰属植物进行了分子系统发育研究,他们发现分子数据与Hara 提出的以前的分类基本一致.然而,其涉及的金腰种类偏少,覆盖面不全[9-10].Kim 等人利用ITS 序列对韩国金腰属的毛金腰系植物进行系统发育分析,结果证实ITS 序列能有效地将毛金腰系中不同的变种区分开,但对不同居群的同一种几乎没有分辨率[2,11].Han 等人也利用ITS 序列结合种皮形态进行系统发育重建,结果证明ITS 构建的系统发育树与种皮形态分类结果一致,说明利用ITS 序列对金腰属植物进行系统发育研究是有效的[12]. Li 等人也通过对6286 份材料进行ITS 序列特征分析,发现ITS 序列适用于虎耳草科的研究[13].
目前,利用分子系统发育的方法虽然仍有很多因素影响会物种鉴定,但是生物系统发生的进化历程在一级结构和二级结构上都会保留进化痕迹,并且高级结构与功能有更加密切的联系,这一层面上的二级结构更能直观地揭示物种进化史[14]. 已有研究认为利用ITS 的二级结构信息对系统进化树的误差进行更加精确的修正重构是可行有效的,特别是在核苷酸序列突变频率较大,但二级结构却很保守的真核生物核糖体ITS2 序列上[15-16]. 在许多以二级结构的分析为工具来进行系统进化的研究中,RNA 的折叠结构常用来改善多序列的比对,虽然序列比对的结果还不能完全正确,但这种方法确实能够提高序列比对的准确性,加强系统发育的重建[17]. 当前在与ITS 二级结构相关的研究中,补偿性碱基配对变化(compensatory base pair changes, CBCs)开始越来越多地应用到种间和种内关系的研究中[18-19].由于rRNA 的多个拷贝之间有协同进化的趋势,因此CBCs 可作为两个居群之间没有发生基因交换的重要指标. 这个假设被称为CBC criterion,此后Muller 等人用使用了30 万个ITS2二级结构对此假设进行了验证,通过综合分析后他们得出了两个独立的结论,如果两个个体之间存在CBC,那么它们是两个不同的种的概率约为93%;当两个个体之间不存在CBC,那么它们是同一种的概率约为76%[20].本研究旨在通过分析核糖核酸内部转录间隔区(ITS)的DNA 序列,以解决金腰属中两大亚属的分类问题.
1 材料和方法
实验材料共选取金腰21 种,其中8 种来自NCBI,包含的外类群金腰的近缘属涧边草属的涧边草(Peltoboykinia tellimoides),其它13 种由实验室取自武陵山区,实验材料来源见表1.
1.1 金腰基因组DNA 的提取
金腰干燥的样品各取15 mg,在液氮中研磨成粉.采用改良的CTAB 法提取各样品的总DNA[21]. 用紫外分光光度计检测DNA 浓度,再用1%琼脂糖电泳检测质量,-20 ℃冰箱中保存备用.
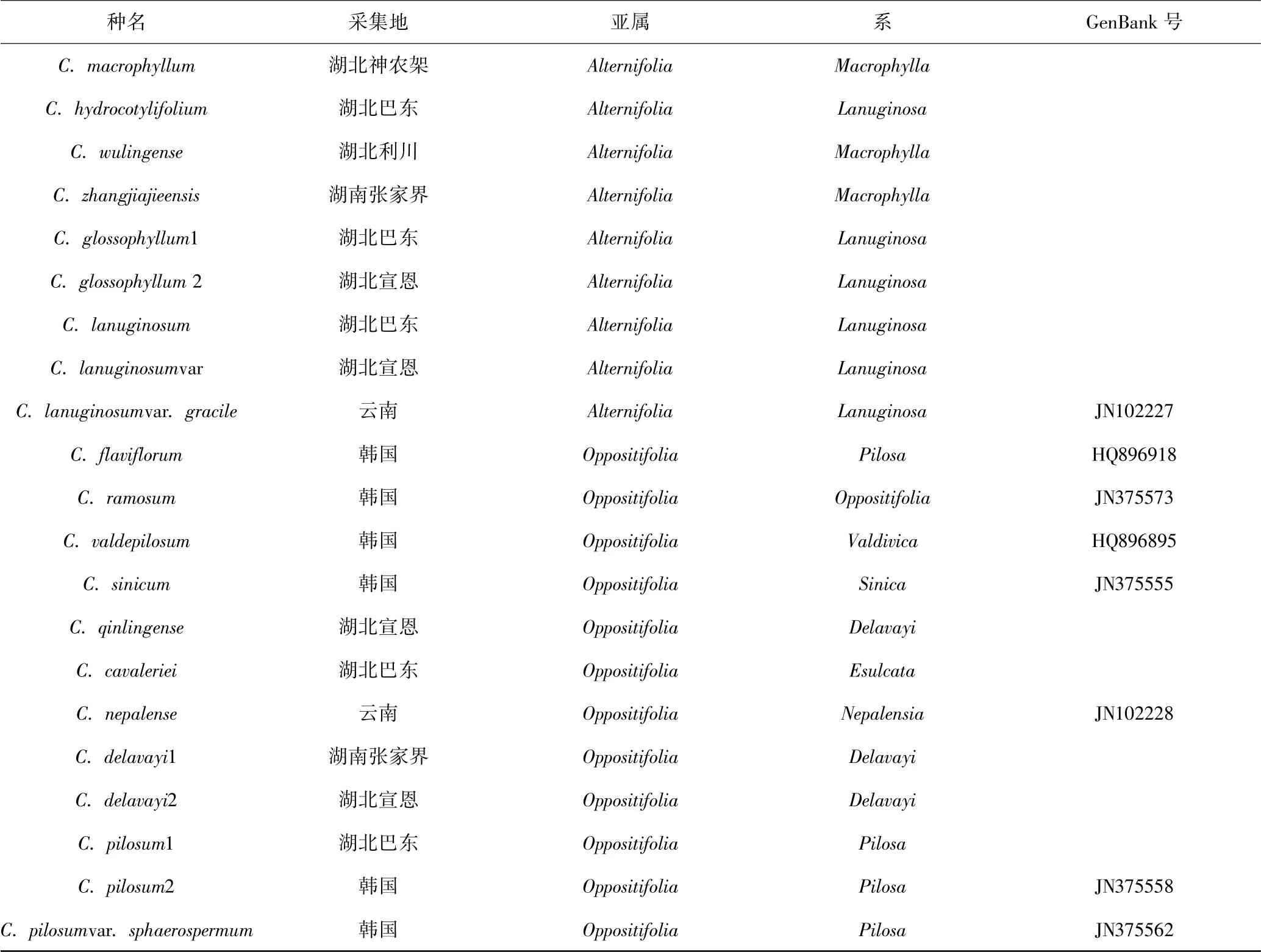
表1 实验材料基本信息Table 1 Information of the experimental materials
1.2 PCR 扩增测序与数据处理
选取在系统发育中被广泛应用的ITS 序列扩增,引物序列为:
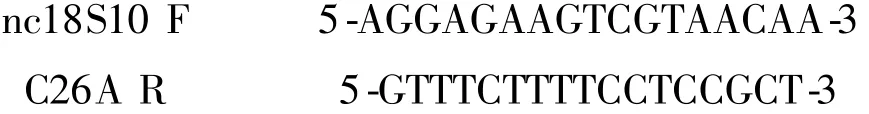
PCR 扩增体系参考Kim 等人[11],将样品送至北京华大科技有限公司测序.所得序列用Mega6.0 软件进行剪接与校队[22],并进行Clustal W 多序列比对,分析碱基组成,GC 含量,计算遗传距离(K2P),并用ML 法(Maximum Likelihood)构建最大似然树,所有树均以1000 次bootstrap 检验树分支支持率. 再用Mrbayes 软件构建贝叶斯(Bayes)树[23-24].ITS2 核心二级结构的预测采用ITS2 database[25-26]. CBCs 统计分析使用软件4SALE[27].
2 结果与分析
2.1 金腰属ITS 序列特征
分析结果显示ITS 序列总长为643 ~653 bp,检测到保守位点392 个、变异位点273 个、简约信息位点176 个.与ITS1 和ITS2 相比,5.8S 的变异位点和信息位点都极少,显示出这一区域在长度和碱基变化上都非常稳定,多数集中在ITS1 和ITS2 区域.从表2可看出,在对生叶的种间比较,其ITS1 的GC 含量比ITS2 高,在互生叶的种间比较正好相反,ITS2 的GC含量比ITS1 高,而在对叶亚属与互叶亚属间比较,对生叶ITS1 的GC 含量比互生叶高,在ITS2 序列中相反,互生叶的GC 含量比对生叶高.在对生叶的种中,中华金腰C. sinicum 和山溪金腰C. nepalense 这两个种的GC 含量偏高,特别是在ITS2 序列中,其GC 含量明显高于其它对生叶的种和平均值,并且在之后的系统分类中,这两个种的分类位置也较特殊.
从表3 可看出除了中华金腰C. sinicum 和山溪金腰C. nepalensel 这两个种以外,互生叶种之间的种间遗传距离和对生叶种之间的种间遗传距离比互生叶种和对生叶种之间的遗传距离要小.而C. sinicum和C. nepalense 这两个种表现出来的特征却正好相反,它们都属于对叶亚属,但与互生叶种的种间遗传距离较小,与对生叶种的种间遗传距离却较大.

表2 金腰属对叶亚属与互叶亚属的ITS 序列长度(bp)与GC 含量(%)Table 2 The length(bp) and GC content(%) of Chrysosplenium ITS sequence
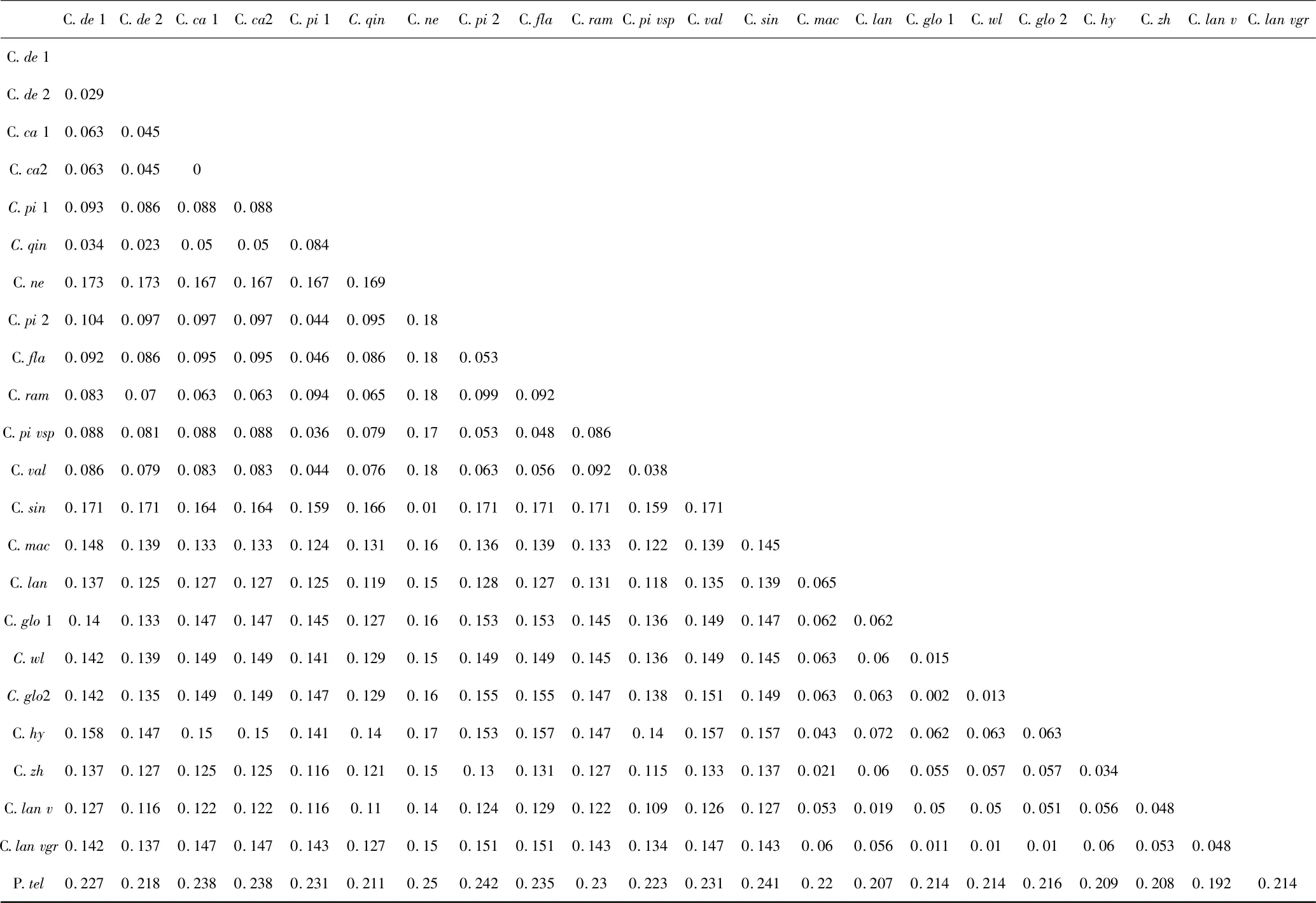
表3 金腰属种间遗传距离Table 3 Genetic distance among species of Chrysosplenium
2.2 金腰属系统发育分析
基于ML 法和Bayes 法构建了系统进化树,将它们的聚类情况结合形态分类学互相比较分析,如图1和图2 结果显示ML 树与Bayes 树的聚类结果都能将互叶亚属与对叶亚属区别开并聚类在一个较大分支中,共同将C. sinicum 和C. nepalense 这两个对生叶的种被一起聚类到互叶亚属和对叶亚属并列的另一分支中,结合这个两个种的GC 含量明显高于其它种且遗传距离也与其它种较远的情况,这个分类结果可能说明这两个种比其它种更为原始或者其进化速率更缓慢,它们的ITS 序列更为保守,结构更稳定.但在节点给出的置信度上,Bayes 树普遍比ML 树高,如C.delavayi 1 与C. delavayi 2 这两个种的小分支节点ML树置信度为70%,Bayes 树为89%;C. flaviflorum、 C.pilosum 1 和C. pilosum2 三个种组成的分支的节点置信度在ML 树上为68%,Bayes 树为97%.Bayes 法给出进化树与形态学分类结果较为接近,其置信度和种间的分辨率也较高.

图1 基于最大似然法ML 对金腰属种构建的进化树Fig.1 Maximum Likelihood tree of Chrysosplenium
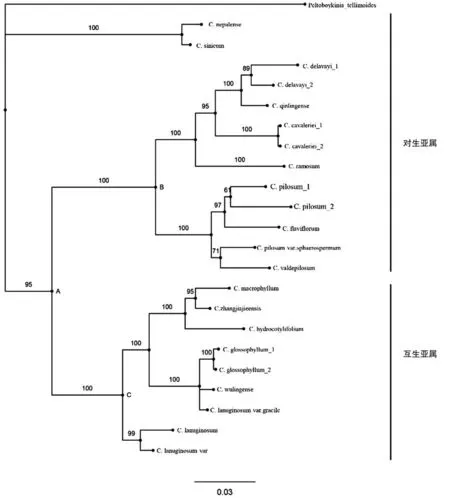
图2 基于Bayes 法对金腰属种构建的进化树Fig.2 Bayes tree of Chrysosplenium
2.3 ITS2 二级结构和CBCs 分析
在图3 中,ITS2 database 的motif 检测结果显示出三个模体的存在(Motif I-III).Motif 显示在灰色区域,绿色背景为保守区域,从图中可看出这两种区域几乎都分布在HII 和HIII 中,这种分布可能对其形成这两个螺旋臂有重要作用,特别是对真核生物ITS2 的二级核心结构的形成提供保障,尤其在HII 臂中,Motif I和II 几乎覆盖了大部分结构以及它周围的位点.位于HIII 的T3 发卡环在所有样本中分为两种结构T3a 和T3b,是由C.pi 2、C.pi 1、C.glo 1、C.glo 2 三个种在图中159 位点处发生U-C 碱基替换而形成大小不同的发卡环.同时在其它螺旋臂上也存在一些较保守的变异位点,这些变异位点都只在个别的种里发生了变化,如图中第40、160、164 位点处,虽然这些位点改变,但对二级结构未造成影响,虽然目前它仅在少数种中发生变化,但笔者推测在之后的进化过程中,此类位点可以被允许较频繁的发生变异. 此外,位于第54、123 的位点变化也表现出在互叶亚属和对叶亚属中组内一致,组间不同的特征,第54 位点在对叶亚属中为G 碱基,互叶亚属中为U 碱基;第123 位点在对叶亚属中为A 碱基,互生叶为G.此外,涧边草的ITS2二级结构位于图3 左上方,其整体结构与金腰属几乎没有差异.
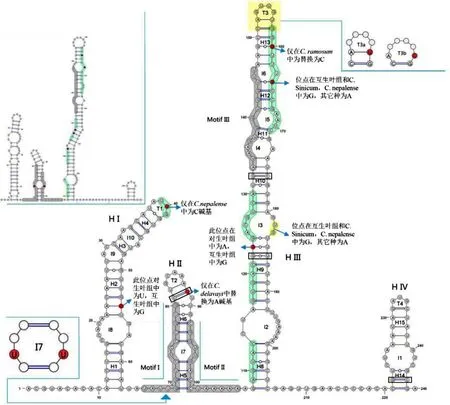
图3 ITS2 一致性结构分析图Fig.3 The predicted consensus secondary structure of the ITS2
在ITS2 二级结构中CBCs 配对(图3 中黑框碱基对)分布在HII、HIII、HIV 臂中,共有4 对.在表6 中,列出了金腰属21 个种和变种两两对应的CBCs 数量,从中 可 看出 C. pilosum var. sphaerospermum、 C.valdepilosum、C. pilosum 1、C. pilosum 2 、C. flaviflorum
这5 个种和其余至少15 个种都有CBCs,表4 显示了在互叶亚属内的种间没有发现CBCs,与之不同的是,对叶亚属内(表5)的种都存在与其它几个种有CBCs的情况,但在原变种与变种之间未发现CBCs. 此外,C. sinicum、C. nepalense 这两个种互相之间未发现CBCs 存在,但它们与其余对生叶种之间都有CBCs 存在,而它们在互叶组中,除了C. davidianum,与其余种都没有CBCs 发现(表4-6).
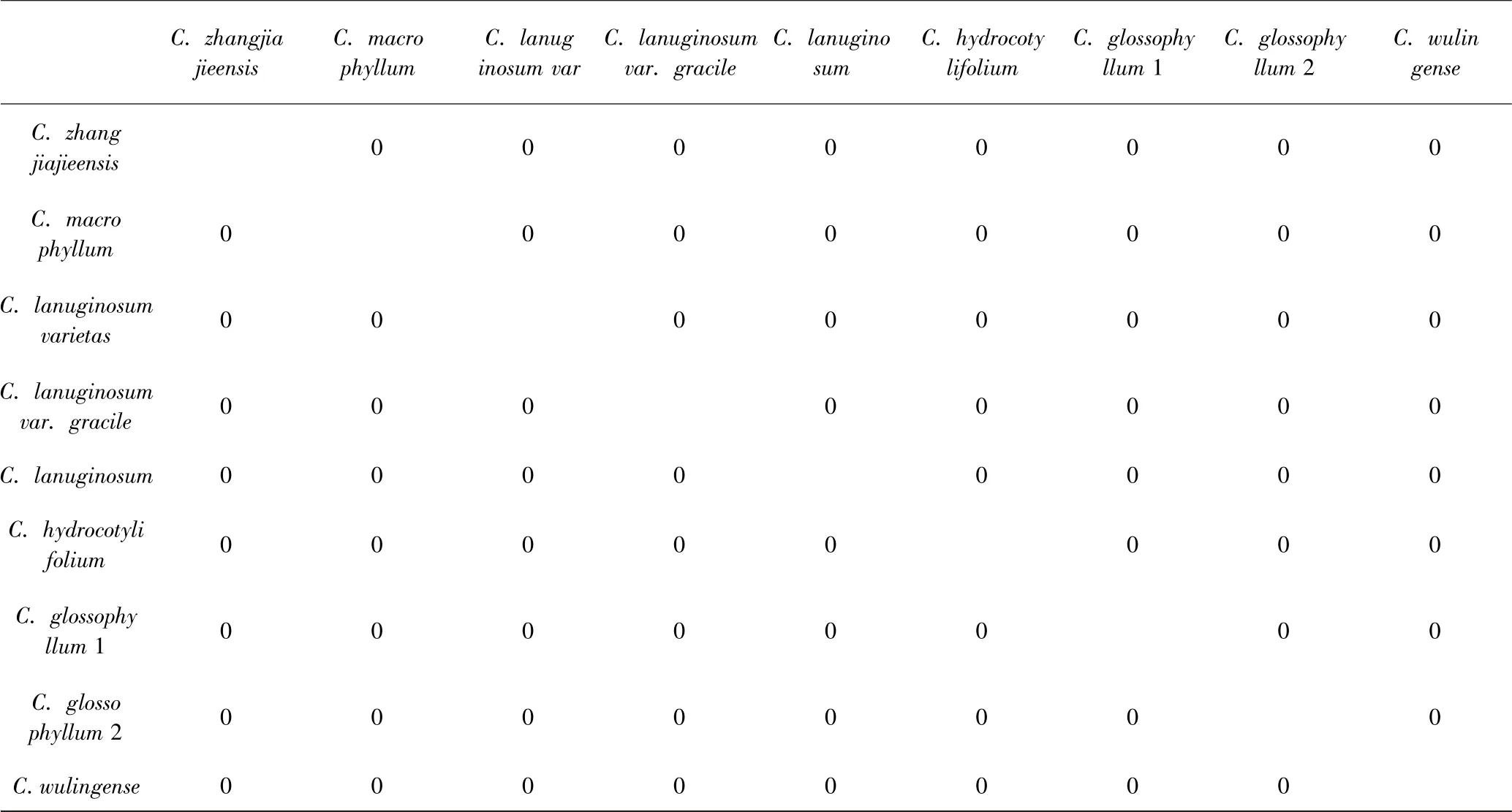
表4 金腰属互叶亚属ITS2 CBCsTable 4 ITS2 CBCs in Alternifolia of Chrysosplenium
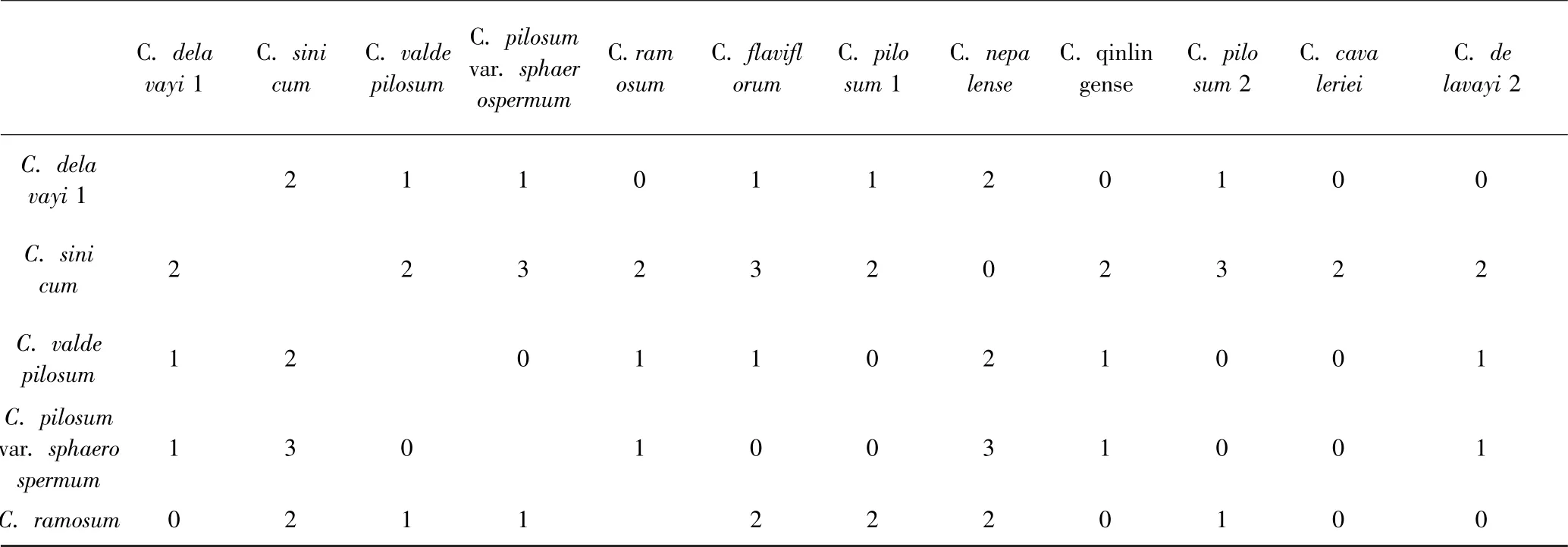
表5 金腰属对叶亚属ITS2 CBCsTable 5 ITS2 CBCs in Oppositifolia of Chrysosplenium

C. flavi florum 1 3 1 0 2 0 3 1 0 0 1 C. pilo sum 1 1 2 0 0 2 0 2 1 0 0 1 C. nepa lense 2 0 2 3 2 3 2 2 3 2 2 C. qinlin gense 0 2 1 1 0 1 1 2 1 0 0 C. pilo sum 2 1 3 0 0 1 0 0 3 1 0 1 C. cava leriei 0 2 0 0 0 0 0 2 0 0 0 C. dela vayi 2 0 2 1 1 0 1 1 2 0 1 0

表6 金腰属ITS2CBCs 比较Table 6 ITS2 CBCs in Chrysosplenium
3 讨论
本文结果表明,对生叶的中华金腰C. sinicum 和山溪金腰C. nepalensel 两个种的特征与其它对生叶种和互生叶种之间的整体差异不符.如:(1)其它对生叶种ITS2 中的GC 含量是明显低于互叶亚属的,而它们的GC 含量甚至比互生叶的种都要高.它们在整个ITS 区域的GC 含量也明显高于其它19 个种. (2)在金腰属种间遗传距离的分析中,我们发现互生叶种内之间的种间遗传距离和对生叶种内之间的种间遗传距离较小,而互叶亚属和对叶亚属之间的遗传距离较高.但C. sinicum 和C. nepalense 虽然是对生叶植物,但与此特征正好相反,它们与互生叶的遗传距离更小.
在系统进化树的聚类分析中,这两个种并未被归于互生叶或对生叶的分支中,而是单独被归于另一与对生、互生分支并列的分支上.在ITS2 二级结构的分析上,我们也标注出了这两个种与其它种都不相同的碱基配对位点,在l3 环上我们还发现了只有这两个种存在碱基插入情况,也或者是其它种在这个位置碱基的集体缺失.在CBCs 的分析中,我们在ITS2 二级结构中它们与对生叶种中都有CBCs,与多数互生叶种却没有,而其它对生叶的种都或多或少与几个互生叶种存在CBCs.
因此,C. sinicum 和C. nepalense 这两个种可能是金腰属种最原始的物种,它们在形态学与对叶亚属相符,但分子序列的特征上却与互叶亚属更相近. 笔者推测,C. sinicum 和C. nepalense 在进化上可能处于更靠近金腰属基部的位置,因此,C. sinicum 和C. nepalense 这两个种可能为金腰属中的原始种,而后分化产生了互生亚属和对叶亚属.
除了中华金腰C. sinicum 和山溪金腰C. nepalensel 这两个种之外,在本实验中,互叶亚属和对叶亚属在分子水平的多个方面表现出稳定的差异,这两个组之间从ITS 序列特征、种间遗传距离和二级结构的信息上都能被区别出来,如两组在序列上存在一些较特殊的位点,这些位点的碱基在两组间不同,在两组内一致,其中最明显的是在5.8S 上的一个位点,这个位点的差异形成了两组间5.8S 二级结构最大差异结构,由于5.8S 的二级结构与蛋白质的合成相关联,两组间这种结构的差异可能与其功能有关. 通过对CBCs 的分析,我们判断对生亚属之间发生杂交的情况和可能性远比互生亚属要少,对生亚属似乎更趋向于各自独立进化,互生亚属可能更容易互相杂交产生变种甚至是新种. 从金腰属ITS 序列提供的信息来看,对生亚属之间的差异明显要比互生亚属之间的差异要大,这可能也说明对生亚属由于各自独立进化,其DNA 的差异也呈增大的趋势,而互生亚属之间由于杂交的可能性高,它们的DNA 能互相融合减小差异或者缓解差异增大的可能性也更大.这可能揭示了金腰属植物叶对生和互生这两种类型的种的进化方式存在差异,随着时间推移它们的差异可能会越来越明显最终表现出完全不同的进化方式,而这种不同的进化方式会导致它们的后代差异越来越大甚至可能分化形成不同的属.
猜你喜欢
区域治理(2022年40期)2022-11-27
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
上海金属(2021年6期)2021-12-02
昆明医科大学学报(2021年3期)2021-07-22
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
生物学通报(2019年3期)2019-02-17
电脑知识与技术(2018年19期)2018-11-01
蔬菜(2018年5期)2018-05-17
