
2-甲基吡嗪分子激发态系间交叉过程的飞秒时间分辨光电子影像研究*
2020-06-04 09:45布玛丽亚阿布力米提凌丰姿邓绪兰魏洁宋辛黎向梅张冰
物理学报 2020年10期
布玛丽亚·阿布力米提 凌丰姿 邓绪兰 魏洁宋辛黎 向梅 张冰
1) (新疆师范大学物理与电子工程学院, 乌鲁木齐 830054)2) (中国科学院武汉物理与数学研究所, 波谱与原子分子物理国家重点实验室, 武汉 430071)(2020年1月14日收到; 2020年3月12日收到修改稿)
飞秒时间分辨光电子影像技术和飞秒时间分辨质谱技术相结合, 研究了2-甲基吡嗪分子电子激发态超快非绝热弛豫动力学. 用323 nm 光作为泵浦光, 把2-甲基吡嗪分子激发到第一激发态S1, 用400 nm 光探测激发态演化过程. 通过时间分辨质谱技术测得S1 态的寿命为98 ps. 实验中, 实时观察到了单重态S1 向三重态T1 的系间交叉过程. 通过分析发现, 跟吡嗪分子S1 态的动力学过程不同, 2-甲基吡嗪分子激发到S1 态后,不仅S1 → T1 系间交叉过程是S1 态主要衰减通道, S1 → S0 内转换过程也是S1 态另一个主要衰减通道. 发挥飞秒时间分辨光电子影像技术的优点, 实验上得到不同泵浦-探测时间延迟的光电子角分布, 从角分布信息结合光电子能谱信息, 尝试观察2-甲基吡嗪分子的非绝热无场准直, 但由于2-甲基吡嗪分子对称性比吡嗪分子更低, 对称性更低分子准直现象的观察更有挑战性, 在实验中未能观察到非绝热准直动力学. 本工作为2-甲基吡嗪分子S1 态非绝热弛豫动力学提供了较清楚的物理图像.
1 引 言
生物分子吸收紫外(UV)光后被激发到电子激发态, 会导致激发态光化学过程, 这些光化学过程可能对生物体产生潜在的危害[1,2]. 但由于许多重要生物发色团, 如DNA 碱基和氨基酸被激发后,通过超快非绝热电子弛豫过程回到基态, 表现出一定的光稳定性[3], 因此, 激发态非绝热电子弛豫过程为电子态能量快速消散提供了高效的路径. 非绝热耦合作用引起的内转换(IC)和旋轨耦合引起的系间交叉(ISC)是两种最主要的激发态超快非绝热电子弛豫途径[4]. 这些非绝热过程不仅是多原子体系光化学过程的基本步骤, 而且在许多生物过程如DNA 自我修复[5]、视觉产生过程、分子开关和光合作用[6]以及许多其他重要过程和应用中都必不可少. 因此, 研究分子吸收紫外光后的电子激发态内转换和系间交叉等非绝热弛豫过程是一个非常重要和有意义的课题. 虽然量子产率的测量等方法提供了理解非绝热动力学基本信息, 但直接观测超快非绝热动力学过程具有很重要的意义. 飞秒时间分辨的光电子影像技术[7,8]可以同时测量光电子能谱分布和光电子角分布, 可以在飞秒时间尺度上跟踪非绝热过程, 是深入研究分子电子激发态超快非绝热动力学非常有效的手段.
芳香族分子是大量生物分子的核心骨架. 在紫外光照射下, 芳香族分子将会被激发到电子激发态. 电子激发态被制备后, 由于激发态不稳定, 迅速启动伴随着能量流动的超快非绝热弛豫[9,10]过程. 苯和苯的衍生物[11,12]作为典型芳香族分子, 引起相关理论和实验研究者广泛关注. 含氮的芳香族分子电子激发态非绝热弛豫过程也引起研究者们极大关注和兴趣[13−18], 尤其是吡嗪(pyrazine)分子成为研究超快非绝热弛豫动力学过程最典型分子体系之一. 吡嗪分子第一次时间分辨光电子影像实验测得了吡嗪分子S1态寿命, 在带源附近S1态寿命为110 ps, 随着S1态振动能级的增加, 寿命瞬速减小, 在260 nm 的泵浦光泵浦时, 测得S1态寿命为23 ps[15]. Tsubouchi 等[19]和Suzuki 等[20]实验上观察到S1态向T1态的系间交叉过程, 从动能分布和角分布中, 观察到了吡嗪分子S1态和T1态电离的过程中和里德堡态共振, 在吡嗪分子中还观察到了含时的分子取向[16]. 跟其他含氮芳香烃化合物相比, 2-甲基吡嗪电子激发态非绝热弛豫过程的实验研究非常少, 很少用飞秒的方法进行研究,因此, 对2-甲基吡嗪分子激发态弛豫过程一直存在争议. 由于甲基的存在, 2-甲基吡嗪分子动力学很有可能跟吡嗪分子不同. 有关苯和苯衍生物研究中, 观察到取代基对电子激发态非绝热弛豫过程的影响. 例如, 实验中观察到甲苯分子S2→S1内转换比例明显高于苯分子, 这个现象被解释为由于CH3的引入增加了甲苯分子的S1态振动能级密度, 从而增加上态S2态与下态S1态之间发生非绝热耦合概率. 2-甲基吡嗪分子S1态被布局后通过什么样的衰减通道衰减? 苯衍生物非简并性导致了更高的振动能级密度和更低对称禁戒性, 2-甲基吡嗪分子中取代基的存在对吡嗪衍生物激发态非绝热动力学过程会有什么样的影响? 这些问题一直存在争议. 因此, 利用时间分辨的光电子影像技术研究2-甲基吡啶分子S1态动力学过程, 实时观察S1态弛豫过程是一个很必要和很有意义的工作.利用时间分辨的光电子影像, 可以同时测量光电子动能分布和角分布. 光电子信号衰减反映激发态布居的衰减过程[17,18], 可以决定不同激发态衰减寿命和弛豫通道. 光电子角分布的变化可以反映转动相干波包演化. 利用这些优点, 本文利用飞秒时间分辨技术研究2-甲基吡嗪在323 nm 泵浦光激发下S1态非绝热弛豫动力学. 实验中选择400 nm 光作为探测光, 进行多光子电离. 多光子电离与单光子电离相比, 在电离过程中由于探测光很有可能与较高的里德堡态发生偶然共振[19,20], 多光子电离对相近电子态之间非绝热耦合过程的探测更灵敏. 考虑2-甲基吡嗪分子比吡嗪分子对称性明显降低, 在吡嗪分子中原本简并的电子态在2-甲基吡嗪分子中其简并性很有可能被解除, 可以预测, 2-甲基吡嗪分子电子激发态超快非绝热动力学很有可能与吡嗪分子不同, 或更为复杂. 因此, 期待观察到2-甲基吡嗪分子S1态丰富的非绝热动力学过程.
2 实 验
本实验工作在自制的光电子影像装置上完成,实验装置实物图和示意图如图1 所示, 实验装置在之前的工作中已有介绍[21−23]. 光电子影像装置由超声分子束进样系统、真空系统、离子透镜、时序控制器, 二维影像探测系统、信号采集及数据处理系统等组成. 实验装置真空腔分为束源室和电离室两部分; 采用转速为400 转/s、抽速为1200 L/s 的分子泵(F200/1200)为束源室和电离室抽真空, 保持较高的真空度. 未进样时, 束源室和电离室都保持腔内真空度在10–6Pa 量级, 当开启脉冲阀进样时, 束源室真空度介于1.0×10–4—3.0×10–4Pa 之间. 为了电子不受外来电磁场的影响, 电离室利用µ金属(铁镍合金)屏蔽层进行屏蔽.
自锁模钛蓝宝石振荡器产生的种子光通过啁啾脉冲再生放大器进行放大, 输出重复频率为1 kHz、脉宽为100 fs、中心波长为800 nm、单脉冲能量为4.5 mJ/pulse 的基频光. 基频光分成两束,其中一束用于泵浦行波光参量放大系统(TOPAS), 产生中心波长为323 nm 的泵浦脉冲,对应共振激发2-甲基吡嗪分子S1态带源(30944 cm–1). 另外一束基频光进行倍频产生中心波长为400 nm 的二倍频光作为探测光. 泵浦光和探测光偏振方向分别利用可变波片和半波片调节为平行于探测器平面. 在不同泵浦-探测时间延迟下采集光电子影像, 再通过BASEX (BAsis Set EXpansion)变换重构其三维影像[24]. 实验中测得泵浦光(323 nm)和探测光(400 nm)交叉相关函数为260 fs.

图1 (a)光电子影像装置实物图; (b)飞秒时间分辨光电子影像装置示意图[21]Fig. 1. (a) Photoelectron imaging apparatus; (b) schematic diagram of the femtosecond time resolved photoelectron imaging setup.
纯度为99.9%的2-甲基吡嗪分子作为样品,未 进 一 步 提 纯. 背 景 压 力2 atm (1 atm =101325 Pa)氦气作为载气, 液体样品饱和蒸汽和载气混合气体经过脉冲阀喷入束源室形成超声分子束. 超声分子束再经过半径为1 mm 的锥形撇勺(skimmer)准直进入电离室, 在离子透镜排斥极(R)和加速极(E)之间, 与激光束发生作用.2-甲基吡嗪分子在泵浦光和探测光作用下被电离产生光电子和光离子, 离子透镜会对产生的光电子或光离子进行加速和聚焦, 最终很好地聚焦到由两块微通道板(MCP)和快速响应的P47 荧光屏构成的二维位置敏感探测器上, 形成高分辨的光电子影像或离子影像. 采集光电子影像时离子透镜三个极 板 采 用 的 电 压 配 置 为:VG= 0 V;VE=–2778 V;VR= –4000 V. 形成的影像利用装在探测器背面的CCD 相机进行采集, 实验中还利用光电倍增管, 采集光电子/离子质谱信号. 整个系统的时序利用时序控制器DG535 进行控制.
3 结果与讨论

图2 (a)泵浦光323 nm, 探测光400 nm 的各自单光和零时刻双光质谱;(b)泵浦 光323 nm, 探测光400 nm 作 用下时间分辨母体离子信号, 图中实线表示拟合曲线, 圆圈代表实验数据Fig. 2. (a) Two color (at time overlap) and one color mass spectra of 2-methlypyrazine at 323 nm pump and 400 nm probe; (b) time-resolved total ion signals of parent ion as a function of delay time between the pump pulse at 323 nm and the probe pulse at 400 nm. The circles are the experimental results, and solid lines are the fitting results.
323 nm 泵浦光、400 nm 探测光各自单光和零时刻双光飞行时间质谱如图2(a)所示, 实验中, 泵浦光和探测光能量控制为单光, 几乎没有信号. 从零时刻双光质谱中可以看出有两个峰, 分别对应母体离子和碎片离子C4H1N+. 计算得到两个峰强度之比为近40∶1, 表明碎片离子C4H1N+产生的光电子可以忽略不计. 此外, 在实验中采集母体和碎片离子信号随时间演化曲线, 并发现母体离子和碎片离子信号强度随时间演化曲线非常类似,可以判断实验中得到的光电子信号大部分都来自于母体分子直接电离.
图2(b)给出了母体离子信号强度随着泵浦-探测时间延迟变化的曲线. 该曲线可以用交叉相关函数和一个指数上升、一个指数衰减函数卷积拟合,并得到两个不同时间常数, 分别为Tdecay=98 ps 和Trise= 235 ps. 323 nm 泵浦光把2-甲基吡嗪分子激发到S1态, 拟合结果表明随着S1态的衰减, 另一个态开始布局, 由此可以推测分子被激发到S1态后, 发生了非绝热耦合过程.时间常数Tdecay= 98 ps 反映S1的衰减, 而Trise= 235 ps反映新布局态的上升. 虽然从时间分辨母体离子信号中可以推测2-甲基吡嗪分子S1态弛豫过程, 但为了得到2-甲基吡嗪分子S1态整个衰减动力学过程, 实验上测得了不同泵浦-探测时间延迟的光电子影像.
图3 显示了一组实验中得到的不同泵浦-探测时间延迟下的光电子影像, 上排给出原始影像, 是光电子三维分布在二维探测器上的投影, 而下排给出将原始影像经过BASEX 变换的三维重构光电子影像. 泵浦-探测时间延迟为0 ps时, 从光电子影像中可以明显看到有五个不同半径同心电子速率环, 随着泵浦-探测时间延迟增加, 这五个环逐渐减弱, 而最里面出现第六个环并逐渐加强. 在光电子影像中, 不同动能的光电子对应不同大小半径的圆环, 外环对应动能相对较大动能的光电子分布,而内环对应动能相对小的光电子分布.

图3 泵浦光323 nm, 探测光400 nm, 在不同时间延迟下的光电子原始影像和BASEX 变换后的影像(上排为原始影像, 而下排为BASEX 变换后的影像.). 泵浦光和探测光都是线偏振光, 偏振方向为图平面竖直方向Fig. 3. Time-resolved photoelectron raw images (shown in the upper row) and BASEX-inverted images (shown in the lower row) at various time delays observed at 323 nm pump and 400 nm probe.
从光电子影像中得到不同时间延迟下的光电子能谱, 如图4 所示, 光电子能谱中不同峰对应于光电子影像中不同环. 如图4(b)所示, 延迟时间为0 ps时, 观察到中心位置分别为1.05, 0.96,0.86, 0.73 和0.66 eV 的五个峰, 分别标记为1st,2nd, 3rd, 4th和5th峰. 当延迟时间为92 ps时光电子能谱中很清楚地观察到中心位置为0.15 eV 的第六个峰, 标记为6th峰. 外面的1st, 2nd, 3rd,4th和5th峰在延迟时间为0 ps时具有最大值, 并随泵浦-探测延迟时间增加表现出一样的变化趋势,故对应初始激发态S1态的电离. 随着时间延迟的增 加, 外 面 的1st, 2nd, 3rd, 4th和5th峰衰减, 而6th峰逐渐增强, 表明初始激发电子态S1向二次布局电子态发生了快速的能量转移过程. 此外, 可以看出这六个峰都比较尖锐, 因此, S1态和二次布局态在电离的过程中很有可能跟里德堡态发生偶然共振[25].
为了更直观地显示图4 中的能量转移过程, 将六个电子峰从整个动能分布中逐一切片出来, 并对每个电子带强度区间单独进行积分, 得到六个电子峰随泵浦-探测延迟时间变化趋势, 如图5 所示.1st, 2nd, 3rd, 4th和5th峰和6th峰不同衰减行为反映这六个光电子峰来自于不同的态的电离, 其中1st, 2nd, 3rd, 4th和5th峰来自S1的电离. 从图5 中可以观察到随着泵浦-探测时间延迟的增加动能较高的五个峰衰减, 而低能量6th峰上升, 反映了S1态和低态T1或S0态之间的非绝热耦合过程.S1态被布局后可能通过系间交叉过程衰减到T1态, 也有可能通过内转换过程衰减到S0态高振动态. 但S0态的高振动态有3.84 eV 的振动能, 这个态电离的Franck-Condon 因子会很小, 400 m探测光无法电离基态这么高的振动态, 因此, 图4中的6th峰应该来自于T1态的电离. 随着1st, 2nd,3rd, 4th和5th峰的衰减, 6th峰 开 始 布 居, 反 映 了S1态向T1态的系间交叉过程.

图4 (a)不同延迟时间下的光电子能谱; (b) 0 fs 和92 ps时的光电子能谱Fig. 4. (a) Photoelectron kinetic energy distributions (PKE)at different time delay; (b) photoelectron kinetic energy distributions at 0 and 92 ps.
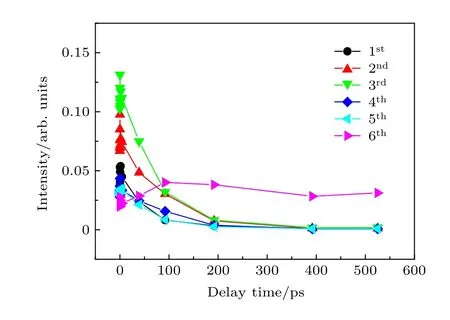
图5 六个峰强度随泵浦-探测时间变化图Fig. 5. Time-resolved PKE bands intensity as a function of representative delay times.
上面都是介于光电子能谱分布讨论可能发生的动力学过程. 但光电子影像与其光电子能谱相比, 优点在于不仅可以得到光电子动能分布而且可以得到光电子角分布. 光电子角分布对分子空间排布很敏感, 因此从光电子角分布随泵浦-探测时间延迟变化中可以得到转动波包演化. Tsubouchi 等[21]从时间分辨光电子角分布中观察到吡嗪分子含时的分子取向, Li 等[31]利用时间分辨离子产率和光电子角分布中观察到嘧啶分子无场准直现象. 实验中偏振方向相同的线偏振光作为泵浦光与探测光.并且泵浦光和探测光偏振方向平行于探测器表面.不同泵浦-探测时间延迟下光电子影像中可以得到光电子角分布, 将光电子角分布实验值拟合即可得到感兴趣能谱区域各向异性参数(b2,b4,b6).图6(b)显示六个峰各向异性参数b2, 由于b4和b6数值太小, 变化不明显, 图中没给出. 图6(a)给出泵浦-探测时间延迟下不同光电子峰强度变化.从图6(a)和图6(b)可以看出, 光电子能谱强度分布和各向异性参数b2在0—1 ps 有比较明显变化,1 ps 之后b2参数保持比较平稳. 根据Felker 和Zewail[32]提出的转动相干理论, 光电子能谱强度和各向异性参数在0—1 ps 内比较明显变化, 可能反映2-甲基分子的准直特征, 但由于各项异性参数值波动比较大, 不能完全确定分子的准直特征, 如果分子非绝热准直, 一个转动周期后可以明显观察到光电子角分布恢复到和零时刻一致. 通过计算得到2-甲基吡嗪分子转动周期为116 ps, 在实验中仔细采集105—130 ps 之间的光电子影像, 尝试观察分子转动恢复现象. 从光电子影像中得到光电子能谱分布和角分布, 如图7(a)和图7(b)所示. 仔细观察105—130 ps 泵浦-探测时间延迟下不同光电子峰强度变化和各向异性参数变化, 并未观察到光电子角分布恢复到和零时刻一致. 因此可以判断泵浦-探测时间延迟0—1 ps 之间的光电子能谱强度和各向异性参数的变化反映的不是分子准直特征. 由于2-甲基吡嗪分子对称性比吡嗪或嘧啶分子更低, 对称性更低的2-甲基吡嗪分子准直现象的观察更有挑战性, 此外, 一个转动周期时光电子信号已衰减到较弱, 在本实验中未能观察到2-甲基吡嗪分子呈现出非绝热准直特征.
实验中观察到了S1态向T1态的系间交叉过程, S1态寿命为98 ps, 而三重态T1态的布局时间为235 ps, S1态的衰减寿命比T1布局时间更短,这个表明S1态的衰减很有可能还有其他更快的通道. 由于光电子能谱中并未观察到其他态对应的光电子峰, 我们推测S1态可能衰减到400 nm 探测光无法探测的态上. S1态被布局后很有可能通过内转换衰减到S0的高振动态, 如果S1态通过内转换衰减到S0态, 400 nm 探测光无法探测S0的高振动态. 因此, 认为S1→S0的内转换过程是S1态衰减的另一个主要通道. 2-甲基吡嗪分子S1态带源寿命(98)略短与吡嗪分子S1态带源寿命(110 ps), 可能是因为较低对称性往往伴随着振动能态密度的增大和原本对称性不允许而禁阻的相互作用增加, 可以加快非绝热耦合过程, 从而缩短了S1态的寿命. 实验中, 离子信号和光电子信号最终都没有完全衰减完, 说明T1态寿命大于实验采用位移平台长度可测的范围800 ps, 2-甲基吡嗪分子T1态的寿命很有可能跟吡嗪分子T1态的寿命一样是ns 量级的寿命. Wang 等[18]有关吡嗪分子时间分辨光电子影像实验中指出, 在S1态带源附近S1态向T1态系间交叉量子产率约为1, 在S1态带源附近S1态向T1态系间交叉过程为S1态主要衰减通道, 没有观察到其他衰减通道. 与吡嗪分子S1态的动力学过程不同, 在2-甲级吡嗪分子中S1态带源附近S1→T1系间交叉和S1→S0的内转换都是S1态衰减的主要通道.
2-甲基吡嗪分子S1态动力学过程总结在图8中. 1st, 2nd, 3rd, 4th和5th峰 来 自S1态的电离,6th峰来自于T1态的电离, S1和T1态在电离的过程中和里德堡态发生偶然共振. S1态被布局后, 通过S1→T1系间交叉过程和S1→S0的内转换过程进行衰减.
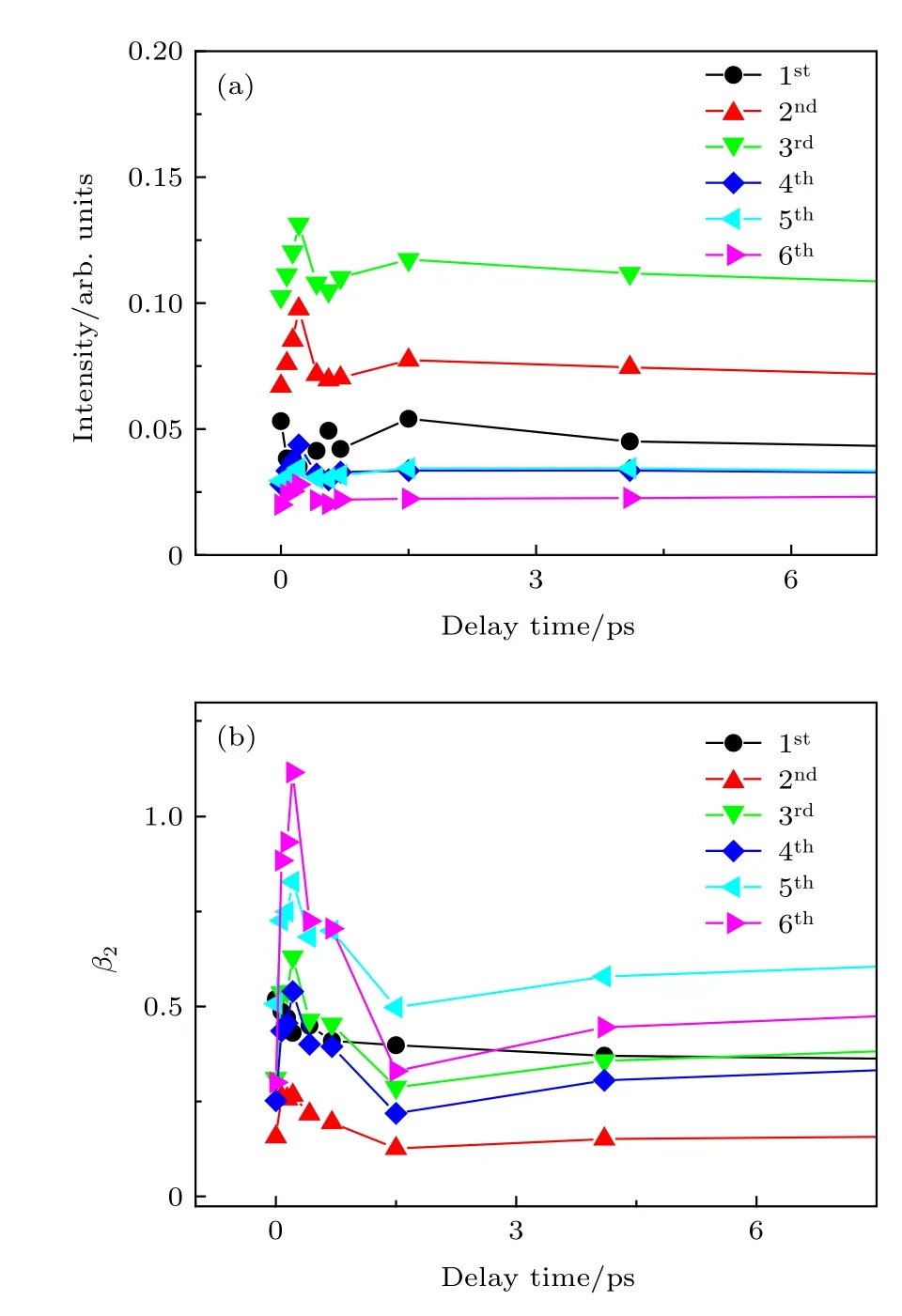
图6 (a) 0—4 ps 泵浦-探测时间延迟下不同光电子峰强度变化; (b)不同光电子峰对应的不同泵浦-探测时间延迟下各向异性参数Fig. 6. (a) The intensity changes of different photoelectronic peaks with 0–4 ps pump- probe time delay; (b) anisotropy parameters of the six rings as a function of pumpprobe time delay.
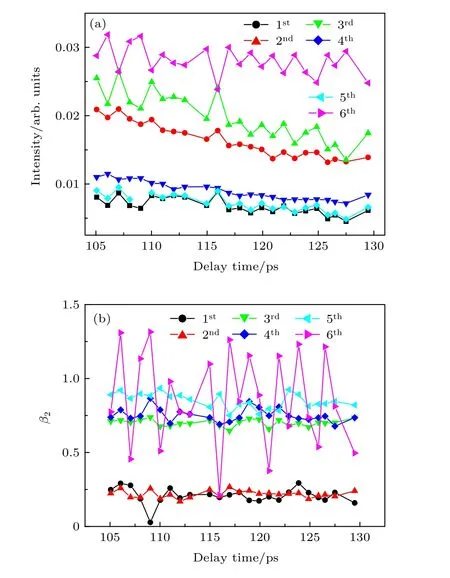
图7 (a) 105—130 ps 泵浦-探测时间延迟下不同光电子峰强度变化; (b)不同光电子峰对应不同泵浦-探测时间延迟下的各向异性参数Fig. 7. (a) The intensity changes of different photoelectronic peaks with 105–130 ps pump- probe time delay; (b) anisotropy parameters of the six rings as a function of pumpprobe time delay.
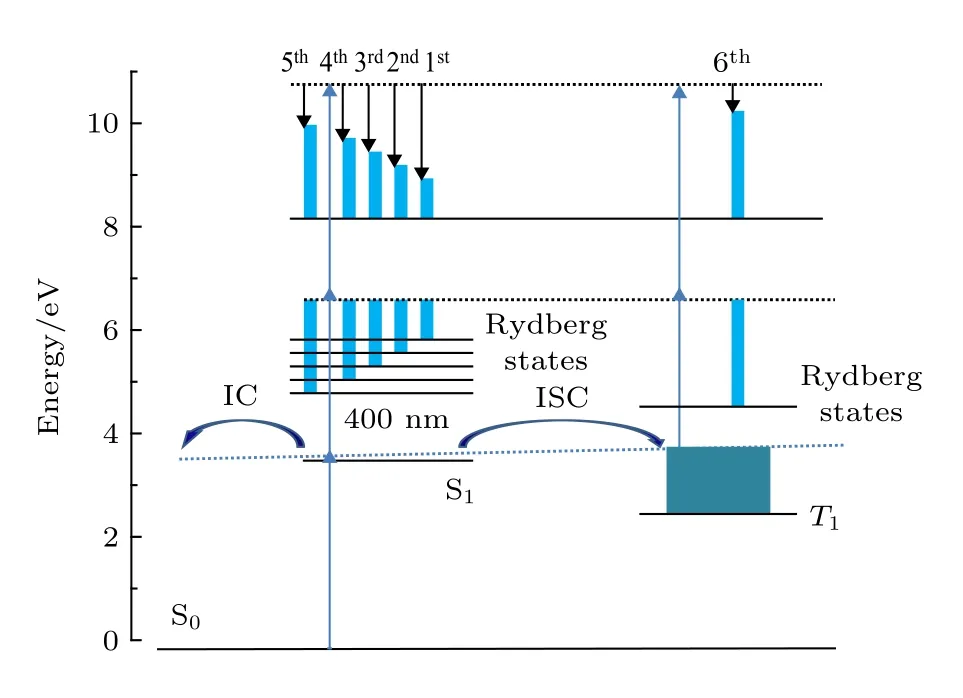
图8 2-甲基吡嗪分子被323 nm 泵浦400 nm 探测下的跃迁和电离机理示意图Fig. 8. Schematic representation of the excitation and ionization scheme of 2-methlypyrazine using 323 nm pump and 400 nm probe pulses.
4 总 结
利用飞秒时间分辨光电子影像技术和飞秒时间分辨质谱技术相结合研究了2-甲基吡嗪分子第一激发态S1态的动力学过程. 实验中观察到了2-甲基吡嗪分子丰富的非绝热动力学过程. 通过时间分辨质谱得到S1态的寿命为98 ps.时间分辨的质谱和时间分辨光电子能谱相结合观察到了S1态向三重态T1态的系间交叉过程. 实验数据表明,S1和T1态在电离的过程中和里德堡态发生偶然共振. 跟吡嗪分子S1态的动力学过程不同, 2-甲基吡嗪分子激发到S1态带源附近时, S1→ T1系间交叉和S1→ S0的内转换都是S1态的主要衰减通道. 实验中直接观察到光学暗态T1态的布局, 但由于S0态高振动态有3.84 eV 的振动能, 这个态电离的Franck–Condon 因子很小, 400 nm 探测光不能电离基态高振动态, 在本实验中未能直接观察到S1→ S0内转换过程. 实验中尝试观察2-甲基吡嗪分子第一电子激发态非绝热准直动力学, 但由于2-甲基吡嗪分子对称性比吡嗪分子更低, 对称性更低的2-甲基吡嗪分子准直现象的观察更有挑战性, 未能观察到非绝热准直现象. 本工作为2-甲基吡嗪分子S1态非绝热弛豫过程提供了较清楚的物理图像.
猜你喜欢
中国防痨杂志(2022年1期)2022-11-24
分子催化(2022年1期)2022-11-02
石油化工(2022年9期)2022-10-19
今日健康(2021年11期)2021-12-09
中国防痨杂志(2021年12期)2021-12-04
酿酒科技(2021年11期)2021-11-24
移动信息(2017年2期)2017-12-28
分析化学(2017年12期)2017-12-25
教育教学论坛(2016年49期)2017-02-27
食品工业科技(2014年6期)2014-05-10
