
腐胺对高龄女性卵巢的作用
2020-05-15 03:04甘冬英韩阁阁刘嘉茵胡艳秋
中国生育健康杂志 2020年3期
甘冬英 韩阁阁 刘嘉茵 胡艳秋
随着人口预期寿命的增长及工业化社会变革,女性生育年龄越来越高。在中国,由于二胎政策的放开,高龄孕妇越来越多。然而,高龄女性在不孕、流产、出生缺陷等方面存在更大风险。因为高龄女性卵巢功能下降、产生的卵母细胞质量变差。虽然辅助生殖技术帮助数百万女性实现了生育,但不能解决女性卵巢衰老的根本问题即卵子数量和质量下降,寻找一个理想的干预措施是解决生殖衰老的关键[1]。多种哺乳动物的卵巢组织在排卵前鸟氨酸脱羧酶(ornithine decarboxylase,ODC)活性(腐胺从头合成的限速酶)及腐胺短暂升高。高龄女性卵巢ODC活性和腐胺下降,且补充腐胺可以减少卵母细胞非整倍体率、减少胚胎畸形率,提示排卵前ODC活性和腐胺的升高可能是一个进化保护机制[2]。本文就高龄女性卵巢特点、腐胺的来源和体内调节及对卵巢的作用进行综述。
一、高龄女性卵巢的特点
高龄女性卵巢特点主要表现在卵巢储备功能下降及卵母细胞质量降低。卵巢储备功能降低包括与年龄相关生理性和与年龄无关非生理性卵巢储备功能降低。本文主要关注前者即人类生育潜能随年龄增长而下降,表现为妊娠率降低、流产率增加、生育平均间隔时间延长等。卵巢储备参考评估指标很多,如卵巢超声检查基础窦状卵泡数(antral follicle count,AFC)、抗苗勒管激素(serum anti-Mullerian hormone,AMH)、基础卵泡刺激素(follicle stimulating hormone,FSH)、雌二醇、抑制素 B(inhibin B,INH B)等。本文简要描述高龄女性卵巢以AFC、AMH为代表的卵巢储备下降特点及自由基、线粒体DNA突变对卵母细胞质量的影响。
1.卵巢储备功能下降
(1)基础窦状卵泡数降低。卵巢中的卵泡出生时约100~200万个,到月经初潮时,卵巢中仅剩余约30~40万个卵泡,在绝经期时卵巢残留的卵泡数不足1000个。 女性一生有超过99.9% 的卵泡都相继发生闭锁,仅400~500 个始基卵泡发育成熟并排卵。研究发现,在20~41岁月经规则女性中,小窦卵泡(2~7 mm)每年下降4.4%,提示基础窦状卵泡数随着年龄的增加而逐渐降低[3]。此外,不孕女性中只有40.0%窦卵泡数大于10个,而86.7%正常生育力女性的窦卵泡数大于10个,说明不孕女性比正常生育力女性下降更明显[4]。
(2)血清抗苗勒管激素下降。AMH属于转化生长因子家族中一员,由小卵泡中颗粒细胞分泌。AMH在月经周期中变化不大,是反应卵巢储备功能的良好指标。AMH在幼年期几乎检测不到,在青春期启动时快速提高。研究发现25至35岁之间的女性,AMH每年下降1.4 pmol/L,大于35岁时,每年下降0.7 pmol/L,大于40岁时,每年下降1.4 pmol/L,到了更年期则检测不到,说明AMH随年龄的增长呈逐渐下降趋势[5]。
2.卵母细胞质量下降
(1)自由基及抗氧化系统。对不同月份的雌性C57BL/6小鼠研究发现,随着年龄的增加,卵巢间质细胞及卵泡氧化脂质、蛋白、DNA含量增加,抗氧化基因、线粒体及胞质中的抗氧化酶基因表达减少。说明在卵巢衰老过程中,卵巢细胞内抗氧化防御能力下降,清除自由基能力降低,造成自由基过量堆积,出现氧化应激反应,导致卵巢抗氧化损伤增加[6]。
(2)线粒体DNA突变及缺失。线粒体分布于能量需求最大的卵质区域,如减数分裂纺锤体的组装和拆卸。随着年龄增加,卵母细胞线粒体DNA(mitochondrial DNA,mtDNA)不稳定性增加、DNA突变增加,mtDNA数量减少,出现形态及功能的异常。表现为线粒体肿胀、空泡化、嵴改变,导致线粒体氧化磷酸化异常、供能障碍,最终导致卵母细胞的发生及成熟过程出现异常,如卵母细胞减数分裂时纺锤体形成不佳,不利于染色体分离,增加卵母细胞非整倍体率[7]。
二、腐胺的来源及其在体内平衡调节
腐胺是一种二胺,化学结构为NH2-CH2CH2CH2CH2-NH2,水溶性,存在于多种器官如胰腺、胸腺、皮肤、脑、子宫、卵巢。且随着年龄的增加,腐胺水平逐渐下降。腐胺也普遍存在食物中,如麦芽、青椒、大豆、开心果、橙子等[8]。腐胺的来源有外源性和内源性两种途径。外源性经典来源:精氨酸和脯氨酸通过代谢生成鸟氨酸,鸟氨酸在ODC作用下脱羧基转化生成腐胺;外源性非经典来源:精氨酸在精氨酸脱羧酶(arginine decarboxylase,ADC)作用下合成胍丁胺,胍丁胺在胍丁胺水解酶作用下生成腐胺。内源性来源:精胺与亚精胺/精胺N1-乙酰转移酶(N1-acetyltransferase,SAT1)、多胺氧化酶(polyamine oxidase)的结合,转化为亚精胺,亚精胺同样途径转化为腐胺[9]。见图1。
腐胺稳定主要依靠三方面调节:合成调节、分解代谢调节、细胞内外的转运调节。ODC是腐胺合成的限速酶,合成调节最主要的是调节ODC活性。鸟氨酸脱羧酶抗酶(ornithine decarboxylase antizyme,AZI)与ODC的26s蛋白酶体结合促进ODC降解,鸟氨酸脱羧酶抗酶抑制剂(ornithine decarboxylase antienzymeinhibitor,AZIN)与AZI竞争,以更高亲和力结合ODC,抑制ODC被AZI降解,AZI-AZIN通过控制ODC活性来调节腐胺的合成。此外,腐胺可以负反馈抑制ODC、S-腺苷甲硫氨酸脱羧酶(s-adenosylmethionine decarboxylase,SAMDC)、SAT1和AZIN翻译,减少腐胺合成原料、促进腐胺-亚精胺-精胺正向合成、抑制其逆向转化,从而负向调节自身合成。在细胞内外转运调节中,腐胺和肝素化磷脂酰肌醇聚糖-1分子(heparansulfate on glypican-1 molecules,HSG)结合,以受体介导的细胞内吞作用进入细胞内,细胞内游离腐胺通过可溶性载体SLC3A2转出。此外AZI减少腐胺转入细胞内,而AZIN通过与AZI负性作用刺激腐胺摄取[10]。见图1。
三、腐胺对卵巢的作用
1.腐胺对卵母细胞的影响
(1)降低卵子非整倍体率。高龄女性面临生育难题的很大原因归于卵子非整倍体率高,其将导致胚胎染色体异常、流产或出生缺陷。高龄卵子非整倍体率高于年轻女性,因为卵子在长达10~50年的发生过程中减数分裂时同源染色体或/和姐妹染色单体错误分离。这可能是因为染色体着丝粒和染色体臂的黏连蛋白随着年龄的增长逐渐减少,也可能是纺锤体形成异常以致染色体非直线排列于赤道板上[2],最终导致染色体分离障碍。研究发现排卵前高龄小鼠卵巢中的ODC和腐胺含量降低,补充腐胺可改善妊娠结局[2,11],这表明腐胺在短暂的排卵过程中扮演着重要角色。如果促使高龄小鼠排卵前ODC升高,能使卵子非整倍体率从12.7%降至5.3%。而高龄女性ODC活性在排卵前较年轻女性低[2],ODC降低导致卵子非整倍体增加。卵母细胞减数分裂在排卵期的短暂时间排出第一极体,成为次级卵母细胞,这说明腐胺对卵母细胞成熟过程中染色体的正确分离起重要作用。此外,未成熟卵母细胞体外成熟过程中添加鸟氨酸脱羧酶抑制剂α-二氟基鸟氨酸(α-difluoromethyl ornithine,DFMO),卵母细胞非整倍体率明显增加,可能是ODC在减数分裂I期中保护靠近着丝粒的黏连蛋白,促进同源染色体分开的同时,保护两条姐妹染色单体,减少其过早分离,从而减少卵母细胞非整倍体率[12]。
(2)促进卵母细胞质成熟。研究表明大多数自发性流产胚胎是整倍体[13],说明非整倍体不是造成胚胎发育不良的主要原因,但在高龄小鼠中补充腐胺可降低胎儿自发流产率(从60.0%至28.0%)[11]。可能腐胺对卵母细胞核及胞质质量扮演重要角色。还有研究表明,去卵质卵子与正常精子结合形成的受精卵发育成囊胚时OCT4阳性细胞比例丧失,但高龄小鼠卵母细胞体外成熟过程中添加腐胺可以提高囊胚内细胞团OCT4阳性细胞比,可能是因为腐胺促进卵细胞胞质成熟从而提高胚胎发育潜能[14]。

图1 腐胺的来源及其调节注:ADC:arginine decarboxylase精氨酸脱羧酶;SAT1:N1-acetyltransferase亚精胺/精胺N1-乙酰转移酶;ODC:ornithine decarboxylase 鸟氨酸脱羧酶;AZI:ornithine decarboxylase antizyme鸟氨酸脱羧酶抗酶;AZIN:Ornithine decarboxylase antienzymeinhibitor鸟氨酸脱羧酶抗酶抑制剂;HSG:heparansulfate on glypican-1 molecules肝素化磷脂酰肌醇聚糖-1分子。
正常发育的卵母细胞含有丰富的线粒体,但它不是均匀分布于卵母细胞胞质中,而是聚集于能量需求最大的卵质区域,减少与线粒体呼吸相关的活性氧(reactive oxygen species,ROS)的产生[2]。随着年龄的增加,卵母细胞成熟间期的线粒体分布、形态、功能出现异常[7,15],氧化还原调节失去平衡,产生大量ROS,诱导卵母细胞加速凋亡。减数分裂周期G2/M转换展示高水平的ROS,同期ODC活性提高,提示ODC有对抗ROS对卵母细胞损害作用,同时发现腐胺可以降低非洲爪蟾蜍ODC吗啉反义寡聚核苷酸(morpholino oligonucleotides antisense to xenopus ODC,xODC mo)引起的卵母细胞ROS升高,说明腐胺可以改善卵母细胞线粒体氧化还原反应失调,改善卵巢微环境,促进卵母细胞胞质优化[16]。
2.腐胺对黄体生成素的作用
研究表明在规律月经周期的不同女性中,排卵前黄体生成素峰(Luteinizing hormone surge,LH surge)在形态(单峰、平台型、双峰、多峰)、振幅和持续时间上都有很大变异[17]。它是即将排卵的可靠标志,在排卵中是不可或缺的,可促进卵母细胞恢复减数分裂排出第一极体,成为次级卵母细胞。此外,LH峰促进排卵前卵泡黄素化,产生少量孕酮,二者协同激活卵泡液内蛋白溶酶活性,消化卵泡壁部分胶原形成排卵孔,LH在整个排卵过程中扮演重要角色。但与ODC、腐胺相似,高龄女性LH峰振幅降低[18],故而提示ODC、腐胺和LH是否有一定的关联?
(1)腐胺对LH峰形成的影响。Aslam等人的体外试验表明,用黄体生成素释放激素(luteinizing hormone releasing hormone,LHRH)刺激大鼠动情前期的垂体时,ODC和LH升高,DFMO可抑制ODC和LH的升高,但腐胺可以消除DFMO降低效应。奇怪的是,腐胺不能提高LH峰振幅或者增大垂体对LHRH反应性,猜测腐胺在LH峰形成过程中不是必须的,只是为LH峰的形成创造条件,促进LH峰的形成[19]。但体内试验研究发现,在LH峰形成前,DFMO可以降低下丘脑LHRH水平,从而降低LH峰的振幅和总LH含量(图2),说明ODC和腐胺是LH峰形成必须的,总之,体内体外试验证明腐胺在LH峰形成中扮演着重要角色[20]。
(2)腐胺和LH对排卵的影响。排卵前LH可以提高卵巢ODC活性,说明LH可以控制ODC活性。且LHRH只在大鼠动情前期提高ODC水平,在动情间期对ODC无影响,提示ODC与排卵相关[21]。用xODC mo抑制ODC时,虽不影响卵母细胞第一极体的释放,但影响减数第二次分裂中期赤道板上染色体排列,抑制卵母细胞核成熟,导致胚胎碎片化(图2),说明腐胺是产生健康卵子所必须的[16]。此外,DFMO抑制动情前期ODC时,卵子数目减少70.0%(图2)[20],DFMO导致排卵障碍可能是通过抑制ODC对LH反应性,也可能是通过钝化LH峰,减弱LH的排卵作用,具体机制尚不明确。
(3)腐胺和LH对黄体化过程影响。在小鼠动情前期用DFMO抑制ODC活性后,通过减少卵巢中类固醇生成因子-1、细胞色素胆固醇侧链裂解酶、类固醇合成急性调节蛋白的表达,降低血浆和卵巢孕酮水平,同时减少黄体血管密度。推测排卵前的ODC升高是排卵时卵泡颗粒细胞和卵泡膜细胞黄体化的关键,结合LH峰是黄体化所必须的且颗粒细胞和卵泡膜细胞中ODC对LH高度反应性,说明ODC是LH在卵泡发育和早期黄体化过程中的重要介质[22]。黄体分泌的孕酮是囊胚植入必须的,因此抑制排卵前ODC的活性将导致不孕,原因可能是:ODC的缺乏不利于颗粒细胞和卵泡膜细胞充分黄体化,使产生的孕酮不足[23]。
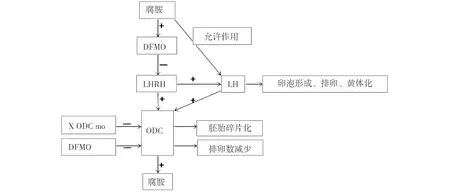
图2 腐胺和ODC对黄体生成素的作用注释:DFMO:α-difluoromethyl ornithine α-二氟基鸟氨酸;LHRH:Luteinizing Hormone Releasing Hormone黄体生成素释放激素;xODC mo:morpholino oligonucleotides antisense to Xenopus ODC非洲爪蟾蜍ODC吗啉反义寡聚核苷酸。
四、展望
高龄女性腐胺在排卵前升高不如年轻女性明显,且腐胺在排卵过程中可以提高卵母细胞质量,因此推测排卵期补充腐胺可能是补救卵巢衰老的理想干预措施之一。腐胺是机体自然产生的;外源性补充腐胺很容易被吸收,且停止补充后,腐胺会被快速清除,对母亲和胎儿都是安全的;补充腐胺所需要的干预时间很短;在IVF和自然受孕都可以应用[11],但腐胺因在人体中研究较少,其有效性及安全性尚不能证实,还尚未应用于临床中,需更多的研究证实其效用及安全性。
猜你喜欢
医学研究生学报(2022年5期)2022-12-07
临床肺科杂志(2022年3期)2022-11-26
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中国畜牧杂志(2022年4期)2022-04-15
中国畜牧杂志(2021年5期)2021-12-05
中国卒中杂志(2021年7期)2021-11-29
中国畜牧杂志(2020年8期)2020-12-18
保健与生活(2020年18期)2020-09-26
中国畜牧杂志(2020年6期)2020-07-11
西北农业学报(2020年5期)2020-06-04
