
桉树种植对林地土壤丛枝菌根真菌群落结构及多样性的影响
2019-05-31 01:05李佳雨林家怡裴晨羽黄锐洲唐光大
生态学报 2019年8期
李佳雨,林家怡,裴晨羽,黄 林,黄锐洲,唐光大
华南农业大学林学与风景园林学院/中国南方石灰岩植物研究中心, 广州 510640
桉树是桃金娘科(Myrtaceae)桉属植物的总称。由于其环境适应能力强、生长迅速、轮伐期短、产量高,具有较高的经济价值,已被广泛引种栽培[1- 2]。目前,全世界约5000万hm2的热带人工林中,桉树的种植面积占了40%[3- 4]。中国是世界上第二大桉树生产国,截至2016年,种植面积已经达到368万hm2,预计仍将以每年20万hm2的速度增长[5]。在产生巨大经济效益的同时,桉树种植带来的林下生物多样性降低、生物栖息地减少等生态问题也不容忽视[6- 8]:一方面,桉树根系和凋落物产生的化感物质直接或间接影响了土著植物的生长及光合作用等[9- 13];另一方面,桉树耗水耗肥量大,而大量的人工施肥改变了原生土壤的养分循环节奏,长期种植可能引起土壤质量变化。而在土壤质量变化过程中,理化特征和生物学性质等均会产生敏感响应。已有的研究结果显示,桉树的连续种植会导致土壤有机碳和全氮含量降低,同时降低土壤微生物的生物量碳和氮含量[14- 15];而随着种植年份的增加,土壤细菌群落结构会发生显著变化[16- 18]。
AM真菌隶属于球囊菌门,普遍存在于土壤环境中,能够与80%以上的陆生植物形成共生体系[19],在利用植物提供的有机养分的同时,促进宿主植物对氮、磷等营养物质的吸收,提高植物抗逆抗病能力[20- 21],承担着养分运输、能量流动和信息传递等生态功能,对生物多样性的维持和生态系统稳定具有重要作用。同时,AM真菌可以减轻入侵植物对本土植物的消极影响,抑制入侵植物的化感作用,以维持入侵地的生态平衡[22- 23]。AM真菌群落结构也会受土壤类型、海拔高度、坡向、水分状况和肥力等的影响[24- 25],植被类型和土壤理化性质的改变会造成AM真菌群落发生变化。例如,长期种植毛竹(Phyllostachysedulis(Car.) J. Houz.)会使 AM真菌生物量显著降低,群落结构明显改变,不利于土壤碳固存和生态系统稳定[26];在人工种植柚木(TectonagrandisLinn.)的林地土壤中,也发现真菌孢子密度发生了显著变化[27]。而对于桉树人工林中AM真菌群落的变化特征,尚缺乏足够的了解。
开展桉树种植对林地土壤AM真菌影响的相关研究,探讨大面积人工纯林种植对土壤微生物环境的长期影响,对森林经营和植被恢复等具有较强的指导意义。本研究比较了次生林改种桉树后,不同种植年份的桉树林中土壤AM真菌种类、群落组成及多样性的变化,试图厘清桉树林土壤中AM真菌群落随种植年限的变化趋势,以期为桉树林的栽培管理和环境修复等提供理论依据。
1 研究样地概况
研究样地位于广东省惠州市惠阳区镇隆镇(22°57′N,114°21′E),属于亚热带季风性气候,年平均降雨量约2000 mm,集中在4月至9月,年平均温度为22℃,降雨量充足,光照丰富。试验地选取皆伐种植的桉树林,并以邻近的次生林作为对照。桉树林下最常见的植物有盐肤木(RhuschinensisMill.),光叶山黄麻(TremacannabinaLour.),山麻杆(Alchorneatrewioides(Benth.) Muell. Arg.),春花(Rhaphiolepisindica(L.) Lindl. ex Ker),展毛野牡丹(MelastomanormaleD. Don),芒草(MiscanthussinensisAnderss.),山菅兰(Dianellaensifolia(L.) DC.),异叶鳞始蕨(LindsaeaheterophyllaDry.)等;次生林中常见植物有马尾松(PinusmassonianaLamb.),鼠刺(IteachinensisHook. et Arn.),鸭脚木(Scheffleraheptaphylla(Linn.) Frod.),山乌桕(Sapiumdiscolor(Champ. ex Benth.)Muell.-Arg.),黑面神(Breyniafruticosa(Linn.) Müll. Arg.),桃金娘(Rhodomyrtustomentosa(Ait.) Hass.),梅叶冬青(Ilexasprella(Hook. et Arn.) Champ. ex Benth),春花,芒草,异叶鳞始蕨,芒萁(Dicranopterispedata(Hout.) Nakai.),鬼灯笼(ClerodendrumfortunatumLinn.),乌毛蕨(BlechnumorientaleLinn.)等。
2 试验方法和材料
样本采集于2017年3月,在3个相邻但种植年份不同的桉树林(2年生、5年生和10年生)中进行,同时在邻近的次生林样地采集对照样本。按照“S”型曲线在坡向相同、海拔相近(110—120 m)处设置10 m×10 m样方,每个样地设3个样方,参照Qin等和Carrillo-Saucedo等的方法[28-29],样方内以“5点取样法”采集表层土壤(0—15 cm)共计1 kg左右;同时在不伤及植物根系结构的前提下,在样方内采集桉树根样,放入FAA(Formalin-acetic acid-alcohol)固定液中。将同一样方内的土壤样品混合均匀,并剔除植物残体及石砾,放入无菌封口袋,置于冰盒中迅速带回实验室。土壤样品过4 mm筛后分成2份,保存于4℃冰箱:1份用于土壤理化性质的测定,另1份用于土壤AM真菌孢子的分离和鉴定。
2.1 土壤理化性质分析

2.2 AM真菌孢子鉴定与侵染率测定
采用湿筛倾析-蔗糖离心法[31]分离土壤AM真菌孢子,压片,根据其形态特征,参考国际VA菌根网站(https://invam.wvu.edu/home)的鉴定图片,形态描述及最新分类标准进行种属的检索和鉴定。
菌根侵染率测定步骤如下:将采集的根样从FAA固定液中取出,剪成约1 cm长的根段,置于装有10% KOH溶液的玻璃瓶中,放在90℃水浴锅中消解至透明(根系产生明显气泡);取出根样,放入锥虫蓝乳酸甘油溶液(锥虫蓝0.24 g;乳酸10 mL;甘油160 mL;蒸馏水160 mL)中,染色15—60 min,将根段整齐摆放在载玻片上,盖片后镜检。菌根侵染率的计算采用根段频率常规法[32],在Leica光学显微镜下观察每条根段的侵染情况,根据侵染点、菌丝、丛枝和泡囊等菌根结构所占比例,分为0—5共6个等级,然后根据丛枝和泡囊在菌根结构中的有无及丰富度,将1—5级划分为A0,A1,A2,A3共4个级别。统计不同桉树种植年份中AM真菌菌根侵染情况,每个年份随机选取30条根段进行镜检计数,最后将数据输入MYCOCALC软件计算菌根侵染率。
2.3 数据分析
2.3.1AM真菌群落结构特征指标:
(1)AM真菌孢子密度和种丰度
孢子密度:指每100 g干土中孢子或孢子果的数量,其中1个孢子果按照1个孢子计数。
种丰度[33]:物种丰富度,以每个土壤样本中AM真菌的物种数目表征。
(2)物种多样性指数和均匀度
本文采用香农指数、辛普森指数和均匀度表示物种多样性及分布情况。
香农指数(H)=-∑(PilnPi)
辛普森指数(D)=1-∑(Pi)2
均匀度指数(J)=H/lnS
式中,Pi表示土样中种i的AM真菌孢子数ni与孢子总数N的比值;S为土样中AM真菌种的数目。
2.3.2统计分析
采用“Microsoft Excel 2017”软件对土壤理化性质、孢子密度、种丰度、物种相对丰度和多样性指数等数据进行录入及初步分析;采用SPSS 19.0进行单因素方差分析(不同组间土壤性质和AM真菌丰度差异等);采用非度量多维尺度法(Nonmetric Multi-Dimensional Scaling,NMDS)对Bray-Curtis距离进行降维分析以解析群落结构;进一步通过冗余分析(Redundancy Analysis, RDA)解析不同环境因子对于AM真菌群落结构的影响,并通过999次蒙特卡洛置换来检验因子的显著性。Bray-Curtis距离计算、矩阵降维及RDA均在R 3.3.3中使用 “vegan”包计算[34]。
3 结果与分析
3.1 桉树种植对土壤理化性质的影响

表1 不同桉树种植年份土壤理化性质
同一列不同字母代表组间差异显著(P<0.05)
3.2 桉树种植对土壤AM真菌群落的影响
3.2.1AM真菌种类变化
从所有的检测样本中鉴定出AM真菌共9属18种,其中无梗囊霉属Acaulospora6种,斗管囊霉属Funneliforms4种,球囊霉属Glomus1种,原囊霉属Archaeospora1种,和平囊霉属Pacispora1种,近明球囊霉属Claroideoglomus1种,盾巨孢囊霉属Scutellospora1种,根孢囊霉属Rhizophagus2种,隔球囊霉属Septoglomus1种(表2)。次生林土壤中的AM真菌种类最多(15种),2年生、5年生和10年生桉树林土壤中分别分离出11种、9种和13种AM真菌,均少于次生林真菌种类。其中,A.bireticulata、Acaulosporasp1、F.mosseae、F.geosporus、F.verruculosus、Funneliformssp1和Glomussp1在次生林和所有桉树林土壤中均有发现,Pacisporasp1和Scutellosporasp1只在次生林中存在。另外,A.spinosa、Archaeosporasp1、Claroideoglomussp1和Rhizophagussp2在5年生桉树林中未发现,却在2年生、10年生桉树林和次生林中存在。A.capsicula和S.deserticola只在桉树林中存在,在次生林中未发现。
3.2.2AM真菌孢子密度和种丰度变化
在2年生、5年生、10年生桉树林及次生林土壤中,每100 g干土中孢子个数分别为2520个、1553个、2953个和6233个。整体而言,桉树林土壤中的真菌孢子密度显著低于次生林,在2年生桉树林土壤中明显下降,5年生土壤中降到最低,但在10年生桉树林土壤中得到一定程度恢复(图1)。AM真菌种丰度变化与之类似,桉树林的土壤AM真菌种丰度明显低于次生林(11.33),随种植年份的增加逐渐下降,在10年生桉树林土壤中亦有所恢复(图2)。

表2 不同桉树种植年份土壤AM真菌群落组成变化
“+”表示有,“-”表示无

图1 不同桉树种植年份土壤AM真菌孢子密度 Fig.1 Spore density of soil AM fungi in differentEucalyptusplanting years

图2 不同桉树种植年份土壤AM真菌种丰度 Fig.2 Species richness of soil AM fungi in differentEucalyptusplanting years
3.2.3不同种植年限的桉树林土壤中AM真菌菌根侵染变化
5年生桉树根系的AM菌根侵染率最高,其次是2年生,而10年生的根系菌根侵染率最低;丛枝丰富度方面,5年生桉树根系的丰富度最高,其次是10年生、2年生(表3)。
3.2.4AM真菌孢子多样性和均匀度
土壤中AM真菌在桉树种植2年时香农指数和辛普森指数分别最高,而种植5年时均为最低,但单因素方差分析比较结果显示:组间多样性指数差异不显著(P<0.05)。桉树种植2年的土壤中AM真菌均匀度最高,显著高于5年生及次生林土壤,但与10年生的土壤相比差异不显著(表4)。

表3 不同桉树种植年份根系侵染率(%)
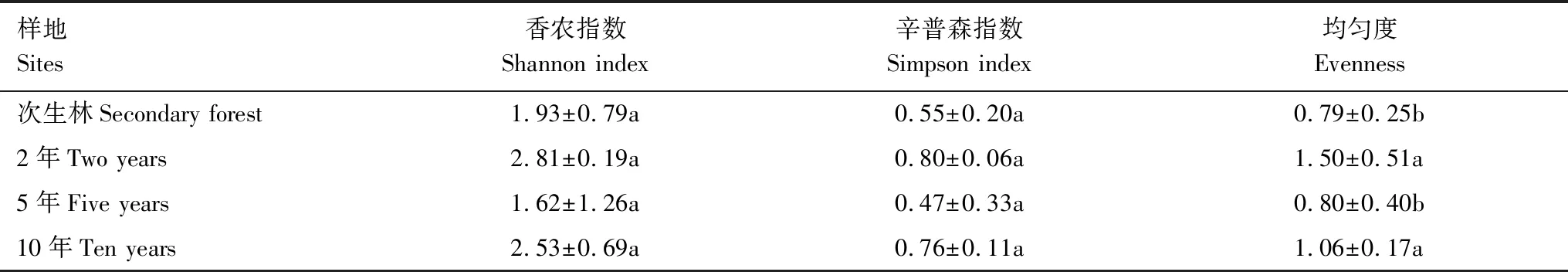
表4 不同桉树种植年份土壤AM真菌孢子多样性和均匀度
3.2.5AM真菌群落结构

图3 不同桉树种植年份土壤AM真菌非度量多维尺度分析 Fig.3 Nonmetric Multi-Dimensional Scaling (NMDS) of AM fungi in differentEucalyptusplanting years
非度量多维尺度分析(NMDS)结果显示,桉树林与次生林的AM真菌群落组成存在明显差异,相互距离较远,尤其是2年生和5年生桉树林的样本,差异最大;在不同种植年份中,AM真菌群落组成不同,在图中分布在不同的位置(图3)。
根据不同组间孢子相对丰度的计算和单因素方差分析,次生林、5年生和10年生桉树林土壤中优势种为Funneliformsgeosporus,但在2年生土壤中显著降低;而2年生的优势种为Septoglomusdeserticola,平均相对丰度为36.28%,显著高于5年生和次生林;Archaeosporasp1在次生林土壤中占比相对较高,为8.65%,显著高于其他桉树林土壤;Funneliformsmosseae在2年生桉树林土壤中占比相对较高,为10.64%,显著高于其他年份的桉树林和次生林土壤;F.verruculosus在2年生和10年生土壤中显著高于次生林;Acaulosporabireticulata的相对丰度在10年生土壤中有显著增长,达到4.7%;Acaulosporasp2在桉树种植2年生土壤中未发现,但在5年生和10年生土壤中有所恢复,且与次生林接近(图4)。另外,Pacisporasp1和Scutellosporasp1仅在次生林土壤样本中出现,且丰度较低,在不同年份之间无明显组间差异。

图4 不同桉树种植年份土壤AM真菌相对丰度Fig.4 Relative abundance of soil AM fungi in differentEucalyptusplanting years

图5 不同桉树种植年份土壤AM真菌与土壤理化因子冗余分析Fig.5 Redundancy Analysis (RDA) between AM fungi community and soil properties in differentEucalyptusplanting years
3.3 环境因子对桉树林土壤中AM真菌群落的影响
土壤AM真菌群落与土壤环境因子的冗余分析(RDA)结果显示(图5),土壤pH、SOC、TN、TK是影响土壤AM真菌群落的主要因子,RDA1解释率为82.3%,表明土壤理化性质是影响土壤中AM真菌群落结构的主要因素,在次生林中与pH、SOC、TN、TK呈正相关关系;RDA2的解释率仅为4.36%,主要影响因子为TK。根据蒙特卡洛置换试验结果,pH、TN、SOC和TK作为具有显著影响的环境因子,在第一轴上的解释率分别为89.88%,57.72%,34.69%和33.2%,表明pH值是土壤中AM真菌群落的主要影响因子。
4 讨论
在陆地生态系统中,土壤微生物能够影响植被生长、群落组成和多样性,同时陆地植被的改变也会影响土壤中微生物生态系统,两者形成动态相互作用的关系。植物通过根系分泌物、凋落物的淋溶、分解、释放化合物质等,直接或间接改变土壤微生物的组成和结构[35]。桉树作为外来树种被广泛引种种植,将物种多样性较高的原生植被替代为桉树纯林,其自身生理代谢和人为管理措施(施肥、灌溉等)都可能使土壤理化性质发生改变,如SOC降低[14,36-37]。本研究结果显示,2年生和10年生桉树林中,土壤SOC显著降低,可能是因为桉树种植对土壤团聚体产生了影响,从而引起土壤中SOC的降低[38];5年生桉树林土壤SOC显著高于2年和10年桉树林,这可能是第5年桉树凋落物积累较多,而土壤微生物分解率低所造成的结果[39],而凋落物淋溶产生的化感物质形成了不利于AM真菌孢子生长和繁殖的环境,导致孢子密度和种丰度显著降低;有研究表明土壤SOC与AM真菌侵染状况呈显著正相关[40],这可能是5年生土壤中AM真菌侵染率较高的原因之一。其他土壤营养元素,如TP、AP等在桉树林中的增加和降低也可能与凋落物的分解、淋溶、矿化等有关[41-42]。
当宿主植被群落发生改变时,土壤中AM真菌的种类和结构可能会随之改变;而当土壤中AM真菌群落改变后,地表植物的种类和数量可能也会改变,植物群落结构和多样性也随之改变[43]。本研究结果证实了桉树种植使土壤中的AM真菌孢子密度和种丰度显著降低,真菌群落发生了显著改变。然而,AM真菌种类和群落改变后,是否引起桉树林下植被和多样性发生改变,尚需进一步开展相关研究。林地土壤中,P含量是影响AM真菌孢子密度的因素之一,与孢子密度呈负相关[44],本研究发现TP和AP含量高的桉树林土壤中,孢子密度相对较低,故土壤营养元素的变化,也会影响AM真菌的种类和多样性。
从AM真菌属种水平分析结果来看,Acaulospora属和Funneliforms属在种植前后均为土壤中AM真菌的优势属,这与其他陆地生态系统中土壤优势属相吻合[45-47],说明桉树的种植对土壤中AM真菌优势属的影响较小。然而,F.geosporus作为次生林中的优势种,在种植桉树后相对丰度显著降低,冗余分析结果显示pH值对土壤AM真菌群落影响较大,可能桉树种植后,土壤酸化对F.geosporus繁殖和生长产生了抑制。Pacisporasp1和Scutellosporasp1在次生林中存在,而在桉树种植后消失,可能与土壤理化因子的变化有关。A.capsicula和S.deserticola是桉树林土壤中分离到,但未在次生林中发现的种类,可能这两种AM真菌对桉树林土壤环境有较强的适应能力。桉树的种植对AM真菌多样性指标影响较小,可能是土壤中AM真菌物种的增加或减少在植被改变后得到了及时补偿,有些真菌减少或消失,但有些真菌又新增。
土壤pH作为衡量土壤状况的一个重要指标受地面植被类型影响较大,桉树的种植容易导致土壤pH值降低[48-49]。本研究结果表明,2年生和10年桉树林土壤pH值显著降低,可能桉树种植加快了土壤中酸性离子的积累[50]。AM真菌孢子密度与土壤pH值呈负相关[51],因此5年生桉树林土壤AM真菌孢子密度和种丰度较低可能与土壤pH升高有关。另外,酸性的土壤环境更适应AM真菌中Acaulospora属的生长[52],这可能是桉树林在2年生和10年生时Acaulospora属孢子的相对丰度均高于次生林和5年生的原因之一。
本研究采用湿筛倾析法分离土壤AM真菌孢子,土壤清洗过程中部分孢子可能会丢失,结果有一定的误差,但鉴定依据较为充分,调查结果可反映AM真菌群落状况,与高通量测序法相比,鉴定更加准确,并能获得孢子密度等统计数据。后期研究中,建议湿筛倾析法与高通量测序方法相结合,充分探讨森林植被改变后土壤真菌群落和多样性的相关变化。森林土壤生态系统中,AM真菌与植物根系形成非常复杂的地下生态网络,森林植被的改变,土壤真菌系统也可能发生变化,相关机制有待深入研究。
猜你喜欢
现代农村科技(2022年5期)2022-11-18
湖南林业科技(2020年5期)2020-12-07
湖南林业科技(2020年2期)2020-09-18
生态环境学报(2019年4期)2019-05-31
南方农业·下旬(2018年5期)2018-09-14
绿色科技(2018年9期)2018-01-29
现代农业研究(2017年11期)2018-01-12
北广人物(2017年2期)2017-03-25
作文周刊·小学六年级版(2016年26期)2017-03-06
现代园艺(2017年20期)2017-02-03
