
水稻GLW7基因功能标记的开发和基因效应分析
2020-05-27 09:41梁文化孙旭超陈涛岳红亮田铮赵凌赵庆勇赵春芳朱镇张亚东王才林
江苏农业学报 2020年2期
关键词:水稻
梁文化 孙旭超 陈涛 岳红亮 田铮 赵凌 赵庆勇 赵春芳 朱镇 张亚东 王才林


摘要:根据水稻粒长和粒质量调控基因GLW7已知功能位点的核苷酸差异设计分子标记,并对国内外搜集的315份籼、粳稻品种资源进行基因型检测,分析其不同基因型的分布。同时,通过粒型性状的测定,分析该基因的遗传效应,评估其育种利用价值。结果表明,设计的功能标记能准确、有效地区分出201 bp和190 bp 2种带型,即大粒(Large grain haplotype,LGH)和小粒(Small grain haplotype,SGH)2种等位变异。从籼、粳亚种间的基因型分布来看,籼亚种中LGH和SGH的比例分别为95.65%和4.35%,而粳亚种中LGH和SGH比例分别为25.50%和74.50%,2种等位变异的分布在籼粳亚种间存在明显差异。粒型的测定结果表明,含不同等位变异的品种在粒长、粒厚、长宽比和千粒质量上存在显著或极显著差异,而粒宽则没有明显差异。进一步通过籼、粳分类分析发现,亚种间2种等位变异对粒型的效应并不完全一致,但都具有提高籽粒质量的作用。
关键词:水稻;粒型;GLW7基因;功能分子标记
中图分类号:S511.032文献标识码:A文章编号:1000-4440(2020)02-0257-08
Abstract: In this study, molecular markers were designed according to the nucleotide differences of functional loci of GLW7 gene regulating rice grain length and grain weight. Genotypes of 315 indica and japonica varieties collected at home and abroad were detected, and the distribution of different genotypes was analyzed. Simultaneously, the genetic effects of the gene were analyzed, and its breeding value was evaluated by the determination of grain shape traits. The results showed that the designed functional markers could accurately and effectively distinguish 201 bp and 190 bp bands, which was corresponding to large grain haplotype (LGH) and small grain haplotype (SGH), respectively. According to the genotype distribution between indica and japonica subspecies, the proportion of LGH and SGH in indica subspecies was 95.65% and 4.35%, while that was 25.50% and 74.50% in japonica subspecies, respectively. It was indicated that there was a significant difference in genotype distribution between two rice subspecies. The results of the grain size measurement showed that there were significant or extremely significant differences in grain length, grain thickness, length-width ratio and thousand grain weight among varieties with different allele variations, but there was no significant difference in grain width. Further analysis showed that the genetic effects of two allelic variations on grain shape were not identical between indica and japonica varieties, but the two allelic variations could increase grain weight.
Key words:rice;grain shape;GLW7 gene;functional molecular marker
水稻粒型包括粒长、粒宽和粒厚3个基本要素,是影响产量和品质的重要农艺性状[1-2],因此粒型基因的发掘和利用备受重视。目前水稻中已经定位了400多个与粒型相关的QTL,克隆的粒型基因已经超过60个,其中控制粒长的主要基因有GS3、GL3.1/qGL3、GLW7、OsLG3、TGW3、OsMADS1和GS9等[3-12],控制粒宽的基因有GW2、GW5/qSW5、GS5、GW7和GW8等[13-18],控制粒厚的基因有WTG1[19-20]。從对粒型的效应看,GS3、qGL3、GW2、qSW5/GW5、GS9等基因对粒型具有负调控效应,而GS5、GW8、GW7、GLW7、OsLG3、OsMADS1等基因则对粒型具有正调控作用。大量的研究结果证实,通过调节籽粒性状可以增加千粒质量,从而提高水稻产量。
GLW7基因编码植物特异性转录因子OsSPL13,具有SPL家族特有的保守结构,能调节多个重要的生物学进程[8]。GLW7基因正调控粒长、粒厚和粒质量,而对粒宽没有明显的影响。在OsSPL13位点5′-UTR区域的一个6 bp的串联重复序列CACTTC重复次数的变化是影响该基因转录与表达的关键。Si等[8]通过对47个热带粳稻品种OsSPL13位点的测序分析,发现该位点可分为2种不同的变异类型,即6个bp的CACTTC序列重复1次和2次两种类型,而对部分籼稻品种进行分析只检测到该序列重复1次的等位型。进一步研究发现大粒水稻的OsSPL13位点6 bp序列重复1次,而小粒水稻品种该序列重复2次,根据两种基因型对应的籽粒形态特征,分别用大粒型(LGH)和小粒型(SGH)表示。通过对基因碱基序列的进化分析和1 040个水稻品种7号染色体遗传差异的研究,推测热带粳稻中大粒等位型是籼稻品种渐渗而来的。通过对野生型和突变体的种子外稃长轴方向上细胞数目和大小分析,发现GLW7是通过增加细胞体积而使籽粒变大的。进一步研究证实GLW7能够与DEP1基因互作,同时对穗长、一次枝梗数目和二次枝梗数目产生显著影响[8]。因此,GLW7基因可以显著改善籽粒大小和穗粒结构,在水稻育种中具有重要的利用价值。
分子标记辅助选择是作物育种中利用优异基因的有效手段[21],通过开发粒型基因分子标记能够准确、高效地鉴定调控作物籽粒大小的有利等位变异[22-25]。为加快GLW7基因有利等位变异在水稻育种中的利用,本研究根据GLW7基因已知功能位点的核苷酸差异设计分子标记,并对国内外搜集的315份籼、粳稻品种资源进行基因型检测,分析其不同基因型的分布,同时,通过粒型性状的测定,分析该基因的遗传效应,评估其在育种中的利用价值。
1材料与方法
1.1试验材料
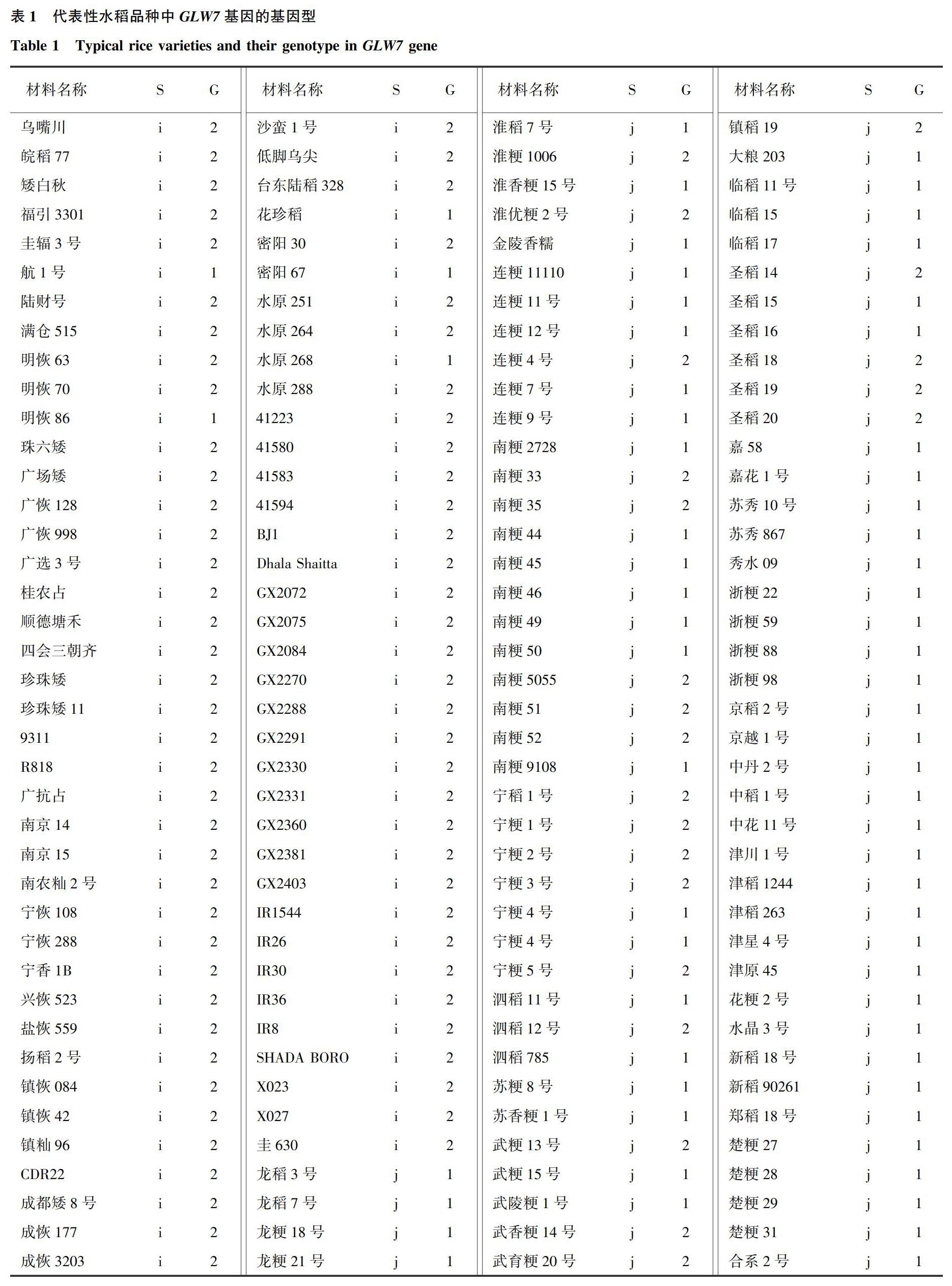
试验材料为江苏省种质资源保护与利用平台保存的315份国内外籼、粳稻品种,其中籼稻115份,粳稻200份(表1)。上述材料于2017年种植于江苏省农业科学院试验田,5月10日播种,6月10日移栽,每个品种种植4行,每行10株,株距为13.5 cm,行距为16.5 cm,常规栽培和管理。
1.2水稻成熟种子粒型相关性状数据的测定
成熟后,每个品种按单株收取5个植株的种子。每个单株随机挑选10粒饱满种子使用游标卡尺(精度0.01 mm)测量粒长、粒宽和粒厚,用电子天平(精度0.001 g)测定单株1 000粒风干种子的质量(千粒质量)。每个性状以5株的平均值为最终的表型值。
1.3GLW7基因功能标记开发和引物合成
根据Si等[8]的报道,从NCBI(http://www.ncbi.nlm.nih.gov/)下载到小粒品种Dongjing(DJ)和大粒品种GP7 GLW7的基因组序列LT159866.1和LT159955.1。通过多序列比对分析,选取变异位点上、下游200 bp的核苷酸序列,利用Primer Premier 5.0 软件对GLW7基因5′端非翻译区(Untranslated region,UTR)中-146~-135 bp区域CCATTC串联重复的变异位点设计引物,并由北京擎科新业生物技术有限公司合成。
1.4DNA提取、PCR扩增和电泳检测
在水稻分蘖盛期,取新鲜幼嫩的叶片,采用CTAB法提取水稻基因组DNA[26],并进行PCR扩增。10 μl的PCR反应总体系包含:ddH2O 7.2 μl,模板DNA(20 ng/μl)1.0 μl,10×Buffer(25.0 mmol/L)1.0 μl,dNTP(2.5 mmol/L)0.2 μl,正、反向引物(10 μmol/L)0.4 μl,Taq DNA聚合酶(2 U/μl)0.2 μl。PCR扩增反应条件:94 ℃预变性5 min;94 ℃变性30 s,57 ℃复性30 s,72 ℃延伸1 min,30个循环;72 ℃延伸7 min,4 ℃保持2 min。引物扩增产物在9%聚丙烯酰胺凝胶上电泳分离,银染后在胶片观察灯上拍照、统计。
1.5数据分析
利用Excel(2016)和SPSS(22.0)软件进行粒型相关数据分析。
2结果与分析
2.1GLW7基因的碱基序列分析及功能标记开发
从NCBI网站下载小粒品种Dongjing(DJ)和大粒品种GP7的基因组序列LT159866.1(5 864 bp)和LT159955.1(5 840 bp)进行序列比对分析。结果表明,GLW7基因在大粒品种中5′-UTR区域-146~-135 bp CACTTC重复1次,而在小粒品种中重復2次,这与已报道的结果完全一致,而该短序列串联重复次数差异是影响GLW7基因表达水平和籽粒大小的关键。5′-UTR区域的6 bp短序列重复1次,GLW7基因表达水平升高,正调控颖壳细胞体积而使籽粒变大;而该短序列重复2次则降低了基因的表达水平,使籽粒变小[8]。在其转录起始位点(ATG)上游5′-UTR区域串联重复序列变异的位点附近分别设计上游引物GLW7-F 5′-TATCCCTTTCAACCTTTTCCA-3′和下游引物GLW7-R 5′-GACGACGAGCTAGTGCTACTGT-3′。序列比对发现在功能位点附近大粒品种GP7还存在5 bp的缺失,因而最终PCR扩增产物的差异是由两个邻近位置的插入缺失(InDel)造成的,因而小粒品种Dongjing(DJ)能扩增出201 bp的条带,而大粒品种GP7则能扩增出190 bp的条带(图1)。
2.2GLW7基因功能标记验证
为验证功能标记GLW7-F/GLW7-R在水稻品种中是否能进行有效扩增并区分GLW7基因的不同等位变异,从315份水稻资源中随机选取籼、粳稻品种各40个进行PCR扩增,通过聚丙烯酰胺凝胶电泳进行检测。结果显示每个品种都能扩增出单一、清晰的条带,没有非特异性扩增和拖尾现象,说明本研究设计的引物特异性较好,扩增效率高。从带型来看,所有籼、粳稻品种都能扩增出与大、小粒品种GP7和Dongjing(DJ)同样的190 bp或201 bp 2种类型的条型,分别代表GLW7基因功能变异区域6 bp短序列重复1次和2次的结果(图2、图3)。这表明,该功能标记能够准确、有效地区分籼、粳稻品种中GLW7基因2种等位变异。
2.3GLW7基因功能标记对不同水稻品种的基因型鉴定
为明确GLW7基因在籼、粳稻品种中的基因型分布以及对粒型的效应。利用功能标记GLW7-F/GLW7-R继续对所有的115份籼稻和200份粳稻资源进行基因型检测。结果显示大粒基因型和小粒基因型的品种数分别为161份和154份;115份籼稻品种中大粒基因型110份(95.65%),而200份粳稻品种中大粒基因型仅为51份(25.50%)。由此可见GLW7基因等位变异在籼稻和粳稻中的分布存在明显差异(表1)。
为分析GLW7基因不同等位变异对粒型的影响,我们在2017年对315份水稻品种资源进行粒型相关性状的测定。结果显示,含GLW7大粒等位变异的品种在粒长、粒厚、长宽比和千粒质量上极显著高于小粒等位变异的品种,而粒宽却极显著低于小粒等位变异的品种(表2)。
由于基因GLW7大粒型等位变异在籼、粳稻中的分布存在明显差异,为进一步明确GLW7基因不同等位变异在水稻亚种中的效应,在籼、粳分类的基础上根据对基因型的检测结果进行分析。结果显示,在籼稻中,含GLW7大粒等位变异的品种粒长、长宽比、千粒质量极显著高于小粒等位变异品种,其粒长平均增加11.79%,千粒质量平均增加7.96%,但二者的粒宽和粒厚却没有显著差异(表3)。而在粳稻品种中,两种基因型品种的粒长和千粒质量存在极显著差异,在粒厚上存在显著差异,但粒宽却没有显著差异,含GLW7大粒等位变异的粳稻品种粒长和千粒质量分别比小粒等位变异的粳稻品种增加2.23%和4.32%(表4)。这也证实了GLW7基因的大粒型等位变异确实对增加粒长、提高千粒质量具有重要的作用。
3讨论
表型选择是传统育种的主要方法,而基因型、环境以及它们之间的互作等因素往往影响了育种家对表型的判断,因而传统育种不仅准确性差,而且周期较长。分子标记辅助选择可以提高育种的效率和准确率,在品种选育中具有重要作用[27-29]。本研究基于基因GLW7功能位点的序列差异设计了特异性引物,并对315份来自国内外水稻品种进行基因型检测,发现该位点只有2种等位变异类型,设计的功能标记GLW7-F/GLW7-R能够准确、高效地区分不同品种等位变异。对115份秈稻和200份粳稻的基因型检测结果表明,GLW7基因大粒等位变异在籼稻中占95.65%,而在粳稻中仅占25.50%,说明该基因的等位变异在亚种中分布存在明显的差异。粳稻中大粒型的等位基因频率较低,这可能与不同地区对粳稻粒型的选择性有关,也可能与本研究中所用的粳稻材料多为圆粒型,而长粒型粳稻相对偏少有关。
粒型是水稻产量的重要构成因素,籽粒的大小和形状决定了水稻经济产量的潜力,对提高千粒质量有着重要的影响。对GLW7基因T-DNA插入突变研究结果表明,突变体的粒长,粒厚和粒质量均显著降低[8]。通过对315份水稻资源进行粒型性状的测定分析,发现GLW7基因2种等位型的品种在粒长、粒厚及千粒质量上均有极显著的差异。基因型为小粒型和大粒型的品种粒宽平均值分别为3.38 mm和2.98 mm,前者明显大于后者,进一步分析结果表明出现这种现象的根本原因是粳稻和籼稻之间的差异造成的。为深入研究GLW7基因对粒型的作用效应,将315个水稻品种按籼、粳分为2组分别进行分析。结果显示,籼稻中具有大粒等位变异的品种粒长和千粒质量显著增加,而粒宽和粒厚没有明显差异;粳稻中2种等位变异的品种粒长和千粒质量差异极显著,粒厚差异显著,粒宽差异不显著。这与之前报道的结果[8]一致。综上所述,GLW7基因在籼稻和粳稻中对粒型不同性状的效应并不完全一样,这可能是由籼稻和粳稻中不同遗传背景差异造成的,也可能与粳稻中其他粒型基因的相互作用相关。
利用分子标记辅助选择进行优异基因的聚合是水稻育种的趋势[30-31]。在粒型方面, GS3、GW8和GW7基因的聚合,能有效改良水稻的粒型,同时产量和品质也得到显著提高[17-18,32-33]。研究结果证明GLW7基因与GS3基因对籽粒的调控是独立的通路[8],推测聚合这2个基因可能对粒型改良有明显的作用,因而GLW7基因在粒型基因聚合育种中具有较大的潜力。本研究开发的功能标记可以有效地区分GLW7不同的等位变异,为该基因在育种中的利用奠定了基础。
参考文献:
[1]HUANG R Y,JIANG L G,ZHENG J S,et al. Genetic bases of rice grain shape: so many genes, so little known[J]. Trends in Plant Science,2013,18(4):218-226.
[2]徐正进,陈温福,马殿荣,等. 稻谷粒形与稻米主要品质性状的关系[J]. 作物学报,2004,30(9):894-900.
[3]FAN C C,XING Y Z,MAO H L,et al. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein[J]. Theoretical and Applied Genetics,2006,112(6):1164-1171.
[4]MAO H L,SUN S Y,YAO J L,et al. Linking differential domain functions of the GS3 protein to natural variation of grain size in rice[J]. Proceedings of the National Academy of Sciences,2010,107(45): 19579-19584.
[5]SUN S Y,WANG L,MAO H L,et al. A G-protein pathway determines grain size in rice[J]. Nature Communications,2018,9(1):851.
[6]QI P,LIN Y S,SONG X J,et al. The novel quantitative trait locus GL3.1 controls rice grain size and yield by regulating Cyclin-T1;3[J]. Cell Research,2012,22(12): 1666-1680.
[7]ZHANG X J,WANG J F,HUANG J,et al. Rare allele of OsPPKL1 associated with grain length causes extra-large grain and a significant yield increase in rice[J]. Proceedings of the National Academy of Sciences,2012,109(52):21534-21539.
[8]SI L Z,CHEN J Y,HUANG X H,et al. OsSPL13 controls grain size in cultivated rice[J]. Nature Genetics,2016,48(4): 447-456.
[9]YING J Z,MA M,BAI C,et al. TGW3 , a major QTL that negatively modulates grain length and weight in rice[J]. Molecular Plant, 2018,11(5): 750-753.
[10]YU J P,XIONG H Y,ZHU X Y,et al. OsLG3 contributing to rice grain length and yield was mined by Ho-LAMap[J]. BMC Biology,2017,15(1): 28.
[11]LIU Q,HAN R,WU K,et al. G-protein βγ subunits determine grain size through interaction with MADS-domain transcription factors in rice[J]. Nature Communications, 2018,9(1): 852.
[12]ZHAO D S,LI Q F,ZHANG C Q,et al. GS9 acts as a transcriptional activator to regulate rice grain shape and appearance quality[J]. Nature Communications, 2018,9(1): 1240.
[13]SONG X J,HUANG W,SHI M,et al. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase[J]. Nature Genetics,2007,39(5): 623-630.
[14]SHOMURA A,IZAWA T,EBANA K,et al. Deletion in a gene associated with grain size increased yields during rice domestication[J]. Nature Genetics, 2008,40(8): 1023-1028.
[15]WENG J F,GU S H,WAN X Y,et al. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight[J]. Cell Research,2008,18(12): 1199-1209.
[16]LI Y B,FAN C C,XING Y Z,et al. Natural variation in GS5 plays an important role in regulating grain size and yield in rice[J]. Nature Genetics, 2011,43(12): 1266-1269.
[17]WANG S K,LI S,LIU Q,et al. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality[J]. Nature Genetics,2015,47(8): 949-954.
[18]WANG S K,WU K,YUAN Q B,et al. Control of grain size, shape and quality by OsSPL16 in rice[J]. Nature Genetics, 2012,44(8): 950-954.
[19]HUANG K,WANG D K,DUAN P G, et al. WIDE AND THICK GRAIN 1, which encodes an otubain-like protease with deubiquitination activity, influences grain size and shape in rice [J]. Plant Journal, 2017,91(5): 849-860.
[20]WANG S S,WU K,QIAN Q,et al. Non-canonical regulation of SPL transcription factors by a human OTUB1-like deubiquitinase defines a new plant type rice associated with higher grain yield[J]. Cell Research, 2017,27(9): 1142-1156.
[21]程丽, 胡茂龙, 浦惠明, 等.M342抗除草剂基因CAPS标记的开发与应用[J]. 江苏农业学报, 2019,35(2):241- 247.
[22]丁丹,张亚东,赵春芳,等. 水稻粒长基因GS3和qGL3功能标记的设计及应用[J]. 江农业学报,2014, 30(6):1191-1197.
[23]王军,杨杰,许祥,等. 水稻千粒重基因TGW6功能标记的开发与利用[J]. 中国水稻科学,2014,28(5):473-478.
[24]裔传灯,王德荣,蒋伟,等. 水稻粒形基因GW8的功能标记开发和单体型鉴定[J]. 作物学报,2016,42(9): 1291-1297.
[25]丁丹. 水稻5个粒型相关基因的分子标记开发与效应分析[D]. 南京:南京农业大学,2014.
[26]卢扬江,郑康乐. 提取水稻DNA的一种简易方法[J]. 中国水稻科学,1992,6(1):47-48.
[27]任海,吕小红,杜萌. 多抗水稻分子标记辅助育种方法[J]. 江蘇农业科学,2017,45(19):154-158.
[28]孙大元,周丹华,张景欣,等. 广谱抗源H4中2个主效抗病基因的单基因系构建及评价[J]. 江苏农业学报,2017,33(1): 1-5.
[29]陈红萍,刘开浪,杨宙,等. 利用分子标记辅助选择改良水稻稻瘟病抗性[J]. 湖北农业科学, 2019,58(2):29-32.
[30]刘冬梅,娄喜艳,吴狄,等. 基于棉花转录组测序的SSR分子标记的开发[J]. 江苏农业科学,2019,47(7):32-35.
[31]田大刚,杨小双 ,陈子强,等. 利用分子标记辅助选择改良闽恢3301稻瘟病抗性[J]. 南方农业学报,2019,50(8):1665-1670.
[32]李扬,徐小艳,严明,等. 利用GS3基因功能性分子标记改良水稻粒型的研究[J]. 上海农业学报,2016,(1):1-5.
[33]NAN J Z,FENG X M,WANG C,et al. Improving rice grain length through updating the GS3 locus of an elite variety Kongyu 131[J]. Rice, 2018,11(1): 21.
(责任编辑:张震林)
猜你喜欢
世界热带农业信息(2021年7期)2021-08-11
格言·校园版(2021年30期)2021-07-06
好孩子画报(2021年3期)2021-04-02
农民致富之友(2020年8期)2020-05-11
农民致富之友(2020年1期)2020-02-11
农民致富之友(2019年30期)2019-10-21
农民致富之友(2019年23期)2019-08-16
农民致富之友(2019年14期)2019-05-24
新农村(2018年29期)2018-04-02
农产品市场周刊(2017年35期)2017-10-25
