
不同施氮水平对兴安落叶松化学计量特征的影响
2019-06-07 07:22陈璐王庆贵闫国永邢亚娟
森林工程 2019年2期
陈璐 王庆贵 闫国永 邢亚娟
摘要:以興安落叶松为研究对象,在大兴安岭南瓮河自然保护区建立统一立地条件的4个氮(N)处理水平样地,分别为对照(CK,0g N /(m 2·a))、低N(LN,2.5g N /(m 2·a))、中N(MN,5g N /(m 2·a))、高N(HN,7.5g N /(m 2·a))处理。测定凋落叶、根和鲜叶在不同施N水平下C、N、P、K、Ca、Mg的含量。研究目的是为了阐明兴安落叶松叶和根各元素含量及化学计量比的分配格局,揭示叶片凋落前各元素的重吸收率,探讨各元素及其化学计量之间的关联以及对不同水平施N处理的响应。结果表明:①鲜叶、根、凋落叶C、N、P变化范围分别为393.174~518.362、4.405~23.963、0.743~1.671 mg/g,其中鲜叶C、P含量最高,根最低。②LN,MN,HN均显著提高了根的N含量,MN,HN显著提高了鲜叶N含量。植物养分含量在不同器官中存在显著差异。③植物叶片对Ca表现为富集状态,对N、P、K、Mg各元素则体现为重吸收状态。结论显示,鲜叶和根是影响兴安落叶松C、N、P及化学计量比的主导因子。兴安落叶松在N素充足的条件下,施N导致兴安落叶松对N和P奢侈吸收,提高了凋落叶的分解速率并加快了生态系统的养分循环。
关键词:施N;生态化学计量比;养分重吸收率;兴安落叶松
中图分类号:S718.5文献标识码:A文章编号:1006-8023(2019)02-0001-10
Effect of Long-term Nitrogen Deposition on the Stoichiometric Characteristics of Larix gmelinii
CHEN Lu, WANG Qinggui, YAN Guoyong, XING Yajuan
(College of Agricultural Resource and Environment, Heilongjiang University, Harbin 150000)
Abstract:Taking Larix gmelinii from Greater Khingan Mountains as the research object, four nitrogen (N) treatment level plots with unified site conditions were established in Nanwenghe Nature Reserve of Greater Khingan Mountains, included contrast CK (CK, 0g N / (m 2·a)), low nitrogen (LN, 2.5g N / (m 2·a)), middle nitrogen (MN, 5g N / (m 2·a)), high nitrogen (HN, 7.5g N / (m 2·a)). The contents of C、N、P、K、Ca、Mg in leaves, roots and fresh leaves were determined under different N addition gradient. The purpose was to elucidate the aboveground organ leaves and underground organ roots of Larix gmelini . The distribution pattern of each element content and stoichiometry ratio revealed the heavy absorption rate of each element before leaf litter and discussed the relationship between each nutrient element and response to different levels of N application. The results showed that: (1) C、N、P of fresh leaves, roots and littered leaves ranged from 393.174-518.362 mg/g, 4.405-23.963 mg/g, and 0.743-1.671mg/g. The range of C and P content in leaves was the highest, and the lowest in roots. (2) LN, MN, HN significantly increased N content in roots, and MN, HN significantly increased N content in roots. There were significant differences in plant nutrient content in different organs. (3) The plant leaves were enriched to Ca and reabsorbed to the elements. It conclude that the leaf and root were the main factors that affected the chemical measurement ratio and the C, N, P of Larix gmelinii . Under the condition of sufficient nitrogen, nitrogen deposition resulted in the extravagant absorption of N and P by Larix gmelinii , which could increase the decomposition rate of litter leaves and nutrient cycling in its ecosystem.
Keywords:Application of N; ecological stoichiometric ratio; nutrient recovery rate; Larix gmelinii
0引言
从1980年至2010年间中国各地的氮(N)沉降动态对生态系统产生巨大影响。研究发现在20世纪80年代(13.2 kg/hm 2)和21世纪(21.1 kg/hm 2)之间,平均每年N的体积沉降量增加了约8 kg/hm 2 [1]。人类活动导致大气N沉降不断增强 [2],严重影响生态系统结构功能及其养分循环过程 [3],进而也对植物化学计量的平衡产生影响。
先前的研究表明,植物的化学计量特征可以表征植物维持体内化学计量内稳性的能力 [4],并反映植物对环境变化的适应性 [5]。长期自然选择,使植物进化出一套适应环境变化的策略 [6]。例如,植物能通过调节根系养分的捕获或者叶片养分的重吸收来维持其化学计量平衡 [7]。养分重吸收是指将衰老叶片中的元素吸收到其它功能活跃的植物组织的过程 [8]。养分重吸收是一种重要的内部营养保护机制,可以减少植物对外部养分的依赖 [7]。相反,养分捕获被认为是一种外部养分获取策略。植物对于养分捕获和重吸收的分配方式取决于环境中养分的可利用性和它们所涉及的成本 [9-10]。在缺乏营养的环境中,能量消耗常与养分捕获成比例增加 [11]。但随着养分供应的增加,观察到与养分捕获有关的酶(例如几丁质酶和磷酸酶)的下降 [12]。这也可以被解释为根系获取养分的减少,因为根系对养分的捕获与酶的合成有关。
人为施N可以改变N和其他限制性养分的有效性 [13-14]。从而影响植物对养分的捕获和重吸收 [15-16]。Yuan和Chen [17]最近的一次Meta分析表明,N肥可以提高绿叶中的N浓度,但降低了对N的吸收效率。此外,增加N的利用率也会对其他营养物质的吸收产生影响 [18]。由于N沉积持续会超过植物和微生物对N的需求,土壤中过量的N可能会产生各种有害影响,如土壤酸化、增加硝酸盐的淋溶,以及营养限制的强度和类型的变化 [19]。然而,尽管在元素之间存在化学计量耦合,但尚不清楚植物如何保持养分捕获和养分重吸收的平衡来响应N沉降,所以仍需要进一步的研究和发展。
植物各营养器官养分含量的变异亦对植物生长速率以及应对环境变化起着调节作用 [19]。当前,不同地区植物化学计量格局及其驱动因素仍然是热点话题,其结果对于理解化学计量比的趋同和分异有重要的意义,并且还能够预测生态系统功能,响应全球气候变化。此外,近年来对不同地域空间尺度的研究较多,且大多集中于叶片而忽略了针对植物体不同器官的研究。从丰富资源经济谱理论的角度看,加强不同器官养分化学计量关系,及各器官之间的相关性十分必要 [21]。现如今对化学计量比的研究大多集中在热带、温带森林和草地,对于北方森林研究较少。
基于以上问题,笔者在我国北方森林建立长期施N样地,研究兴安落叶松各元素及化学计量关系在不同施N条件下会发生怎样的变化,兴安落叶松地上部分叶及地下部分根化学计量比的关联性,不同水平施N将如何影响养分重吸收率。研究通过测定不同水平施N下植物鲜叶、根和凋落叶中的C、N、P含量,计算养分重吸收效率,以揭示营养器官叶和根养分元素的分配格局,化学计量比在不同环境下的动态平衡,为进一步研究兴安落叶松对N沉降适应机制提供科学依据。
1材料和方法
1.1研究区概况
实验地点位于大兴安岭南瓮河国家级自然保护区( 125°07'55″~125°50'05″E,51°05'07″~51°39'24″N),最低海拔370 m,最高海拔1 044 m。该地处寒温带,属于寒温带大陆性季风气候,年平均气温-3 ℃,最低气温-48 ℃,最高温36 ℃。年降水量500 mm左右,积温1 500 ℃左右,无霜期90~100 d,植物生长期110 d左右,土壤为暗棕色砂砾土。样地内优势树种为兴安落叶松( Larix gmelinii Rupr.)和白樺( Betula platyphylla),其中兴安落叶松占90%以上。
1.2实验设计
在保护区内选择立地条件相对一致的区域,共设置16块20 m×20 m的实验样地,每两个样方间缓冲带宽为10 m。共设4个梯度4个重复的施N处理,即对照(CK)、低N( LN)、中N( MN)和高N(HN),分别按0、2.5、5.0 、7.5 g /( m 2·a) 进行外源N( NH4NO3)添加处理(不包含大气N沉降量) 。根据N处理水平,于2011年5月起,在植物生长季内(5-9月)每月月初将每个样方所施的NH4NO3溶解于32 L纯净水中,以背式喷雾器人工均匀喷洒在林地上。为保持处理一致性,对照处理则喷洒等量的纯净水。
1.3样品采集和处理
为了研究兴安落叶松化学计量特征对N沉降的响应,于2017年7月植物生长季对样品进行采集,此时植物生长处于旺盛期,鲜叶和凋落叶共存。在每个样方内选取长势良好的3株树,上、中、下3个不同高度向阳和背阴两个方向剪取新鲜叶片混合为一个样品,取同期凋落叶约300 g,并用平板利锹挖取完整的根段,放入写好标签的封口袋中,共取样品36个。对采集完毕的样品先用蒸馏水冲洗至少1次,然后用去离子水冲洗3次,去掉灰尘及泥沙等杂物。清洗后的样品在烘箱里于105 ℃下杀青10 min,70 ℃下烘干至恒重。同一样方内鲜叶、凋落叶、根粉碎后过100目筛,待测。使用碳氮分析仪(Multi N/C 3100,Analytikjene AG,Germany)测定样品C、N含量,使用钼锑抗比色法测定P含量,使用原子吸收分光光度计测定金属元素K、Ca、Mg的含量。在本研究中,以当月凋落叶中大量营养元素的浓度来计算养分重吸收能力 [8,22]。较低的值对应于更高的吸收能力 [23]。营养吸收效率( RE )是以凋落叶和鲜叶中养分浓度的百分之数差来估算的 [8]。叶片养分再吸收效率( RE )的计算方法如下:
RE = (1-C1 /C2 )×100% 。
式中: C1和C2分别代表凋落叶和鲜叶中养分元素的含量。
1.4数据处理
采用Excel 2003和Origin8.5、Spss20.0进行数据处理与制图,利用Spss20.0软件中单因素方差分析方法对4种不同施N水平的样地兴安落叶松鲜叶、根及凋落叶养分元素含量的差异进行比较。分析均采用LSD多重比较不同组的差异;利用Pearson相关系数及线性回归模型分析叶和根之间C、N、P含量及化学计量比之间的关系。文中显著性水平设置为 P = 0.05。C、N、P采用质量分数,C∶N、C∶P、N∶P均采用质量比。
2结果与分析
2.1不同施N水平下兴安落叶松鲜叶、凋落叶、根养分含量及其化学计量比
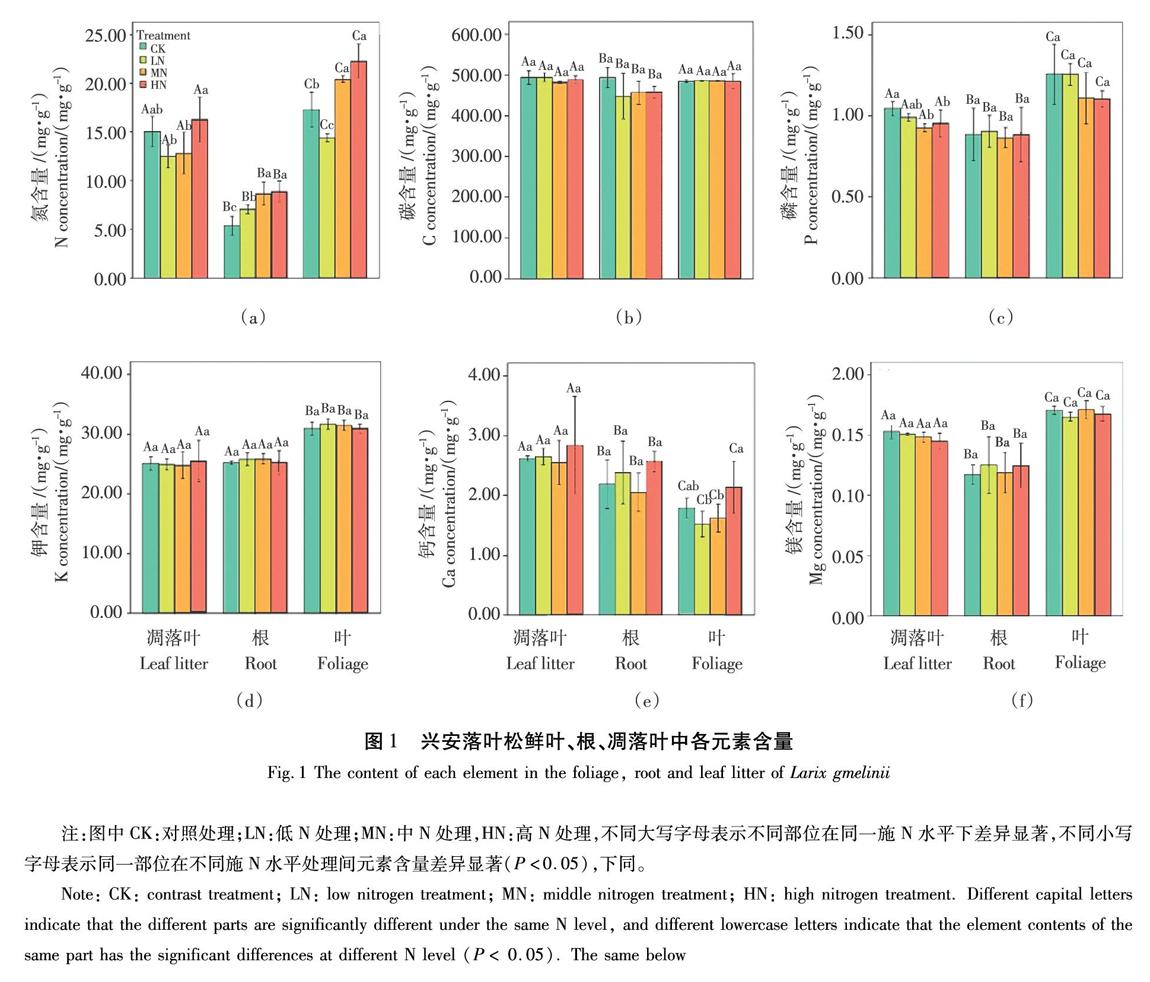
鲜叶、根及凋落叶中C、N、P含量的变化范围分别为393.174~518.362、4.405~23.963、0.743~1.671 mg/g。其中以N的变异最大,变异系数达38.8%,C的变异系数最小,仅为4.5%。金属元素K、Ca、Mg含量的变化范围分别为0.23~0.33、1.32~3.61、0.1~0.18 mg/g。鲜叶和根对C、N、P、K、Ca、Mg含量均有显著影响( P <0.05)(图1)。
不同水平施N下对凋落叶、鲜叶、根C元素含量均无显著影响,如图1(b)所示。HN处理比LN处理下凋落叶的N含量显著提高( P <0.05);LN、MN、HN处理下根N含量均显著提高( P <0.05);MN和HN均提高了鲜叶N含量,如图1(a)所示。MN和HN处理显著降低了凋落叶P含量( P <0.05);各施N水平对根和鲜叶的P含量均无显著影响,如图1(c)所示。凋落叶、根、鲜叶的K、Mg含量在各施N水平下无显著变化,如图1(d)(f)所示。凋落叶和根的Ca含量在各施N水平下变化不显著。鲜叶Ca含量在HN处理下显著高于LN和MN处理( P <0.05),凋落叶和根的Ca含量在各施N水平下变化不显著,如图1(e)所示。
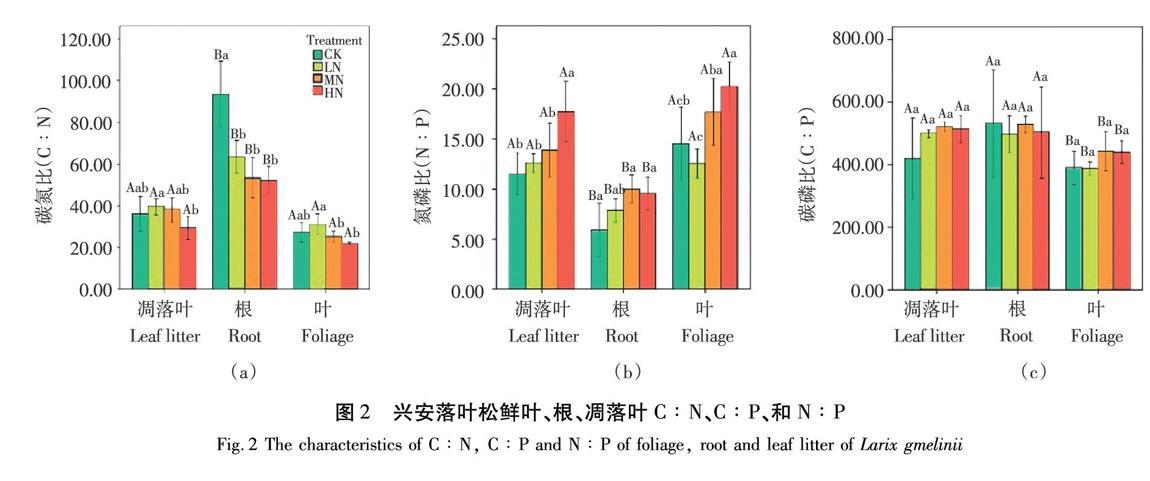
根的C∶N在LN、MN、HN处理下均显著降低( P <0.05)。此外,各施N处理对鲜叶和凋落叶的C∶N呈现相同的趋势,即鲜叶C∶N在LN处理下显著高于HN处理组( P <0.05),凋落叶C∶N在LN处理下显著高于其他处理组( P <0.05),如图2(a)所示。各施N处理对鲜叶、根及凋落叶C∶P均无显著影响,如图2(c)所示。鲜叶N∶P在HN处理下显著高于CK组( P <0.05)。根N∶P在MN和HN处理下与CK相比显著提高( P <0.05)。凋落叶N∶P在HN处理下均显著高于其它处理组( P <0.05),如图2(b)所示。
2.2不同施N水平下兴安落叶松鲜叶、凋落叶、根养分含量及化学计量比相关性
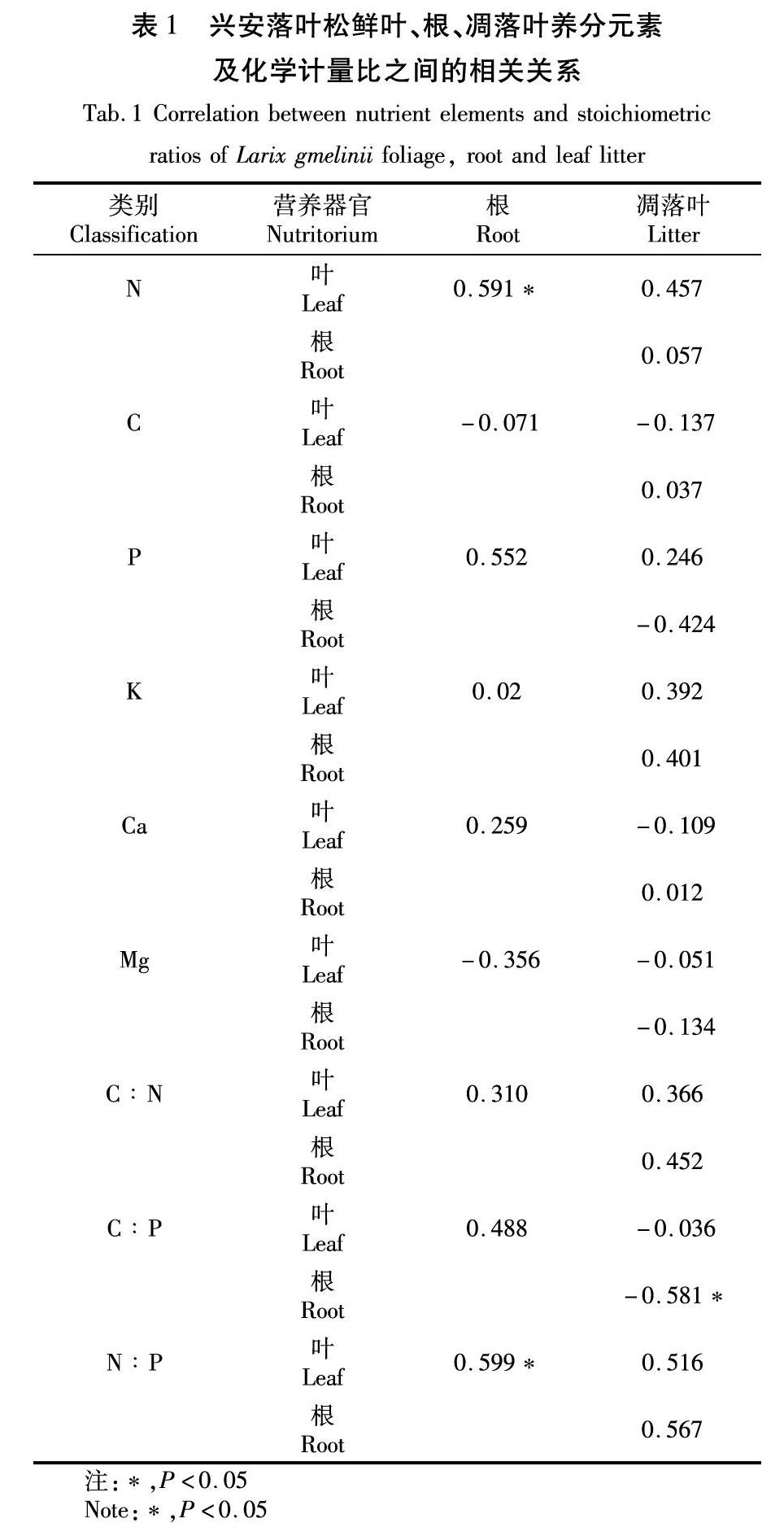
兴安落叶松鲜叶、根、凋落叶C、N、P、K、Ca、Mg含量相关性多数未达到显著性水平。鲜叶和根N具有显著正相关性( P <0.05)。鲜叶、根、凋落叶C∶N化学计量比的相关性均未达到显著水平。凋落叶和根C∶P表现为显著负相关性( P <0.05)。鲜叶和根N∶P表现为显著的正相关性( P <0.05),见表1。
2.3不同施N水平对兴安落叶松养分重吸收率的影响
鲜叶和凋落叶中各元素含量为N>Ca>P>K>Mg。鲜叶N、P、K、Mg含量显著高于凋落叶( P <0.05),鲜叶Ca含量显著低于凋落叶( P <0.05),表现出施N条件下叶片对N、P、K、Mg的重吸收能力加强(图1)。叶片N、P、K含量与凋落叶呈正相关关系,Ca、Mg含量为负相关关系。总体上,N、P、K、Mg重吸收效率分别为29%、30%、16%、8%,而Ca为-55%重吸收效率为负值。MN处理提高了兴安落叶松叶片对N的重吸收效率。LN处理加剧了Ca的富集。研究發现施N并未对兴安落叶松养分重吸收率产生显著影响,如图3所示。
N重吸收率和鲜叶N含量、N∶P之间的相关性不显著,如图4(a)(b)所示,P重吸收率与鲜叶P含量呈显著正相关关系( P <0.05),与鲜叶N∶P呈显著负相关关系( P <0.05),如图4(c)(d)所示。兴安落叶松鲜叶的N重吸收率与P重吸收率无显著的相关关系,如图5所示。
2.4兴安落叶松养分含量与化学计量比的影响因子
通过施N和营养器官叶和根对兴安落叶松C、N、P含量及其化学计量关系影响的一般(GLM)分析发现:施N对兴安落叶松N含量影响极显著( P <0.01),对P含量的影响未达到显著水平。相应的C∶P受施N影响不显著,但C∶N和N∶P受施N的影响极显著( P <0.01)。营养器官叶和根对兴安落叶松各元素含量,以及化学计量比均有极显著影响( P <0.01)。施N与叶和根的交互作用对N含量及C∶N有极显著的影响( P <0.01)(表2)。
3讨论
3.1不同施N水平对兴安落叶松鲜叶、凋落叶、根养分含量及相关性的影响
研究发现,在施N条件下植物C的变异系数最 小,N较大。原因可能是C是构成植物体干物质的主要成分,作为结构性物质,其受环境影响较小从而含量相对稳定变化较小。而N和P则参与植物多个生理活动,包括植物光合作用和细胞分裂等 [24]。施N条件下叶N增加能够提升叶片内部光合作用酶的数量, 从而提高叶片光合速率。此外,N∶P可以反映植物生长受N或P的限制情况 [26-27],植物常通过对N、P的吸收变化来保持相对稳定的N∶P。所以N含量变化较大可能是在施N条件下植物通过N的含量的变化调节植物体化学计量平衡,这是一种应对环境变化的策略或是植物通过N的变化来指示植物受限制情况。
N、 P在营养器官叶和根中的分配存在显著差异(图1),表现为叶高于根。植物不同器官营养元素的分布,除受土壤养分供应影响外,还与相应器官的生理功能密切相关 [28]。根系是吸收营养物质的主要营养器官,将植物连接到土壤,对吸收水分和矿物质养分交换有重要作用,通常需要较高的N、P来支持新陈代谢,还要将过量N、P转移到叶,此外由于采样时处于植物生长季,需要保持高的光合速率,此时作为光合器官的叶就需要保持高的N含量以保证植物光合的正常进行。本研究中施N整体上使鲜叶和根N含量显著提高,反应了施N促进兴安落叶松对N的吸收,这与以往的研究结果一致 [29-31]。植物叶片的矿质元素含量取决于土壤养分的可利用性。因此在养分充足的情况下,植物更倾向于利用土壤养分。以往的研究也显示,植物组织内的矿质元素含量与土壤中相应元素含量呈现显著正相关关系 [32]。此外,功能群和物种也是影响叶片N、P化学计量特征及其对N添加响应的重要因素。而且在施N情况下,植物对N、P等矿质元素的利用策略因需求、竞争力和优势度的不同也存在巨大差异 [33-34]。这些是笔者在试验中没有考虑到的问题。
近年来,越来越多的研究尝试从植物功能性状或资源经济谱的角度来探讨不同器官之间的性状关联, 相关研究不仅有助于理解植物各性状之间相互作用的机制 [35]、植物生长过程中对资源的利用和分配 [28],而且对进一步预测植物对环境变化的响应具有重要意义。研究表明鲜叶和根的N含量、P含量均呈显著正相关 [28,36]。本研究发现兴安落叶松鲜叶和根N含量有显著相关性,N∶P具有显著的正相关关系,这与之前的研究发现一致。根和凋落叶C∶P呈显著的负相关关系,未发现P元素在根和鲜叶之间的显著相关性。原因可能是小尺度范围内植物生长环境差异小,种内个体功能性状因变异幅度较小而关联不显著 [37]。还有一种可能是,采集的根样本组分复杂,其形态、寿命、生理差异巨大,是不同个体组成的高度异质的混合体 [38-39],从而影响了根与鲜叶之间相应元素的相关关系。另外,本研究在个体水平上的样本量有限,结果可能存在一定的随机性。植物鲜叶与细根性状具体的关联程度和机制还有待于进一步研究。
3.2不同施N水平对兴安落叶松鲜叶C∶N、C∶P、N∶P的影响
植物体的C∶N和C∶P能显示植物吸收营养所同化的C的能力,也可以反应植物对N和P的利用效率,一定程度也显示了土壤养分的供应情况,因而有重要的生态学意义。正常情况下C不是限制性元素,因此影响C∶N和C∶P的主要因素是N和P含量的变化。本研究发现,鲜叶和根的C∶N在MN和HN处理下有所下降,表明施N并未提高其中N利用效率。原因可能是在生态系统N含量趋于饱和状态时,N的持续添加超出了植物和微生物的可用限度时会减少生产力,所以随着N输入的增加植物生物量下降,尤其是在HN处理下。因此适度的施N在短期内有利于兴安落叶松生长,N过量处理会超出植物承受的阈值,从而造成植物生理进程紊乱。与对照相比鲜叶在MN处理和HN处理下C∶P高。表明施N增加了兴安落叶松对P的利用效率,指示P的短缺。
叶片是植物进行光合作用的主要器官,所以叶片N∶P的值常用来指示生态系统生产力受限制元素,也用来判断环境对植物养分的供应情况。通过对不同植物的施肥实验,Koerselman W [26]认为当N∶P>16表示生态系统受P限制,N∶P<14表示生态系统受N限制,N∶P在14~16時,表示生态系统受N和P的共同限制或者同时不受二者限制。本试验中,不同施N下兴安落叶松叶片N∶P变化范围为12~20,MN和HN处理下的N∶P明显高于Han等 [40]研究我国753个植物种得出的N∶P平均值(14.40)和世界上植物的平均值(13.80)。而对照组得出的N∶P介于16和14之间,受N、P共同限制,随N浓度增加,使该地受P限制加剧。原因可能是随着N的添加叶片P含量显著下降,或许是因为可利用性N增加,叶片N含量激增,需要吸收更多P才能达到养分平衡,但是当N负荷过重,即使是增加P的吸收机制,从长远来看亦不能阻止N∶P的增加 [41]。
3.3不同施N水平对兴安落叶松养分重吸收率的影响
植物通常会增加叶片养分浓度,但在营养丰富的环境中会减少对养分的吸收 [42]。这可能是为了回应实验中添加的营养物质 [43]。土壤养分有效性对植物养分重吸收效率有无影响一直是生态学界争论的焦点之一。Chapin F S、Arco J M D等 [44-45]对黑松( Pinus thunbergii )的研究发现,黑松叶片养分重吸收随土壤N有效性的降低而提高 [46]。另一些研究发现,叶片养分再吸收效率不受土壤养分有效性影响。Yowhan S [47]的研究表明,施肥(N和P)使日本落叶松N再吸收效率提高,而刚松( Pinus rigida )的N再吸收效率下降 [48]。基于这些发现,笔者预计N吸收策略将会被用于响应增加的N的可用性(从富N土壤中开发N的成本相对较低)而且重吸收策略也不会改变。研究结果表明N添加并未对兴安落叶松N重吸收率产生显著影响,但总体上呈下降趋势(图3),可以预料到,当环境养分供应超过植物对养分的需求时,植物将在最大程度上降低成本 [9]。因此,N吸收量的减少和凋落叶N的增加可能是N对植物生长的限制减弱和植物适应营养丰富环境的一种行为,并支持了养分重吸收率与土壤养分有效性有关这一观点。
如预期的那样,施N减少了凋落叶P含量,这表明采用了P重吸收策略,也表明N对P的重吸收有严格的控制作用 [43,49]。化学计量学稳态调节是植物适应环境变化的一种生态策略 [50]。根据化学计量理论,一种营养物质的有效性的变化必然会影响到其他营养物质的相对可用性,从而影响这些营养物质的获取和保存。笔者观察到,施N增加了叶片中的N∶P比值(>16),表明P限制加重。因此,为了保证其健康和化学计量的动态平衡,植物必须调整和优化P的获取和保存以适应N利用率的变化。另外,P的重吸收率和叶片P含量呈显著的正相关关系。Chapin F S [51]认为就叶片养分元素及其转移的速率来看,叶片本身N含量越高会导致叶片衰老过程中可转移的N的占比越高,即N的转移速率越高 [51]。但是这种规律对P却不适用,这可能是缺P植物具有将P转移到韧皮部来缓解P含量的匮乏的巨大潜能,这有利于植物对营养物质的转移。而P的重吸收率与N∶P呈现负相关关系,这可能是N∶P越大,P限制越严重,植物需要通过降低P的重吸收率来对生活环境的状况进行反馈,通过凋落叶归还到土壤,来缓解N诱导的P限制。
本研究显示除了N和P重吸收之外,叶片对K重吸收率为16%,Mg重吸收率为8%低于全球范围K(70.1%)和Mg(28.6%) [52]。Ca呈现富集状态,重吸收率为-55%这与全球范围Ca重吸收率(10.9%)不一致,但与喀斯特地区Ca重吸收率(-44%)一致 [53]。原因是Ca作为植物的一种结构元素,尤其是细胞壁中的Ca,在衰老过程中比其他元素吸收更少且Ca素不易流动便富集在凋落叶中 [54]。
总体上,实验表明N添加增加了凋落叶N的养分含量,降低了叶片养分重吸收的能力,降低了凋落叶P的含量,提高了P的重吸收能力,但对N、P的重吸收效率影响不显著。这一结果与以前的研究结果一致,即养分的添加对重吸收有很强的影响。但对重吸收效率没有明显影响 [46,55]。因此,笔者的发现支持这样的论点,即选择取决于衰老叶片中的残余养分浓度,而不是比例吸收本身 [9,23]。另外Kobe R K [56]认为研究结果迥异的原因大概是由于被用来实验物种间的差异、生长环境的差异以及在实验处理方法上的差异所导致的。比如,自然条件的光照、积温、降水和病虫害等的参与都能影响养分重吸收。还有就是,叶片在衰老过程中质量分数下降,叶面积缩水下降,所以用质量和叶面积来表征养分重吸收率存在误差。由此可见,对于植物养分重吸收的方法和理论均有待完善。
3.4兴安落叶松养分含量与化学计量比的影响因子
通过施N和各营养器官叶和根对兴安落叶松C、N、P含量及其化学计量关系影响的一般线性模型(GLM)分析(表2),结果表明营养器官的叶和根是影响,兴安落叶松C、N、P及化学计量比的主导因子。而N添加对兴安落叶松各元素及化学计量比无显著影响。
化学计量内稳性(Stoichiometric homoeostasis)是生态化学计量学存在的基础理论。化学计量内稳性是一种调节机制,它的工作原理是调节生物对环境因子的响应,从而维持生态系统结构、功能的稳定性 [45]。经研究发现C、N、P化学计量比中N∶P随着施N的增加除HN外均影响不显著,保持了相对稳定性。这一结果表明N沉降条件下兴安落叶松仍能形成稳定群落并处于优势地位是该地区的优势种。内稳性是其中重要的调节机制,但这一机制的作用机理有待于进一步研究。可以对植物养分与土壤养分,特别是速效养分之间关系进行动态监测以阐明其作用原理。植物养分含量及化学计量关系的影响因素众多,其中包括植物的进化史、生活史、遗传和不同器官的生理功能等,同时与土壤营养元素的有效供给及其在植物体内的运输、储存和利用等均有联系 [57-58]。本研究仅对植物地上器官叶,地下根做了比较,对植物其他的的生理功能器官间的相关性未进行探索,这还有待于实验进一步研究。
4结论
大兴安岭兴安落叶松营养器官叶和根各养分含量存在显著差异,表明叶和根是兴安落叶松养分含量变化的主导因子。在养分含量充足的情况下植物更倾向于利用转化成本低的养分,N添加N、P的重吸收能力有显著影响,并且N添加對植物体N、P状况的影响是不对称的,对N∶P的影响大于对个体中N、P含量的影响,使得N∶P增大,加剧了P的限制。
总之,植物在寻求化学计量平衡时,可以调节外部养分获取(根捕获)和内部养分保存(叶片重吸收)。植物养分经济中对根系捕获和叶片吸收的分配依赖于养分限制的强度和类型。考虑到元素的化学计量耦合,研究表明,N负荷的增加不仅对N的获取和保护有着重要的影响,而且对其它基本元素(如P)也有重要的影响。
【参考文献】
[1]LIU X J, ZHANG Y, HAN W X, et al. Enhanced nitrogen deposition over China[J]. Nature, 2013, 494(7438):459-462.
[2]GALLOWAY J N, DENTENER F J, CAPONE D G, et al. Nitrogen cycles: past, present, and future[J]. Biogeochemistry, 2004, 70(2):153-226.
[3]GALLOWAY J N, TOWNSEND A R, ERISMAN J W, et al. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions[J]. Science, 2008, 320(5878):889-892.
[4]STERNER R W, ELSER J J. Ecological stoichiometry: the biology of elements from molecules to the biosphere[M]. Princeton: Princeton University Press, 2002.
[5]ELSER J J, FAGAN W F, KERKHOFF A J, et al. Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change[J]. New Phytologist, 2010, 186(3):593-608.
[6]REICH P B, WRIGHT I J, CAVENDERBARES J, et al. The evolution of plant functional variation: traits, spectra, and strategies[J]. International Journal of Plant Sciences, 2003, 164(3):S143-S164.
[7]AERTS R, CHAPIN F S. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns[J]. Advances in Ecological Research, 1999, 30(8):1-67.
[8]AERTS R. Nutrient resorption from senescing leaves of perennials: are there general patterns?[J]. Journal of Ecology, 1996, 84(4):597-608.
[9]WRIGHT I J, WESTOBY M. Nutrient concentration, resorption and lifespan: leaf traits of Australian sclerophyll species[J]. Functional Ecology, 2003, 17(1):10-19.
[10]WANG M, MURPHY M T, MOORE T R. Nutrient resorption of two evergreen shrubs in response to long-term fertilization in a bog[J]. Oecologia, 2014, 174(2):365-377.
[11]HODGE A. The plastic plant: root responses to heterogeneous supplies of nutrients[J]. New Phytologist, 2004, 162(1):9-24.
[12]OLANDER L P, VITOUSEK P M. Regulation of soil phosphatase and chitinase activity by N and P availability[J]. Biogeochemistry, 2000, 49(2):175-190.
[13]BRAUN S, THOMAS V F, QUIRING R, et al. Does nitrogen deposition increase forest production? The role of phosphorus[J]. Environmental Pollution, 2010, 158(6):2043-2052.
[14]CAIRNEY J W G. Ectomycorrhizal fungi: the symbiotic route to the root for phosphorus in forest soils[J]. Plant & Soil, 2011, 344(1/2):51-71.
[15]ALDOUS A R. Nitrogen translocation in Sphagnum mosses: effects of atmospheric nitrogen deposition[J]. New Phytologist, 2002, 156(2):241-253.
[16]OSTONEN I, HELMISAARI H S, BORKEN W, et al. Fine root foraging strategies in Norway spruce forests across a European climate gradient[J]. Global Change Biology, 2011, 17(12):3620-3632.
[17]YUAN Z Y, CHEN H Y H. Negative effects of fertilization on plant nutrient resorption[J]. Ecology, 2016, 96(2):373-380.
[18]SEE C R, YANAI R D, FISK M C, et al. Soil nitrogen affects phosphorus recycling: foliar resorption and plant-soil feedbacks in a northern hardwood forest[J]. Ecology, 2015, 96(9):2488-2498.
[19]MARSCHNER H. Mineral nutrition of higher plants[M]. Salt Lake City: Academic Press, 2011.
[20]賀金生, 韩兴国. 生态化学计量学:探索从个体到生态系统的统一化理论 [J]. 植物生态学报, 2010, 34(1):2-6.
HE J S, HAN X G. Ecological stoichiometry: searching for unifying principles from individuals to ecosystems[J]. Chinese Journal of Plant Ecology, 2010, 34(1):2-6.
[21]REICH P B. The world‐wide 'fast-slow' plant economics spectrum: a traits manifesto[J]. Journal of Ecology, 2014, 102(2):275-301.
[22]PRIMICIA I, IMBERT J B, TRAVER M C, et al. Inter-specific competition and management modify the morphology, nutrient content and resorption in Scots pine needles[J]. European Journal of Forest Research, 2014, 133(1):141-151.
[23]KILLINGBECK K T. Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency[J]. Ecology, 1996, 77(6):1716-1727.
[24] NIKLAS K J, OWENS T, REICH P B, et al. Nitrogen/phosphorus leaf stoichiometry and the scaling of plant growth[J]. Ecology Letters, 2010, 8(6):636-642.
[25]HESSEN D O, ?GREN G I, ANDERSON T R, et al. Carbon sequestration in ecosystems: the role of stoichiometry[J]. Ecology, 2004, 85(5):1179-1192.
[26]KOERSELMAN W. The vegetation N: P ratio: a new tool to detect the nature of nutrient limitation[J]. Journal of Applied Ecology, 1996, 33(6):1441-1450.
[27]ELSER J J, FAGAN W F, DENNO R F, et al. Nutritional constraints in terrestrial and freshwater food webs[J]. Nature, 2000, 408(6812):578-580.
[28]KERKHOFF A J, FAGAN W F, ELSER J J, et al. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants[J]. The American Naturalist, 2006, 168(4):E103-22.
[29]MAGILL A H, ABER J D, BERNTSON G M, et al. Long-term nitrogen additions and nitrogen saturation in two temperate forests[J]. Ecosystems, 2000, 3(3):238-253.
[30] MCNULTY S G, BOGGS J, ABER J D, et al. Red spruce ecosystem level changes following 14 years of chronic N fertilization[J]. Forest Ecology & Management, 2005, 219(2):279-291.
[31]ROWE E C, SMART S M, KENNEDY V H, et al. Nitrogen deposition increases the acquisition of phosphorus and potassium by heather Calluna vulgaris[J]. Environmental Pollution, 2008, 155(2):201-207.
[32]REICH P B, OLEKSYN J. Global patterns of plant leaf N and P in relation to temperature and latitude[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30):11001-11006.
[33]TILMAN D. Constraints and tradeoffs: toward a predictive theory of competition and succession[J]. Oikos, 1990, 58(1):3-15.
[34]G SEWELL S, KOERSELMAN W. Variation in nitrogen and phosphorus concentrations of wetland plants[J]. Perspectives in Plant Ecology Evolution & Systematics, 2002, 5(1):37-61.
[35]EVINER V T, III F S C. Functional matrix: a conceptual framework for predicting multiple plant effects on ecosystem processes[J]. Annual Review of Ecology Evolution & Systematics, 2003, 34(34):455-485.
[36]GENG Y, WANG L, JIN D, et al. Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits[J]. Oecologia, 2014, 175(2):445-455.
[37]徐冰, 程雨曦, 甘慧洁,等.内蒙古锡林河流域典型草原植物叶片与细根性状在种间及种内水平上的关联 [J. 植物生态学报, 2010, 34(1):29-38.
XU B, CHENG Y X, GAN H J, et al. Correlations between leaf and fine root traits among and within species of typical temperate grassland in Xilin River Basin, Inner Mongolia[J]. Chinese Journal of Plant Ecology, 2010, 34(1):29-38.
[38]PREGITZER K S, DEFOREST J L, BURTON A J, et al. Fine root architecture of nine North American trees[J]. Ecological Monographs, 2002, 72(2):293-309.
[39]GUO D, LI H, MITCHELL R J, et al. Fine root heterogeneity by branch order: exploring the discrepancy in root turnover estimates between minirhizotron and carbon isotopic methods[J]. New Phytologist, 2008, 177(2):443-456.
[40]HAN W, FANG J, GUO D, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2010, 168(2):377-385.
[41]HEERWAARDEN L M V, TOET S, AERTS R. Current measures of nutrient resorption efficiency lead to a substantial underestimation of real resorption efficiency: facts and solutions[J]. Oikos, 2010, 101(3):664-669.
[42]HAYES P, TURNER B L, LAMBERS H, et al. Foliar nutrient concentrations and resorption efficiency in plants of contrasting nutrient acquisition strategies along a 2 million year dune chronosequence[J]. Journal of Ecology, 2014, 102(2):396-410.
[43]MAO R, ZENG D H, ZHANG X H, et al. Responses of plant nutrient resorption to phosphorus addition in freshwater marsh of Northeast China[J]. Scientific Reports, 2015, 5(5):8097.
[44]ENOKI T, KAWAGUCHI H. Nitrogen resorption from needles of Pinus thunbergii, Parl. growing along a topographic gradient of soil nutrient availability[J]. Ecological Research, 2010, 14(1):1-8.
[45]CHAPIN F S, MOILANEN L. Nutritional controls over nitrogen and phosphorus resosrption from Alaskan birch leaves[J]. Ecology, 1991, 72(2):709-715.
[46]LI X, LIU J, FAN J, et al. Combined effects of nitrogen addition and litter manipulation on nutrient resorption of Leymus chinensis in a semi-arid grassland of northern China[J]. Plant Biology, 2015, 17(1):9-15.
[47]YOWHAN S, IMKYUN L, SOUNGRYOUL R. Nitrogen and phosphorus dynamics in foliage and twig of pitch pine and Japanese larch plantations in relation to fertilization[J]. Journal of Plant Nutrition, 2000, 23(5):697-710.
[48]趙琼, 刘兴宇, 胡亚林, 等. 氮添加对兴安落叶松养分分配和再吸收效率的影响[J]. 林业科学, 2010,46(05):14-19.
ZHAO Q, LIU X Y, HU Y L, et al. Effects of nitrogen addition on nutrient allocation and reuptake efficiency of Larix gmelini[J]. Forestry Science, 2010, 46(5):14-19.
[49]L X T, HAN X G. Nutrient resorption responses to water and nitrogen amendment in semi-arid grassland of Inner Mongolia, China[J]. Plant & Soil, 2010, 327(1/2):481-491.
[50]YU Q, ELSER J J, HE N, et al. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland[J]. Oecologia, 2011, 166(1):1-10.
[51]CHAPIN F S, KEDROWSKI R A. Seasonal changes in nitrogen and phosphorus fractions and autumn retranslocation in evergreen and deciduous taiga trees[J]. Ecology, 1983, 64:376-391.
[52]VERGUTZ L, MANZONI S, PORPORATO A, et al. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants[J]. Ecological Monographs, 2012, 82(2):205-220.
[53]LIU C, LIU Y, GUO K, et al. Concentrations and resorption patterns of 13 nutrients in different plant functional types in the karst region of south-western China[J]. Annals of Botany, 2014, 113(5):873-885.
[54]KAZAKOU E, GARNIER E, NAVAS M, et al. Components of nutrient residence time and the leaf economics spectrum in species from Mediterranean old-fields differing in successional status[J]. Functional Ecology, 2010, 21(2):235-245.
[55]KOZOVITS A R, BUSTAMANTE M M C, GAROFALO C R, et al. Nutrient resorption and patterns of litter production and decomposition in a Neotropical Savanna[J]. Functional Ecology, 2007, 21(6):1034-1043.
[56]KOBE R K, LEPCZYK C A, IYER M. Resorption efficiency decreases with increasing green leaf nutrients in a global date set[J]. Ecology, 2005, 86(10):2780-2792.
[57]周紅艳, 吴琴, 陈明月,等. 鄱阳湖沙山单叶蔓荆不同器官碳、氮、磷化学计量特征 [J. 植物生态学报, 2017, 41(4):461-470.
ZHOU H Y, WU Q, CHEN M Y. Stoichiometric characteristics of carbon, nitrogen and phosphorus in different organs of Vitex mongolicum in Sandy Mountain, Poyang Lake[J]. Journal of Plant Ecology, 2017, 41(4): 461-470.
[58]刘芳,葛江丽,施汉钰,等.西洋参地氮磷钾含量变化规律研究[J].林业科技,2017,42(6):19-21. LIU F,GE J L,SHI H Y, et al. Study on changing rule of nitrogen, phosphorus and potassium content in Panax quinquefolium field[J]. Forestry Science & Technology,2017,42(6):19-21.
