
饲粮中添加甘露寡糖对肉仔鸡生长性能及组织天然免疫相关基因表达的影响
2019-10-21 06:19熊阿玲包龙飞许兰娇龙光莲李君惠黎观红
中国粮油学报 2019年9期
熊阿玲 包龙飞 许兰娇 万 根 龙光莲 李君惠 黎观红
(江西农业大学江西省动物营养重点实验室;江西省营养饲料开发工程研究中心,南昌 330045)
Toll样受体(Toll-like receptors, TLRs)是表达于动物免疫细胞和上皮细胞的一类重要模式识别受体(pattern recognition receptors, PRRs),是机体最初感知微生物存在的天然免疫识别受体,是启动哺乳动物和家禽肠道天然免疫应答的关键PRR,而且还作为天然免疫和获得性免疫的桥梁而调节获得性免疫应答[1-2]。肠道上皮细胞和免疫细胞可通过 TLRs区分病原微生物和共生有益菌产生不同免疫效应。TLRs能够识别微生物相关分子模式(microbe-associated molecular patterns, MAMPs)并激活下游信号通路从而释放出一系列免疫效应分子并产生一系列生理和免疫效应,释放的免疫效应分子主要包括抗菌肽(β-防御素和cathelicidins)、细胞因子和趋化因子等[3-4]。基于TLRs在天然免疫应答中的重要作用,近年来,鸡TLRs(chTLRs, chicken TLRs)的研究受到极大关注。在目前已鉴定的10种禽类chTLRs中,TLR2主要识别来自于G+细菌的细胞壁组分如肽聚糖、脂蛋白和脂磷壁酸,TLR4识别G-细菌的LPS[5]。
抗菌肽(antimicrobial peptides, AMPs)在宿主免疫调控中起到至关重要的作用,是鸡抗感染天然免疫至关重要的效应分子[6-8]。β-防御素和cathelicidins是鸡体内的两个主要抗菌肽家族。到目前为止,从鸡组织中鉴定出14种禽β-防御素(avian β-defensins, AvBDs),即AvBD1-14[8-9]。在鸡组织中发现的cathelicidins(Cath)有4 种,即Cath-1、Cath-2, Cath-3和Cath-B1[10]。鸡体内表达的抗菌肽家族中,AvBD9和Cath-B1是广泛表达于鸡组织尤其是消化道和免疫组织器官的一种重要抗菌肽,AvBD9和Cath-B1除具有很强的广谱抗菌活性外,在体内还发挥着重要的免疫调节功能,在鸡的抗感染免疫中发挥重要作用[6-7, 11-12]。
国内外学者对甘露寡糖(mannan-oligosaccharides, MOS)在畜禽生产中的应用进行了大量研究。大多数研究表明,肉鸡饲粮中添加MOS可提高日增重、改善饲料利用率、降低死亡率,促进肠道发育,改善肠道微生物区系,提高肉鸡肠道免疫和系统免疫功能[13-15]。但MOS调节机体免疫功能的作用机制目前还不是十分清楚。MOS是否可通过影响鸡天然免疫相关基因如TLRs和抗菌肽表达而发挥其免疫调节作用?这方面的研究鲜见报道。本研究旨在探讨饲粮中添加MOS对肉仔鸡生长性能以及重要的天然免疫相关基因TLR2、TLR4、AvBD9和Cath-B1基因表达的影响,从天然免疫角度初步探讨MOS的免疫调节作用,为MOS作为抗生素替代品的推广应用提供参考。
1 材料与方法
1.1 试验设计
采用单因子试验设计,选取体重相近的1日龄雄性白羽肉仔鸡(科宝500)176只,随机分成4个处理组,每组4个重复,每个重复11只鸡,试验期为42 d。4个处理组分别饲喂在基础饲粮中添加0(对照组)、0.3、0.6、0.9 g/kg MOS的饲粮。各组饲粮不添加任何抗生素,本试验所用的甘露寡糖来源于食品啤酒酵母细胞壁,纯度≥95%。
1.2 试验饲粮组成及营养水平
本试验采用的基础饲粮为玉米—豆粕型饲粮,按照1~21、22~42日龄龄两个阶段,根据NRC(1994)和我国《鸡的饲养标准》(2004)推荐的营养水平配制。基础饲粮组成及其营养水平见表1。
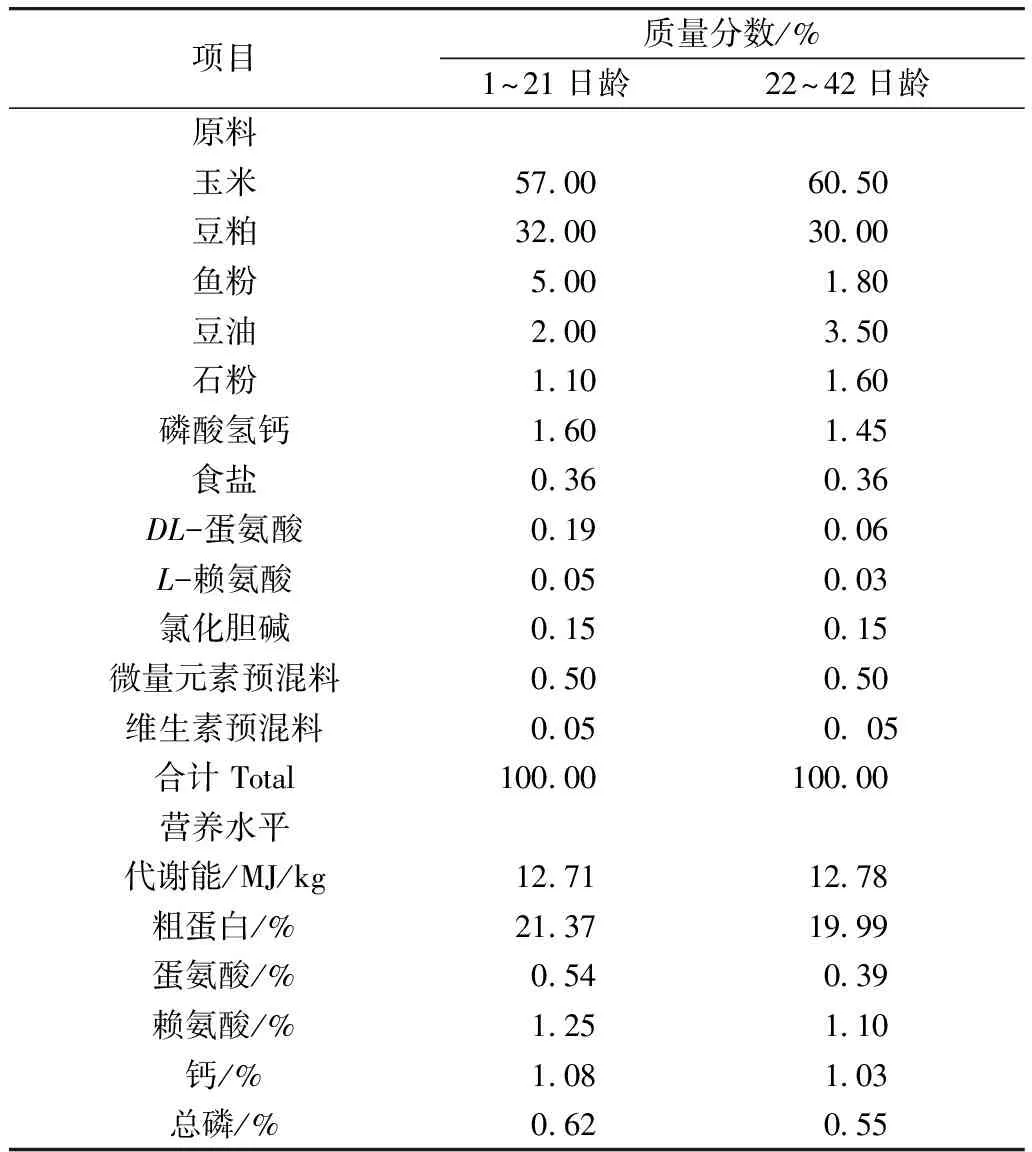
表1 基础饲粮组成及营养水平
注: 微量元素预混料为每千克饲粮提供:铁(FeSO4·H2O)80 mg,铜(CuSO4·5H2O)8 mg,锌(ZnSO4·H2O)60 mg,锰(MnSO4·H2O)80 mg,碘(KI)0.35 mg,硒(Na2SeO3)0.15 mg。维生素预混料为每千克饲粮提供:维生素A 8 000 IU,维生素D33 000 IU,维生素E 10 IU,维生素K32 mg,维生素B11.5 mg,维生素B28.0 mg,维生素B62.5 mg,维生素B120.011 mg,叶酸0.9 mg,烟酸44 mg,D-泛酸钙11 mg,生物素0.11 mg。营养水平为计算值。
1.3 饲养管理
试验鸡采用三层笼笼养。试验前对鸡舍进行充分冲洗和严格消毒,入雏前24 h将鸡舍升温至32~35 ℃,此后温度每周降低2~3 ℃,直至保持在22~24 ℃为止。采用连续光照、自然通风。试验期舍内光照、湿度和温度根据常规饲养管理要求进行控制,鸡只按正常免疫程序进行免疫。整个试验期鸡只自由采食和饮水。
1.4 组织样品的采集与制备
分别在肉仔鸡21和42日龄08 ∶00喂料前从每个重复中随机选取两只体重相近的鸡空腹称重,称重后采用颈动脉放血法将肉仔鸡致死,冰浴上迅速分离法氏囊、胸腺、脾脏、肝脏、回肠和盲肠,用冰冷的生理盐水冲洗干净并用滤纸吸干表面水分,用天平称重法氏囊、胸腺和脾脏,用于计算器官指数。液氮速冻肝脏、脾脏、回肠和盲肠,转入-80 ℃低温冰箱保存备用,用于基因表达量的测定。
1.5 测定指标与方法
1.5.1 生长性能
分别于肉仔鸡1、21和42 日龄对每重复肉仔鸡进行空腹称重,并在试验期间记录饲料耗料量,计算肉仔鸡各阶段的平均日增重(ADG)、平均日采食量(ADFI)和料重比(F/G)。
1.5.2 免疫器官指数
根据所屠宰试验鸡体重与免疫器官(法氏囊、胸腺、脾脏)质量计算各免疫器官指数。计算公式:免疫器官指数(g/kg)=免疫器官重(g)/鸡活重(kg)。
1.5.3 组织TLR2、TLR4、AvBD9和Cath-B1基因mRNA表达量
采用Trizol法提取肉仔鸡肝脏、脾脏、回肠和盲肠组织总RNA,用超微量紫外分光光度仪测定其溶度和纯度,用1%琼脂糖凝胶电泳检测分析其完整性。对质量合格的RNA进行反转录,用于后续试验。采用反转录试剂盒将提取的总RNA反转录成cDNA。根据GenBank数据库中提供的鸡GADPH、TLR2、TLR4、AvBD9和Cath-B1基因序列,用Primer Express 3.0软件设计其引物,引物由ShineGene公司(中国上海)合成,特异性引物及探针的序列见表2。根据试剂盒说明书优化反应体系,对每个样品 cDNA进行目的基因和内参基因荧光定量PCR反应,扩增反应在FTC2000荧光定量PCR系统(Funglyn Biotech, CA)中进行。反应总体系为50 μL:25 μL 2 × Hotstart Fluo-PCR mix, 2 × 1 μL Primers (25 pmol/μL),0.5 μL Probe(25 pmol/μL),1 μL cDNA,21.5 μL DEPC水。反应条件:94 ℃ 4 min,94 ℃ 20 s,60 ℃ 30 s循环40次。PCR扩增反应结束后,PCR仪自带的ABI7300 SDS Software分析软件分析所得结果Ct值。目的基因的mRNA相对表达量用2-△△Ct法分析,内参基因选用GAPDH。

表2 实时定量PCR扩增所使用的特异性引物序列、GenBank登入号及产物大小
1.6 数据处理与统计分析
采用SPSS 17.0软件的One-way ANOVA过程进行单因素方差分析,采用Duncan氏法进行多重比较。结果以“平均值±标准差”表示,P<0.01表示差异极显著,P<0.05表示差异显著,P>0.05表示差异不显著。
2 结果
2.1 甘露寡糖对肉仔鸡生长性能的影响
由表3可知,饲粮中添加MOS对肉仔鸡生长前期(1~21日龄)平均日采食量无显著影响(P>0.05),但显著提高肉仔鸡生长前期平均日增重和显著降低料重比(P<0.05或P<0.01),MOS不同添加水平组之间平均日增重和料重比无显著差异(P>0.05)。与对照组相比,饲粮中添加0.6 g/kg MOS显著提高肉仔鸡生长后期(22~42日龄)平均日采食量(P<0.01)、平均日增重(P<0.05)和料重比(P<0.01),MOS不同添加水平组之间日增重无显著差异。添加0.3~0.9 g/kg MOS显著提高肉仔鸡生长全期(1~42日龄)的平均日采食量和平均日增重(P<0.05或P<0.01)。

表3 饲粮中添加甘露寡糖对肉仔鸡生长性能的影响
注:在同一日龄,同列数据肩标字母相同或无肩标字母表示差异不显著(P>0.05),肩标小写字母不同表示差异显著(P<0.05),大写字母不同表示差异极显著(P<0.01)。余同。
2.2 甘露寡糖对肉仔鸡免疫器官指数的影响
由表4可知,饲粮中添加0.6 g/kg和0.9 g/kg MOS显著提高21日龄肉仔鸡的法氏囊指数(P<0.05),饲粮中添加0.9 g/kg MOS显著提高42日龄胸腺指数(P<0.05),但MOS不同添加水平组之间差异不显著(P>0.05)。添加不同水平MOS处理组对21日龄肉仔鸡胸腺指数、脾脏指数和42日龄的法氏囊指数、脾脏指数均无显著影响(P>0.05)。
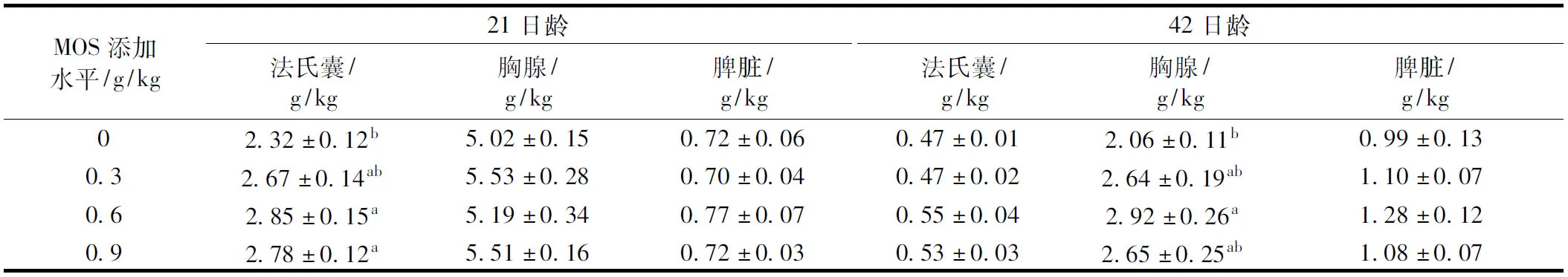
表4 甘露寡糖对肉仔鸡免疫器官指数的影响
2.3 甘露寡糖对肉仔鸡组织TLR2 mRNA表达的影响
由表5可知,饲粮添加MOS对21日龄肉仔鸡肝脏、脾脏、回肠和盲肠以及42日龄肝脏和脾脏TLR2 mRNA表达无显著影响(P>0.05),但显著影响42日龄肉仔鸡回肠和盲肠TLR2 mRNA表达(P<0.05或P<0.01)。与对照组相比,饲粮添加0.3和0.9 g/kg MOS分别极显著提高42日龄肉仔鸡回肠和盲肠TLR2 mRNA表达(P<0.01)。由表5亦可知,TLR2基因在所测定的肝脏、脾脏、回肠和盲肠四个组织中的表达存在组织差异性,以盲肠TLR2 mRNA表达水平最高。
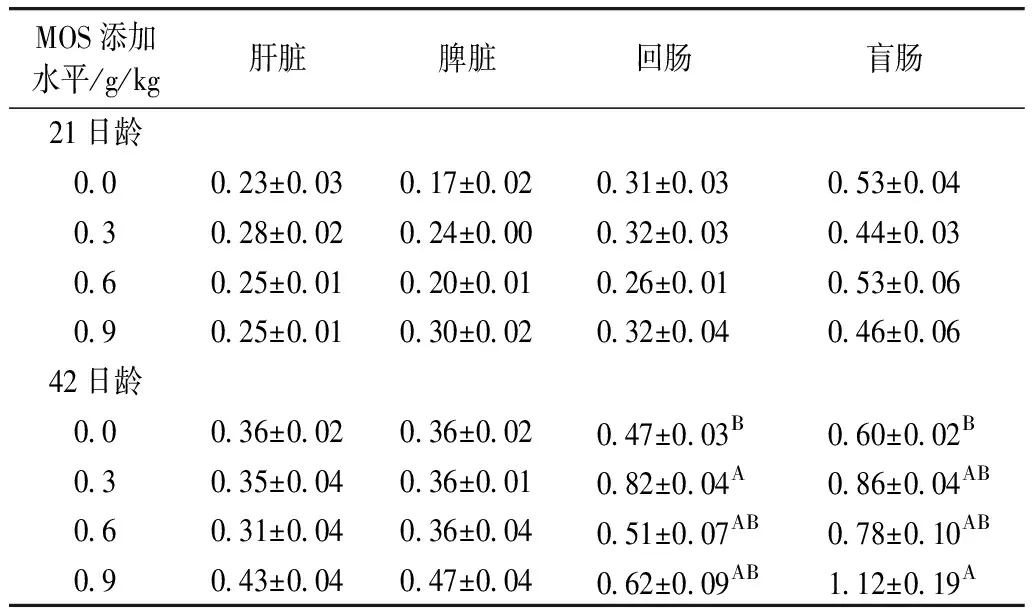
表5 饲粮中添加甘露寡糖对肉仔鸡组织TLR2 mRNA表达的影响
2.4 甘露寡糖对肉仔鸡组织TLR4 mRNA表达的影响
由表6可知,饲粮中添加MOS对21日龄肉仔鸡脾脏、盲肠和42日龄盲肠TLR4 mRNA表达无显著影响(P>0.05),但显著影响21日龄肝脏、回肠和42日龄肉鸡肝脏、脾脏、回肠TLR4 mRNA表达(P<0.05或P<0.01)。在21日龄,饲粮中添加0.6和0.9 g/kg MOS极显著提高肝脏TLR4 mRNA表达(P<0.01),但0.3 g/kg MOS添加组与对照组差异不显著(P>0.05);回肠TLR4 mRNA表达随MOS添加量的增加而呈上升趋势,0.9 g/kg MOS添加组显著高于对照组(P<0.05)。在42日龄,三个MOS添加组均与对照组差异不显著(P>0.05)。饲粮中添加0.6和0.9 g/kg MOS分别显著提高42日龄回肠(P<0.01)和脾脏(P<0.05)TLR4 mRNA表达。由表6亦可知,在所测定的肝脏、脾脏、回肠和盲肠的四个组织中,以盲肠TLR4 mRNA表达水平最高,而肝脏表达水平最低。

表6 饲粮中添加甘露寡糖对肉仔鸡组织TLR4 mRNA表达的影响
2.5 甘露寡糖对肉仔鸡组织AvBD9 mRNA表达的影响
由表7可知,饲粮中添加MOS显著影响21日龄肉仔鸡脾脏、回肠和盲肠以及42日龄回肠和盲肠AvBD9 mRNA表达(P<0.05或P<0.01)。在21日龄,与对照组相比,饲粮中添加0.3 g/kg MOS显著提高脾脏(P<0.05)和回肠(P<0.01)AvBD9 mRNA的表达;饲粮中添加0.6 g/kg MOS极显著提高回肠和盲肠AvBD9 mRNA表达(P<0.01),而饲粮中添加0.9 g/kg MOS显著提高盲肠AvBD9 mRNA表达(P<0.05)。在42日龄,与对照组相比,饲粮中添加0.9 g/kg MOS显著提高回肠AvBD9 mRNA的表达(P<0.05);饲粮中添加0.3~0.9 g/kg MOS极显著提高盲肠AvBD9 mRNA表达(P<0.01)。由表7亦

表7 饲粮中添加甘露寡糖对肉仔鸡组织AvBD9 mRNA表达的影响
可知,AvBD9 mRNA表达水平在所测定的肝脏、脾脏、回肠和盲肠四个组织中存在差异,肝脏AvBD9 mRNA表达水平最高,而回肠表达水平最低。
2.6 甘露寡糖对肉仔鸡组织Cath-B1 mRNA表达的影响
由表8可知,饲粮中添加MOS对21日龄肉仔鸡肝脏和42日龄回肠Cath-B1 mRNA表达无显著影响(P>0.05),但显著影响21日龄肉仔鸡脾脏、回肠和盲肠以及42日龄肝脏、脾脏和盲肠Cath-B1 mRNA表达(P<0.05或P<0.01)。在21日龄,与对照组相比,饲粮中添加0.3 g/kg MOS显著提高脾脏(P<0.05)以及极显著提高回肠和盲肠Cath-B1 mRNA表达(P<0.01);添加0.9 g/kgMOS极显著提高盲肠Cath-B1 mRNA表达(P<0.01),但显著降低脾脏Cath-B1 mRNA表达(P<0.05)。在42日龄,与对照组相比,饲粮中添加0.6 g/kg MOS极显著提高肝脏Cath-B1 mRNA表达(P<0.01),而饲粮中添加0.9 g/kg MOS显著提高肝脏和脾脏(P<0.05)以及极显著提高盲肠(P<0.01)Cath-B1 mRNA表达。由表8亦可知,Cath-B1 mRNA表达水平在所测定的肝脏、脾脏、回肠和盲肠四个组织中存在差异,Cath-B1 mRNA高表达于脾脏和盲肠,而在肝脏中表达水平最低。

表8 饲粮中添加甘露寡糖对肉仔鸡组织Cath-B1 mRNA表达的影响
3 讨论
国内外学者对饲粮添加MOS对畜禽生长性能的影响进行了大量的研究,但不同研究者所得结果不尽一致。Pelicano等[16]报道,饲粮添加MOS可显著提高1~21日龄肉仔鸡日增重和饲料转化率,MOS对肉仔鸡生长性能的改善效应并不能持续至42日龄。Hooge 等[17]、Kumprecht等[18]和武威等[19]试验结果均表明,MOS可显著提高肉仔鸡的生长性能。本研究发现饲粮中添加MOS能显著改善肉仔鸡生长性能,添加0.3~0.9 g/kg MOS显著提高肉仔鸡生长前期(1~21日龄)的平均日增重和降低料重比,且添加0.9 g/kg MOS的可取得最佳的料重比;添加0.6 g/kg MOS对提高肉仔鸡生长后期(22~42日龄)平均日采食量和平均日增重的效果最明显。本试验结果与上述报道的研究结果类似。然而,Biggs等[20]和Baurhoo等[21]均报道,饲粮添加MOS对生长前期肉鸡的生产性能没有改善作用。Yang等[22]研究报道,MOS对肉鸡生长后期(22~42日龄)的生产性能没有改善作用。相反,Sohail等[23]和Cheng等[24]分别发现,饲粮中添加MOS可显著改善热应激条件下肉鸡日增重、采食量和饲料转化率。有关MOS对肉鸡生产性能的改善作用,不同学者的研究结果不尽一致,其原因可能与MOS的来源、添加量、饲粮组成、鸡的生长阶段及饲养环境等因素有关。
禽类的免疫器官主要包括法氏囊、胸腺和脾脏,其中,法氏囊和胸腺为禽类的中枢免疫器官,脾脏为禽类的次级免疫器官。免疫器官指数的大小常用来反映或评价机体免疫功能状态[25]。本研究发现,饲粮中添加0.6 g/kg可分别提高21日龄肉仔鸡法氏囊指数和42日龄胸腺指数,由此说明,饲粮添加适量的MOS可提高肉鸡免疫功能。同样地, Sohail等[26]和Attia等[27]均报道,饲粮添加MOS可显著提高肉仔鸡法氏囊和胸腺指数,但对脾脏指数无影响,这与本研究结果相一致。
TLRs是启动哺乳动物和家禽天然免疫应答的关键模式识别受体,并可调节获得性免疫应答,在机体的抗感染免疫中发挥重要作用。研究表明,TLR2可识别酵母细胞壁甘露聚糖,介导宿主相关免疫效应分子的分泌和天然免疫应答的产生[28]。TLR4亦可识别葡萄糖醛酸木糖甘露聚糖(glucuronoxylomannan)、酿酒酵母(Saccharomycescerevisiae)和假丝酵母(Candidaalbicans)来源的甘露聚糖[29]。TLR2和TLR4表达的提高及其介导的下游信号通路的激活与胃肠道屏障功能的改善密切相关,由此提高肠道对病原菌入侵和对感染的抵抗力[30-31]。来源于酵母细胞壁MOS作为甘露聚糖大分子的一部分,亦可能作为MAMP而被TLRs识别而启动机体天然免疫应答。本研究发现,在本试验添加水平范围内(0.3~0.9 g/kg),饲粮添加MOS可不同程度地提高肉鸡回肠和盲肠中TLR2和TLR4 mRNA的表达。Yitbarek等[32]报道,产气荚膜梭菌(Clostridiumperfringens)感染对肉鸡回肠TLR2和TLR4以及盲肠扁桃体TLR4基因表达无影响,但饲粮中添加MOS显著提高感染产气荚膜梭菌肉鸡回肠TLR2和TLR4以及盲肠扁桃体TLR4基因表达,但感染产气荚膜梭菌的肉鸡其盲肠扁桃体TLR2基因表达显著升高,而饲粮中添加MOS对感染产气荚膜梭菌肉鸡盲肠扁桃体TLR2的表达无影响。Cheled-Shoval等[33]研究发现,出壳前3d给肉鸡胚胎注射MOS可提高小肠TLR4基因表达,但对小肠TLR2基因表达无影响。本研究结果与Yitbarek等[32]报道的研究结果相一致,但与Cheled-Shoval等[33]报道的TLR2基因表达结果不一致。同样地,Munyaka等[34]报道,饲粮添加MOS对42日龄肉仔鸡回肠和盲肠扁桃体TLR2 mRNA的表达无影响,但显著降低TLR4 mRNA的表达,而本研究发现饲粮添加MOS对盲肠TLR4 mRNA无显著影响。功能性寡糖如菊粉和低聚果糖除直接与肠道上皮细胞和肠道免疫细胞上的碳水化合物识别受体如某些TLRs、甘露糖受体、C型凝集素受体等受体结合以启动免疫应答而提高机体对病原菌的清除能力和保护肠道健康等作用外,部分被吸收的寡糖可引起局部如脾脏、肝脏和胸腺或系统免疫应答反应[35-36]。本研究发现,MOS除不同程度提高肠道TLR2和TLR4表达外,MOS还可不同程度地提高肝脏和脾脏TLR2和TLR4 mRNA的表达。由此表明,MOS除直接在肠道发挥免疫调节作用外,部分MOS被完整吸收后或其被肠道微生物作用后的代谢产物被吸收后调节机体其他组织尤其是免疫器官组织细胞TLRs表达并由此调节机体局部和系统免疫功能,但这需进一步的试验证明。此外,本研究发现,TLR2和TLR4 mRNA的表达存在组织差异性,在所测的四个组织中,以盲肠TLR2和TLR4 mRNA表达量最高。
动物体内源表达的抗菌肽如β-防御素和cathelicidins是TLRs激活后产生的关键免疫效应分子之一[3-4]。研究表明,表达于肠道上皮细胞和免疫细胞的重要模式识别受体TLR2可识别酵母细胞壁甘露聚糖,甘露聚糖与TLR2结合后激活信号通路分子而产生免疫效应分子介导天然免疫和获得性免疫应答反应。禽AvBD9和是Cath-B1广泛表达于鸡各种组织尤其是消化道和免疫器官的一种重要抗菌肽,具有很强的杀灭致病细菌和真菌活性,同时,在鸡的天然免疫和获得性免疫中亦发挥重要作用[6-9,12]。本研究发现,饲粮中添加适宜水平的MOS可不同程度地提高肉仔鸡回肠、盲肠、肝脏和脾脏AvBD9及Cath-B1基因的表达,与相应组织中TLRs尤其是TLR2表达结果呈现相似的变化。综上结果可知,MOS可能够通过提高肉鸡组织TLRs表达并由TLRs介导上调β-防御素和cathelicidins等抗菌肽表达而提高肉鸡天然免疫防御功能。
4 结论
综合本试验结果,饲粮中添加适宜水平的MOS可显著提高肉仔鸡生长性能,且不同程度地提高肉鸡肝脏、脾脏、回肠和盲肠中TLR2、TLR4、AvBD9和Cath-B1的表达。MOS可能通过调控肉鸡组织TLRs表达并由TLRs介导上调β-防御素和cathelicidins等抗菌肽的表达而提高肉鸡天然免疫防御功能。综合生长性能、免疫器官指数和天然免疫相关基因表达指标,日粮添加0.6 g/kg MOS可取得较佳效果。
猜你喜欢
动物营养学报(2022年8期)2022-09-01
今日农业(2022年10期)2022-07-03
湖南农业科学(2022年3期)2022-05-18
养殖与饲料(2021年5期)2021-11-30
大连民族大学学报(2021年1期)2021-11-28
中国饲料(2021年17期)2021-11-02
河南畜牧兽医(2020年5期)2020-01-14
河南畜牧兽医(2020年19期)2020-01-10
江苏农业科学(2019年8期)2019-08-20
中外医学研究(2017年15期)2017-06-29
