
碱胁迫对大豆光合特性及内源激素含量的影响
2020-05-27 09:41曹帅杜仲阳刘鹏李志刚向殿军迟晓雪宫晓旭何德鑫
江苏农业学报 2020年2期
曹帅 杜仲阳 刘鹏 李志刚 向殿军 迟晓雪 宫晓旭 何德鑫

摘要:用NaHCO3和Na2CO3混合溶液对25 d苗龄耐碱大豆品种吉育257和碱敏感品种东农63进行7 d胁迫处理,通过分析混合碱胁迫对不同大豆品种光合作用、叶绿素荧光参数和内源激素含量的影响,探讨大豆激素与光合作用的作用机制,为耐碱品种的选育和盐渍土地的有效利用提供理论依据。结果表明:混合碱胁迫后,吉育257叶片SPAD值、光化学猝灭系数(qP)、赤霉素(GA)含量、生长素(IAA)含量和细胞分裂素(CTK)含量均呈先升高后降低的趨势,东农63呈持续降低趋势;两品种净光合速率(Pn)均低于对照,脱落酸(ABA)和ETH含量均呈持续升高趋势。大豆可以通过调节自身CTK和ABA含量来调控自身气孔的开闭状态。30 mmol/L混合碱胁迫对吉育257各性状有一定的促进作用,对东农63各性状的抑制不明显,大豆植株光合速率下降的原因可能是气孔限制或内源激素调节降低了植株自身的生理需求。两品种在60 mmol/L混合碱浓度下均受到不同程度影响,耐碱性品种所受影响小于敏感性品种;90 mmol/L混合碱浓度下两品种均受到严重影响,其中光合速率下降的原因可能是非气孔限制和激素合成能力下降。60 mmol/L混合碱胁迫浓度可能为大豆耐碱性临界点,该浓度下,吉育257的防卫能力明显优于东农63。
关键词:大豆;碱胁迫;光合作用;叶绿素荧光;内源激素
中图分类号:S565.101文献标识码:A文章编号:1000-4440(2020)02-0284-08
Abstract:The mixed solution of NaHCO3 and Na2CO3 was used to treat 25-day-old alkali-tolerant soybean variety Jiyu 257 and alkali-sensitive variety Dongnong 63 for seven days. The effects of mixed alkali stress on photosynthesis, chlorophyll fluorescence parameters and endogenous hormone content of different soybean varieties were analyzed. Moreover, the action mechanism of soybean hormones and photosynthesis was discussed to provide theoretical basis for the selection of alkali resistant varieties and the effective utilization of saline land. The results showed that the chlorophyll meter (SPAD) value, photochemical quenching coefficient (qP), gibberellic acid (GA) content, indoleacetic acid (IAA) content and cytokinin (CTK) content in leaves of Jiyu 257 increased first and then decreased, while those indicators of Dongnong 63 continued to decrease. The net photosynthetic rate (Pn) of the two varieties was lower than that of respective control, and the abscisic acid (ABA) and ethylene (ETH) tended to increase continuously. Soybean could regulate the opening and closing states of their stomata by adjusting the contents of CTK and ABA. The mixed alkali (30 mmol/L) stress had a certain promotion effect on various traits of Jiyu 257, but it had no obvious inhibition on traits of Dongnong 63. The reason for the decrease in photosynthetic rate of soybean plants might be the limitation of stomata or the regulation of endogenous hormones, which further reduced the physiological needs of the plants. The physiological traits of two soybean varieties were affected in different degrees under mixed alkali (60 mmol/L) stress, and the influence of alkali-tolerant was less than that of alkali-sensitive variety. The physiological traits of the two soybean varieties were seriously affected under the treatment of 90 mmol/L mixed alkali, and the decrease of photosynthetic rate may be caused by the decrease of hormone synthesis ability and non-stomatal limitation. When the concentration of mixed alkali was 60 mmol/L, soybean might reach the critical point of alkali tolerance. At this concentration, the defense ability of Jiyu 257 was obviously better than that of Dongnong 63.
Key words:soybean;alkaline stress;photosynthesis;chlorophyll fluorescence;endogenous hormones
目前中国盐渍化土地面积占全球盐渍土地面积的10.4%,尤其在北方土壤蒸发量大、干旱和半干旱地区及滨海地区[1-2]。中国盐渍土主要是由中性盐和碱性盐组成,与中性盐相比,碱性盐可导致大量离子毒害和pH胁迫,对生态造成的危害更大[3-4]。土壤盐碱化问题在中国乃至全球普遍存在,并且呈现持续增加状态,严重制约农业发展[2]。大豆是中国主要粮食、油料和饲用作物之一[5],属于中度耐盐碱作物,主要通过拒盐碱作用来适应盐碱胁迫[4]。研究碱胁迫下大豆光合生理特性和内源激素含量及对应的耐盐机制,对有效利用盐渍土地具有重要的理论和实践意义。碱胁迫除了直接抑制植物种子发芽和营养吸收之外,还可间接通过光合作用对植物正常生长进行影响,随碱浓度的升高和胁迫时间的延长,对植物的影响越强烈[6]。在将光能转化为化学能的光合过程中,PSII光系统在碱胁迫植物过程中具有重要地位[7]。然而盐碱胁迫造成植物光合特性发生变化的主要原因没有统一的观点。余汉基等[8]研究结果表明,盐胁迫导致反映PSII电子传递能力有关的荧光参数ETR显著下降。刘玉兰等[9]研究结果表明,低浓度的碱性盐对小粒大豆生长抑制作用不明显,较高浓度的碱性盐对其影响較大,大豆的净光合速率、气孔导度和蒸腾速率等光合参数均显著下降,胞间CO2浓度明显升高。叶绿素荧光参数的变化能反映与环境的关系[10]和环境胁迫对光系统的损伤程度[11],可以用于评价环境胁迫对植物造成的影响。植物激素在调节植物受到逆境胁迫过程中发挥着重要的作用[12]。脱落酸(ABA)可对多种逆境环境发生响应,盐胁迫可增加植物内源ABA含量,其中大部分与叶片水势有关[13]。生长素(IAA)可促进细胞生长和根系的发生[14]。赤霉素(GA)主要促进植物茎节的伸长生长和花芽分化,GA与其他激素相互作用,调节植物在盐胁迫下的生长发育[15]。李海洋等[12]研究结果表明,盐胁迫导致向日葵IAA、ABA和玉米素核苷(ZR) 3种激素含量呈先升高后降低的趋势,且随盐浓度增加变化趋势越大。植物激素对盐胁迫的反应过程中,往往并不是单一激素而是多种激素协同变化,即激素的平衡效应[16]。有关大豆耐碱性研究主要集中在抗氧化防御系统、渗透调节和形态指标等方面[17-18]。而碱胁迫下综合分析大豆叶片光合特性、荧光参数和内源激素含量变化的相关研究甚少。因此,为避免单一指标不能全面反映不同品种间存在的差异,本研究选取前期在18份大豆品种中筛选鉴定出的耐碱型大豆吉育257和碱敏感型大豆东农63两个品种,探讨不同碱浓度下大豆叶片光合特性、叶绿素荧光动力学特征和内源激素含量变化,在了解大豆幼苗耐碱机理的基础上,阐明大豆幼苗光合相关指标、内源激素含量与碱胁迫之间的关联,深入了解大豆激素与光合特性的作用机制,进一步了解大豆的耐盐机理,同时为耐碱品种的选育和盐渍土地的有效利用提供理论依据。
1材料和方法
1.1供试大豆材料
试验材料选用2个大豆品种,其中,吉育257为耐盐碱大豆品种,由吉林省农业科学院大豆研究所提供;东农63为盐碱敏感品种,由东北农业大学提供。这2个大豆品种是从18个大豆品种中经混合碱(NaHCO3∶Na2CO3=9∶1,摩尔比)溶液胁迫筛选出来的[19]。
1.2试验设计
试验于2018年6-7月在内蒙古民族大学温室内进行,采用水培方式种植。每个品种均挑选大小一致、无虫害的饱满种子,在5%的次氯酸钠溶液中消毒5 min,自来水冲洗3次,去离子水冲洗3次,待种子表皮无水分,置于培养皿中进行发芽。待胚根生长至1.5 cm左右移栽至水培盒内,在规格为32.5 cm×26.0 cm×11.5 cm的网盘中等距离相间摆放,每品种每盘20株。从移栽结束后开始计时,5 d后(第1片复叶长出)开始浇灌Hoagland营养液,25 d后开始胁迫处理。本试验根据前人研究的大豆耐受盐碱浓度范围[20]及内蒙古通辽市主要盐碱地盐碱组成成分[21],共设置0(CK)、30 mmol/L、60 mmol/L、90 mmol/L混合碱(NaHCO3∶Na2CO3的摩尔比为9∶1、pH值9.11±0.104)溶液进行胁迫处理。各种溶液均在1/2 Hoagland营养液(pH为7.8)基础上进行配制。胁迫7 d后对生长健壮的幼嫩叶片测定光合指标和荧光参数等指标。
1.3测定指标与方法
1.3.1叶绿素含量测定利用便携式叶绿素仪SPAD-502Plus测量大豆植株自上而下第3片复叶的中间叶片,每一叶片随机选取10处测量并取平均值。同一品种大豆3次重复。
1.3.2光合气体交换参数的测定光合指标选用LI-6400型便携式光合作用测定系统(Li-Cor.公司产品)进行测定。盐胁迫处理7 d后,于上午9∶00-11∶00对2个大豆品种叶片光合气体交换参数进行测定,包括净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。测定时采用固定红蓝光源进行试验,光强为1 200 μmol/(m2·s),为避免测定时环境 CO2浓度的变化对测定结果的干扰,将仪器的进气口与装有恒定CO2浓度的钢瓶相接,钢瓶CO2浓度配制成370 μmol/mol,温度设定为25 ℃,空气流速0.5 L/min,相对湿度60%,选取3株大豆的主茎第3片复叶的中间叶片上进行测定。
1.3.3叶绿素荧光参数的测定叶绿素荧光参数选用FMS-2脉冲调制式荧光测定系统(英国Hansatech公司产品)进行测定。在测定光合指标的同时先测定光适应下的稳态荧光相对电子传递速率(ETR)、光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)等参数;叶片经暗适应30 min后,测定初始荧光(Fo)、最大荧光(Fm)和PSII光量子效率(Fv/Fm)等参数。
1.3.4内源激素含量测定选取3株长势基本一致的大豆,取顶部第2片复叶中间叶片,用液氮速冻0.5 h后,转入-70 ℃冰箱保存备用。内源激素生长素(IAA)、细胞分裂素(CTK)、脱落酸(ABA)、赤霉素(GA)、乙烯(ETH)含量参照王庆美等的酶联免疫吸附方法(ELISA)进行测定[22]。样品处理方法为:将大豆叶片1.0 g于液氮中速冻,用80%甲醇溶液[含二叔丁基对甲苯酚(BHT)1 mmol/L]匀浆,4 ℃提取8 h,4 000 r/min离心15 min,沉淀用80%甲醇重复提取3次,合并上清液,氮气吹干,用PBSTG(0.01 mmol/L磷酸缓冲液、9 g/L NaCl、0.1% Triton X-100,pH=7.2)溶解定容,用于ELISA测定。试剂盒购于奇松生物科技有限责任公司,操作步骤按说明书进行。
1.4数据统计分析
采用Excel2003和SPSS 20.0 进行数据统计分析。
2结果分析
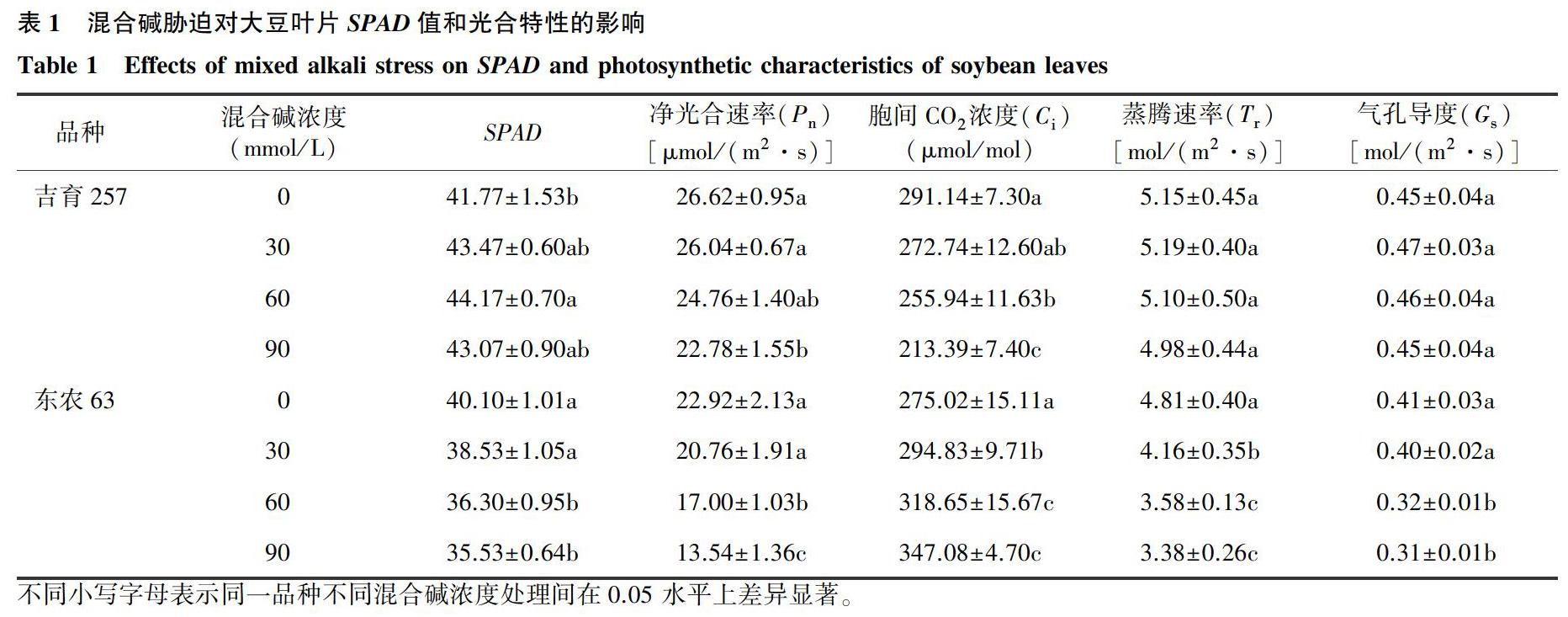
2.1混合碱胁迫对大豆叶片光合作用气体交换参数的影响
从表1可知,吉育257叶片SPAD值在各个胁迫浓度下均高于对照,在60 mmol/L混合碱胁迫下SPAD值升到最高,较对照增加5.75%,且达到显著差异水平。东农63叶片SPAD值呈一直下降趋势,在60 mmol/L和90 mmol/L混合碱胁迫下显著低于对照,相比于对照分別下降9.5%和11.39%。
两大豆品种净光合速率(Pn)对混合碱胁迫的响应呈下降趋势,在60 mmol/L混合碱胁迫下,东农63显著低于对照,吉育257差异不显著。在90 mmol/L混合碱胁迫下,两大豆品种均显著低于对照,吉育257相比于对照下降14.43%,东农63相比于对照下降40.93%。
吉育257叶片胞间CO2浓度(Ci)随混合碱浓度增加而降低,在60 mmol/L和90 mmol/L混合碱浓度处理下均显著低于对照,相比于对照分别降低12.09%和26.71%;东农63叶片胞间CO2浓度(Ci)呈现出相反趋势,在各个混合碱浓度处理下均显著高于对照,30 mmol/L、60 mmol/L和90 mmol/L混合碱浓度处理相比于对照分别增加了7.21%、15.86%和26.20%。
吉育257叶片蒸腾速率(Tr)除在30 mmol/L混合碱浓度下略有上升外,其余混合碱浓度处理下均低于对照,但差异均不显著;东农63叶片蒸腾速率(Tr)随混合碱浓度的增加呈现一直下降的趋势,且均显著低于对照,相比于对照分别降低13.46%、25.61%和29.69%。
吉育257叶片气孔导度(Gs)呈现先上升后下降的趋势,在90 mmol/L混合碱浓度下达到最低值,但是各个混合碱浓度处理均与对照没有显著性差异;东农63叶片气孔导度(Gs)随混合碱浓度的增加呈现一直下降趋势,在60 mmol/L和90 mmol/L混合碱浓度处理下均显著低于对照,相比于对照分别下降22.11%和25.80%。
2.2混合碱胁迫对大豆叶片叶绿素荧光参数的影响
从表2可知大豆叶片叶绿素荧光参数的变化趋势。初始荧光(Fo)是光系统II(PSII)反应中心处于完全开放时的荧光水平。受到混合碱胁迫后,吉育257 Fo呈现先降低后上升的趋势,且90 mmol/L混合碱浓度处理相比于对照增加4.29%,但各混合碱浓度处理均与对照没有显著性差异。东农63叶片Fo随混合碱浓度增强而升高,且在各胁迫浓度下Fo均显著高于对照,30 mmol/L、60 mmol/L和90 mmol/L混合碱浓度处理相比对照分别增加14.93%、17.72%和32.09%。
最大荧光(Fm)是PSII反应中心处于完全关闭时的荧光水平,可反映PSII的电子传递情况。在各个混合碱胁迫浓度下吉育257叶片Fm随胁迫浓度的增加呈现持续上升的趋势,而东农63呈现先降低后升高再降低的趋势,在60 mmol/L混合碱浓度处理下达到最高值,相比于对照增加6.33%,且叶片Fm在各处理下均低于吉育257。90 mmol/L混合碱浓度处理下吉育257叶片Fm显著高于对照和30 mmol/L混合碱浓度处理,分别增加16.38%和14.48%。
Fv/Fm为PSII光量子效率,主要反映PSII最大光能的利用效率。30 mmol/L混合碱浓度处理下吉育257叶片Fv/Fm相比于对照略有上升,其余2个处理均低于对照,但各混合碱浓度处理均与对照未达到显著差异水平。东农63叶片Fv/Fm随混合碱浓度的增强呈持续下降趋势,各处理相比于对照分别显著降低6.02%、8.43%和12.05%。
ETR为相对电子传递速率,受到混合碱胁迫后2品种叶片ETR均有所减小,且混合碱浓度越大,减小幅度越大。吉育257叶片ETR大于东农63,且下降幅度也小于东农63。受胁迫后吉育257在60 mmol/L和90 mmol/L混合碱浓度处理下显著低于对照,相比于对照分别下降17.61%和23.45%;东农63在各混合碱浓度处理下均显著低于对照,相比于对照分别下降20.64%、33.59%和39.27%。
光化学猝灭系数(qP)是反映PSII原初电子受体QA的氧化还原状态。受到混合碱胁迫后,吉育257叶片qP在60 mmol/L、90 mmol/L混合碱浓度处理下显著低于对照,在30 mmol/L混合碱浓度下高于对照,且达到显著差异水平;东农63叶片qp随混合碱浓度增加,呈一直下降趋势,相比于对照分别降低12.59%、18.10%和30.80%
非光化学猝灭系数(NPQ)是反映光系统对过剩光能的耗散能力。受到碱胁迫后两品种大豆叶片NPQ均随碱浓度升高而增加,且吉育257上升幅度小于东农63。吉育257叶片NPQ在60 mmol/L和90 mmol/L混合碱浓度处理下显著高于对照,分别增加15.09%和16.98%。东农63叶片NPQ在各个胁迫浓度下均显著高于对照,分别增加19.61%、33.33%和43.14%。
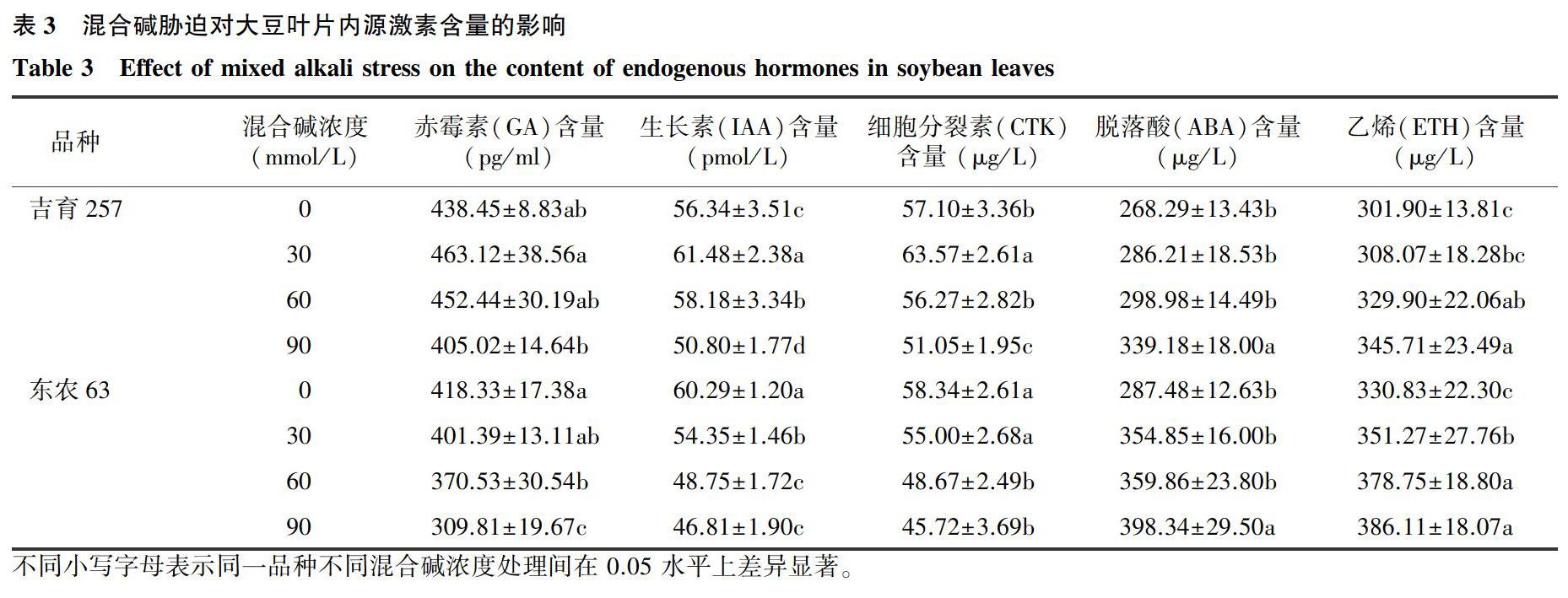
2.3混合碱胁迫对大豆叶片内源激素含量的影响
由表3可见,受混合碱胁迫后,吉育257叶片赤霉素(GA)、生长素(IAA)和细胞分裂素(CTK)含量变化均呈先升高后降低的趋势,而东农63均呈持续降低趋势。吉育257叶片GA含量在90 mmol/L混合碱浓度下显著低于30 mmol/L混合碱浓度处理,且在30 mmol/L混合碱浓度处理下达到最高值,较对照增加5.63%;东农63叶片GA含量在60mmol/L和90mmol/L混合碱浓度处理下显著低于对照,相比于对照分别下降11.43%和25.94%。吉育257叶片IAA含量和CTK含量在30 mmol/L混合碱浓度处理下均显著高于对照和其余2个混合碱浓度处理,相比于对照分别增加9.12%和11.33;东农63在各混合碱浓度下IAA含量均显著低于对照,分别下降9.85%、19.14%和22.36%;东农63 CTK含量在60 mmol/L和90 mmol/L混合碱浓度处理下显著低于对照和30 mmol/L碱浓度处理,相比于对照分别下降16.58%和21.63%。吉育257和东农63叶片脱落酸(ABA)含量和乙烯(ETH)含量均随混合碱浓度的升高而增加。吉育257和东农63叶片ABA含量均在90 mmol/L混合碱浓度处理下显著高于对照和其他2个混合碱浓度处理,相比于对照分别增加26.42%和38.56。吉育257叶片ETH含量在60 mmol/L和90 mmol/L混合碱浓度处理下显著高于对照,较对照分别增加9.27%和14.51%,而东农63叶片ETH含量在各混合碱浓度下均显著高于对照,相比对照分别增加6.17%、14.41和16.71%。
2.4混合碱胁迫下大豆各指标参数的相关性分析
由表4可见,Fv/Fm和qP与GA、IAA和CTK含量均达到极显著正相关水平,与ABA和ETH含量均达到极显著负相关水平。NPQ与GA、IAA和CTK含量均达到显著或极显著负相关水平,与ABA和ETH含量达到极显著正相关水平。GA含量除与Ci和Fm相关性较小外,与其他性状均达到显著或极显著相关水平,跟Pn的相关性最高,相关系数为0.965。IAA和CTK含量与SPAD值、Ci和Fm相关性较小,与其他性状均达到显著或极显著相关水平,均与Fv/Fm的相关性最高,相关系数分别为0.919和0.922。ABA和ETH含量除与Ci和Fm的相关性较小外,与其他性状均达到显著或极显著相关水平。
3讨论
3.1混合碱胁迫对大豆叶片光合参数的影响
盐碱胁迫主要影响植物生长和代谢,其中光合作用所受到的影响最为突出。由于植株叶片受盐碱胁迫后,其体内的叶绿素酶活性增强,进而导致叶绿素含量降低[23-24],而作物产量离不开光合作用,所以研究大豆光合特性对其耐碱性评价具有重要意义[25]。本研究结果表明,在各个混合碱浓度下,吉育257叶片SPAD值均高于对照,而东农63表现出相反趋势。盐碱胁迫是造成植物光合速率下降的关键因素,主要是引起渗透胁迫,导致气孔关闭,CO2由外界向内部扩散阻力加强,光合碳固定的底物减少所致[26]。本研究结果表明,吉育257叶片胞间CO2浓度(Ci)随混合碱浓度增加呈持续下降趋势,而东农63表现出相反趋势,两品种叶片Ci均在60 mmol/L和90 mmol/L混合碱处理下与对照达到显著差异水平。东农63叶片Ci升高的原因可能是混合碱胁迫导致细胞内盐离子大量积累,使叶片光合器官损伤,叶绿体结构被破坏,叶肉细胞光合活性下降,进而叶绿素含量降低,说明此时非气孔限制已成为促使光合速率降低的主要因子。叶片净光合速率和蒸腾速率也是光合作用中的重要指标,本研究结果显示,除吉育257叶片蒸腾速率在30 mmol/L混合碱浓度下略有升高外,两品种叶片净光合速率和蒸腾速率均随混合碱浓度的提高呈下降趋势,且东农63下降幅度更大,这与前人研究结果一致[27]。
3.2混合碱胁迫对大豆叶片叶绿素荧光参数的影响
叶绿素在植物光合作用中扮演重要的角色,植物叶绿素荧光参数可反映植物与环境的关系以及植物的生理状态,通过叶绿素荧光参数可间接了解光合作用的过程[28-29]。而叶绿素荧光参数和气体交换系数可直接反映植物对生境光强变化的生理调节过程,此外,叶绿素荧光参数在一定程度上还可反映光系统对光能吸收、传递、耗散等特点以及环境因子对植物的影响[30]。在盐碱胁迫下,Fm和Fv/Fm的下降表明植物叶片发生了光抑制,而Fo上升则表明PSII受到破坏[29]。本研究结果表明,受到混合碱胁迫后吉育257叶片Fm在各混合碱浓度下均高于对照,而东农63均低于对照;东农63叶片Fo在各个混合碱处理下均高于对照,吉育257除了在90 mmol/L混合碱浓度下高于对照外,其余2个混合碱浓度处理均低于对照。在混合碱胁迫条件下,两品种ETR和qP均有所下降,两品种叶片NPQ在各混合碱浓度下均高于对照,吉育257叶片NPQ小于东农63,这与杨淑萍等[31]研究结果相符。说明混合碱胁迫会改变大豆叶片 PSII激发能的分配方式,通过自身消耗更多的激发能来适应胁迫环境[23]。在混合碱胁迫下,吉育257叶片ETR、Fv/Fm、qP下降幅度明显小于东农63,叶片Fo和NPQ的增加幅度也明显小于东农63。吉育257在保持 PSII原初光能转换效率和 PSII潜在活性的同时,启动非辐射热能量耗散机制,将过剩光能以热的形式耗散掉,从而阻止净光合速率过度下降,起到保护光合系统的作用。
3.3混合碱胁迫对大豆叶片内源激素含量的影响
植物内源激素是重要的生长调节物质[32-33]。IAA、CTK和GA对植物生长主要表现为促进作用,而ABA和ETH则主要表现为抑制作用[12]。大豆可通过提高自身ABA含量来增加抗性,减缓细胞的分裂速度,从而降低生理需求以及水分蒸发,以此来适应外界的逆境环境。本研究结果表明,受混合碱胁迫后,耐碱性品种吉育257叶片GA、IAA和CTK含量呈先升高后降低的趨势,碱敏感性品种东农63呈持续下降趋势,2个大豆品种ABA和ETH含量均呈持续增加趋势,碱浓度越高,变化幅度越大,这与张敏等[16]研究结果相符。30 mmol/L混合碱处理对耐碱性大豆激素含量具有一定的促进作用,对碱敏感型大豆抑制不明显;60 mmol/L混合碱处理对耐碱性大豆抑制不明显,对敏感性大豆具有一定的抑制作用,可能是由于受到混合碱胁迫后大豆通过调节这几种内源激素含量来降低自身的生理需求,以此来抵抗胁迫;而90 mmol/L混合碱浓度下2个大豆品种均受到严重的伤害,可能是胁迫浓度过高,大豆不能再抵制胁迫所带来的伤害,IAA、CTK和GA合成能力下降,ABA与ETH合成能力升高,从而对大豆生长和光合能力都造成一定程度的影响。
3.4混合碱胁迫下内源激素与光合参数的相互关系
碱胁迫会诱导植物内源激素含量发生变化,对作物在逆境环境中生长及气孔运动具有重要作用。陈奇等[34]研究结果表明,油菜受到胁迫后气孔开闭状态与内源激素含量存在相关性。周宇飞等[35]研究结果表明,ABA含量升高会导致气孔导度降低。胁迫下高浓度的CTK可有效地维持气孔的开张[35]。因此,碱胁迫下大豆气孔导度的变化应该是ABA和CTK共同作用的结果,同时ABA和CTK还会对光合作用产生调节气孔开闭以外的其他直接影响。ABA可降低细胞膜的电势值,从而导致光合电子传递能力下降[36]。CTK可提高RuBPcase活性,从而促进光合电子的传递。本研究结果表明,气孔导度与ABA和CTK含量均达到显著相关水平,ABA含量降低与CTK含量升高均会使气孔导度升高,从而提高大豆光合能力,与前人研究结果相同[35-36]。IAA、CTK、ABA、GA和ETH含量与Fv/Fm均达到极显著相关水平,说明大豆受到胁迫时会通过调节各激素之间的关系,提高PSII最大光能利用效率,各激素之间的动态平衡可使大豆抵御碱害,保持正常代谢,提高自身光合能力。
4结论
碱胁迫可以抑制大豆PSII原初光能转化率和其潜在活性,增强PSII非辐射能的扩散,同时大豆可以通过调节自身CTK和ABA含量来调控自身气孔的开闭状态,各激素共同调节可提高PSII最大光能利用效率,以此来抵制胁迫,60 mmol/L混合碱胁迫可能为大豆耐碱性临界点。
参考文献:
[1]SETIA R, GOTTSCHALK P, SMITH P, et al. Soil salinity decreases global soil organic carbon stocks[J]. Science of The Total Environment, 2013, 465: 267-272.
[2]王佳丽,黄贤金,钟太洋,等.盐碱地可持续利用研究综述[J].地理学报,2011,66(5):673-684.
[3]DENG X X, ZHANG X Q, SONG X J, et al. Response of transgenic rice at germination traits under salt and alkali stress[J]. African Journal of Agricultural Research, 2011, 6(18): 4335-4339.
[4]季平,张鹏,徐克章,等.不同类型盐碱胁迫对大豆植株生长性状和产量的影响[J].大豆科学,2013,32(4):477-481.
[5]王树安.作物栽培学各论(北方本)[M].北京:中国农业出版社,1994.
[6]王佺珍,刘倩,高娅妮,等.植物对盐碱胁迫的响应机制研究进展[J].生态学报,2017,37(16):5565-5577.
[7]JEANJEAN R, MATTHIJS H C P, ONANA B, et al. Exposure of the cyanobacterium Synechocystis PCC6803 to salt stress induces concerted changes in respiration and photosynthesis[J]. Plant and Cell Physiology, 1993, 34(7): 1073-1079.
[8]佘汉基,张潮,薛立,等.盐胁迫对4种园林植物荧光特性的影响[J].生态科学,2018,37(5):87-93.
[9]刘玉兰,陈殿元,元明浩,等.盐胁迫对小粒大豆幼苗生长发育及光合特性的影响[J].大豆科学,2017,36(6):913-920.
[10]张培,郭俊杰,谌红辉,等.西南桦24个无性系的幼苗叶绿素荧光特性[J].西北林学院学报,2016,31(2):126-129.
[11]种培芳,杨江山.4种金色叶树木叶绿素荧光动力学参数对SO2胁迫的响应[J].西北林学院学报,2013,28(6): 14-19.
[12]李海洋,李爱学,王成,等.盐胁迫对苗期向日葵内源激素含量的影响[J].干旱地区农业研究,2018, 36(6):92-97.
[13]ZHANG J, JIA W, YANG J, et al. Role of ABA in integrating plant responses to drought and salt stresses[J]. Field Crops Research, 2006, 97(1): 111-119.
[14]JAVID M G, SOROOSHZADEH A, MORADI F, et al. The role of phytohormones in alleviating salt stress in crop plants[J]. Australian Journal of Crop Science, 2011, 5(6): 726.
[15]ACHARD P, CHENG H, DE GRAUWE L, et al. Integration of plant responses to environmentally activated phytohormonal signals[J]. Science, 2006, 311(5757): 91-94.
[16]張敏,蔡瑞国,李慧芝,等.盐胁迫环境下不同抗盐性小麦品种幼苗长势和内源激素的变化[J].生态学报, 2008,28(1):310-320.
[17]孟强,姜奇彦,牛风娟,等.盐胁迫下不同抗性野生大豆(Glycine soja)生理生化性状比较分析[J].中国农业科技导报,2017,19(8):25-32.
[18]姜夢婷,王秋岭,周鑫,等.不同油料作物油脂体氧化稳定性差异的研究[J].中国粮油学报,2018,33(10):63-70.
[19]曹帅,杜仲阳,向殿军,等.18份大豆品种耐盐碱性筛选与综合鉴定[J].大豆科学,2019,38(3): 344-352.
[20]牛陆. 盐、碱胁迫对大豆属植物的结构演化及生理特性的影响[D].长春:东北师范大学,2013.
[21]范富,张庆国,邰继承,等.通辽市盐碱地形成及类型划分[J].内蒙古民族大学学报(自然科学版),2009,24(4):409-413.
[22]王庆美,张立明,王振林.甘薯内源激素变化与块根形成膨大的关系[J].中国农业科学,2005(12):2414-2420.
[23]孙璐,周宇飞,李丰先,等.盐胁迫对高粱幼苗光合作用和荧光特性的影响[J].中国农业科学, 2012,45(16):3265-3272.
[24]赵澍,廖里平,张宏武,等.盐胁迫对酸枣幼苗光合生理特性的影响[J].干旱区资源与环境,2018,32(5):149-153.
[25]安飞飞,简纯平,杨龙,等.木薯幼苗叶绿素含量及光合特性对盐胁迫的响应[J].江苏农业学报,2015,31(3):500-504.
[26]裴斌,张光灿,张淑勇,等.土壤干旱胁迫对沙棘叶片光合作用和抗氧化酶活性的影响[J]. 生态学报,2013,33(5):1386-1396.
[27]刘伟成,郑春芳,陈琛,等.花期海蓬子对盐胁迫的生理响应[J].生态学报,2013, 33(17):5184-5193.
[28]黄杰,陈宗福,尹丽英,等.三种外源植物激素对黄花风铃木幼苗生物量、叶绿素荧光参数及光合特性的影响[J].植物科学学报,2018,36(5):745-754.
[29]周玉霞,巨天珍,王引弟,等.3种旱生植物的叶绿素荧光参数日变化研究[J].干旱区资源与环境,2019,33(5):164-170.
[30]敖金成,苏文华,张光飞,等. 不同光强下对马耳蕨叶绿素荧光参数的日变化[J].南京林业大学学报(自然科学版), 2011,35(1):135-138.
[31]杨淑萍,危常州,梁永超. 盐胁迫对不同基因型海岛棉光合作用及荧光特性的影响[J].中国农业科学,2010,43(8):1585-1593.
[32]赵凤,王小乐,房伟民,等. 外源激素和温度对切花菊侧芽萌发与内源激素含量的影响[J].江苏农业学报,2018,34(1):145-151.
[33]李小泉,韦莉萍,韦绍龙,等. 牛角蕉和桂蕉6号内源激素及主要酶系调控花柱脱落的作用机理[J]. 南方农业学报,2018,49(7):1351-1357.
[34]陈奇,袁金海,孙万仓,等.低温胁迫下白菜型冬油菜与春油菜叶片光合特性及内源激素变化比较[J].中国油料作物学报,2017,39(1):37-46.
[35]周宇飞,王德权,陆樟镳,等.干旱胁迫对持绿性高粱光合特性和内源激素ABA、CTK含量的影响[J].中国农业科学,2014, 47(4):655-663.
[36]魏道智,江力,张荣铣,等. ABA和ZT对小麦叶细胞质膜某些生理特性的影响[J].西北植物学报, 2002,22(6):1360-1364.
(责任编辑:张震林)
猜你喜欢
农民致富之友(2018年9期)2018-06-27
学苑创造·A版(2018年5期)2018-05-28
农产品市场周刊(2017年6期)2017-03-10
农产品市场周刊(2017年4期)2017-03-03
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
儿童故事画报·智力大王(2015年10期)2016-01-27
少儿科学周刊·儿童版(2015年10期)2015-11-07
幼儿智力世界(2015年5期)2015-08-20
小天使·二年级语数英综合(2015年6期)2015-05-14
