
温度对文蛤生理代谢的影响
2020-10-14 01:40胡高宇林兴管蔡逸龙蔡景波肖国强柴雪良
浙江农业学报 2020年9期
胡高宇,张 翔,林兴管,蔡逸龙,蔡景波,肖国强,3,柴雪良,3,*
(1.上海海洋大学 水产科学国家级实验教学示范中心,上海 201306; 2.浙江省海洋水产养殖研究所,浙江 温州 325005; 3.浙江省近岸水域生物资源开发与保护重点实验室,浙江 温州 325005)
文蛤(Meretrixpetechialis)属于软体动物门(Mollusca)瓣鳃纲(Lamellibranchia)异齿亚纲(Heterodonta)帘蛤目(Veneroida)帘蛤科(Veneridae)文蛤属(Meretrix),是我国沿海重要的滩涂经济贝类。文蛤具有较高的营养价值,有“天下第一鲜”的美誉,深受东南亚沿海人民的青睐。
呼吸耗氧、摄食排泄是贝类新陈代谢的基本生理指标,是对贝类长势进行判断的依据,也是贝类响应环境变化的评判标准之一[1]。近年来,国内贝类养殖产业发展迅速,养殖规模逐渐壮大。渔民为了提高产量,无视环境承载力,过度追求高密度养殖,致使病害滋生、水质恶化、贝类品质降低等现象频发。如何合理规划生产布局,科学布置养殖密度,是当今养殖生产中急需解决的难题,也是今后引导传统养殖业转型升级的重要指南。研究不同环境因子下不同贝类,如贻贝[2-3]、扇贝[4-5]和牡蛎[6-7]等的生理代谢响应,可为指导贝类的健康养殖提供科学依据,同时也可为贝类养殖容量、生态生理学和生理能量学等研究提供基础数据。
关于呼吸耗氧的测定,之前的研究多采用静水法[4-7]和流水法[8-9]。静水法操作简单,成本较低,至今仍被广泛使用。国内贝类耗氧率的测量方法主要采用静水法,其基本做法是将贝置于盛满海水的容器中密封,以温克勒(Winkler)碘量法测定实验前后水中溶解氧含量变化。但该方法的缺点也较明显:实验证明,静水法中溶解氧分布不均匀,测量过程容易造成水体中氧气的泄露和消耗[10];该方法只监测实验始、末的数据,因而忽视了实验过程中的参数变化;而且,静水法只能用于短期实验,对长时间贝类溶氧的监测难度较大。流水法是将贝置于开放的呼吸室内,根据进出水中溶解氧浓度差异确定耗氧率。相较于静水法,流水法解决了不能长时间实验的问题,但是流水法难以保证进水口溶解氧浓度的一致性,流速太小可能导致供氧量不足,而流速过大又会使得实验期间溶解氧浓度变化太小。同时,流水法对贝类形成的刺激也较大,容易降低滤食率效果,导致实验误差增大[11]。
间歇式呼吸测量法是一种广泛应用于水生生物中的测量技术[12-15]。该技术配备自动化程序,可实现对实验全程的数据测量,有效地减少人为操作导致的误差,便于操作;采用荧光光纤氧气测量技术测定溶解氧,测量精度更高,稳定性更强。该方法的整个测量过程分为水体交换、等待和溶氧测量3个部分。由于在测量中有水泵带动水流,因此不存在溶解氧分布不均匀的问题。同时,在交换阶段通过每次更换呼吸室内水体,排出了呼吸室内积累的代谢产物,确保了长时间实验的可能性。目前,该方法在贝类生理代谢中已得到应用。如Sui等[14]通过模拟高CO2和低氧条件,使用间歇式呼吸测量法,发现厚壳贻贝在72 h内的耗氧率显著降低;Kim等[15]采用该设备进行为期96 h的实验,探究高温与低氧共同胁迫对蛤仔的影响,发现耗氧率受到影响且显著降低。
近年来,国内外发表的关于盐度、温度、pH等环境因子对文蛤能量代谢影响的报道中,文蛤耗氧率的测定方法仍然以静水法或流水法为主[16-18]。本研究采用间歇式呼吸测量法对文蛤耗氧率进行测量,以便更加准确地反映文蛤在环境温度胁迫下的呼吸变化规律,并通过测定摄食率和排泄率,获得文蛤在不同水温下的生理响应,以期为文蛤环境容量的研究提供科学依据。同时,研究结果也将有助于文蛤养殖水平提升和养殖产量提高。
1 材料与方法
1.1 实验材料
文蛤取自浙江省乐清市翁垟街道的贝类养殖区。采集活力强、无损伤、规格整齐的个体带回实验室,用刷子清洗表面附着物后置于实验桶内暂养7 d以上,每天早、晚各投喂藻液(等鞭金藻)1次,连续充气,每天换水1次。养殖海水经沉淀砂滤后使用,海水盐度为2.5%~2.6%,水温在15 ℃左右。
1.2 实验方法
1.2.1 温度设置
根据乐清湾沿岸水温周年变化规律,共设置5个温度梯度——15、20、25、30、35 ℃。水温通过冷水机调节。实验前,文蛤在各个温度梯度里饥饿暂养24 h。
文蛤生长参数测定方法:用数显游标卡尺测壳长、壳宽、壳高(精确到0.01 mm),用Mettler Toledo精密电子天平称其总重(精确到0.01 g),并取软组织烘干测量干重(精确到0.000 1 g)。
1.2.2 摄食排泄排氨实验
按照文蛤的规格设置3个实验组(每组5只)——大规格组、中规格组、小规格组,及1个对照组(不加贝)。将每个实验组的5只文蛤放入3 L锥形瓶中,水浴控温,以文蛤贝壳张开开始计时,实验时间为2 h,实验期间全程充氧并遮光。实验过程中每组文蛤进行多次测量,取平均值用于后续计算。实验结束后,测量文蛤生长数据,解剖取软组织,60 ℃烘干并称重。收集锥形瓶内的水,测定实验前后叶绿素a浓度变化,用于计算摄食率。叶绿素a浓度参照HJ 897—2017《水质 叶绿素 a 的测定 分光光度法》,采用丙酮萃取,分光光度法测定。测定实验水体中氨氮含量,用于计算排氨率。排氨率测定参照《海洋监测规范 第4部分:海水分析》,采用靛酚蓝分光光度法测定。用300目滤网收集实验后文蛤的粪便,烘干测定颗粒态有机物(POM)值,计算排粪率。
摄食排泄排氨实验使用的文蛤生长数据见表1。
1.2.3 呼吸耗氧实验
文蛤的呼吸耗氧实验通过DAQ-M水生生物呼吸代谢测量系统(Loligo Systems公司,丹麦)测量。系统由呼吸室、水泵、溶解氧测量装置、流通池、水管等组成。呼吸室采用圆形呼吸室,体积约为1 600 mL。受呼吸室体积限制,每个呼吸室放入5个文蛤。同样按照文蛤的规格设置3个实验组和1个对照组。流通池使用加热棒进行控温,每个呼吸室各装有一个溶氧探头与主机相连。每个测量周期采用如下参数:测量360 s,水体交换200 s,等待60 s。通过连接电脑,在软件中显示溶解氧浓度变化曲线,每次测量10个周期以上,取平均值,计算耗氧率。
呼吸耗氧实验使用的文蛤生长数据见表2。

表2 呼吸耗氧实验用的文蛤生长数据Table 2 Biology data of Meretrix petechialis used in oxygen consumption test
1.3 数据分析
采用Excel 2016统计数据和作图,在SPSS 18.0软件上做单因素方差分析(one-way ANOVA)。
按以下公式计算各个温度区间内摄食率、排氨率、排粪率、耗氧率的变化情况。
RIR=V[C0-Ct-(C1-C2)]/(N×T×W)。
(1)
式(1)中:RIR为摄食率(mg·g-1·h-1),V为实验海水体积(mL),C0、Ct分别为实验组开始和结束时的饵料浓度(mg·L-1),C1、C2分别为对照组开始和结束时的饵料浓度(mg·L-1),N为实验文蛤数量,T为实验时间(h),W为软组织干重(g)。
RNR=V[(Nt-N0)-(N2-N1)]/(1 000×T×W)。
(2)
式(2)中:RNR为排氨率(μg·g-1·h-1),N0、Nt分别为实验组开始和结束时海水中的氨氮浓度(μg·L-1),N1、N2分别为对照组开始和结束时海水中的氨氮浓度(μg·L-1)。
RFER=RIR×(1-RAE);
(3)
RAE=(F-E)/[(1-E)×F]×100。
(4)
式(3)、(4)中:RFER为排粪率(μg·g-1·h-1);RAE为同化率;F为饵料中有机质的干重比例,即饵料中POM与总颗粒物含量之比;E为粪便中有机质的干重比例,即粪便中POM与总颗粒物含量之比。
ROR=V|(D0-Dt)-(D1-D2)|/(1 000×T×W)。
(5)
式(5)中:ROR为耗氧率(mg·g-1·h-1),D0、Dt分别为实验组开始和结束时海水中的氧气浓度(mg·L-1),D1、D2分别为对照组开始和结束时海水中的氧气浓度(mg·L-1)。
呼吸能(J·g-1·h-1)按照耗氧率和贝类耗氧量的能量系数计算,1 mg O2折算为14.23 J[19]。
RO/N=(ROR/16)/(RNR/14)。
(6)
式(6)中RO/N为氧氮比。
A=(Y2-Y1)/Y1×100。
(7)
式(7)中:A为相应指标(摄食率、排氨率、排粪率、耗氧率)的变化率(%),Y1、Y2分别为各个温度区间内相应指标的始、末值。
2 结果与分析
2.1 不同温度下文蛤的摄食率
如表3和图1所示,3种规格文蛤的摄食率为0.015~0.043 mg·g-1·h-1。随温度上升,同规格文蛤的摄食率总体呈先升后降的趋势。大规格文蛤在25~30 ℃区间摄食率增加最多,中规格文蛤在20~25 ℃区间摄食率增加最多,小规格文蛤在15~20 ℃区间摄食率增加最多。小规格文蛤在25 ℃摄食率达到最大值(0.037 mg·g-1·h-1),而大、中规格文蛤在30 ℃摄食率达到最大值,分别为0.043、0.035 mg·g-1·h-1。在20 ℃条件下,中规格文蛤的摄食率显著(P<0.05)低于大、小规格的文蛤,而在其他温度下不同规格文蛤的摄食率并无显著差异。

表3 不同温度区间下文蛤的摄食率变化率Table 3 Changes of ingestion rate of Meretrix petechialis under different temperature ranges %
2.2 不同温度下文蛤的排氨率
如表4和图2所示,3种规格文蛤的排氨率为1.26~135.25 μg·g-1·h-1。随温度上升,同规格文蛤的排氨率总体呈先升后降的趋势。大规格文蛤在15~20 ℃区间排氨率增加最多,中规格文蛤在20~25 ℃区间排氨率增加最多,小规格文蛤在15~20 ℃区间排氨率增加最多。小规格文蛤在20 ℃排氨率达到最大值(65.21 μg·g-1·h-1),大、中规格文蛤在30 ℃达到最大值,分别为69.15、135.25 μg·g-1·h-1。各规格文蛤的排氨率均在35 ℃最低。在15 ℃条件下,大规格文蛤的排氨率显著(P<0.05)低于小规格文蛤,但中规格文蛤的排氨率分别与大、小规格文蛤相比差异均不显著;在25 ℃条件下,3种规格文蛤的排氨率均存在显著差异(P<0.05),从高到低依次为中规格>大规格>小规格。在其他温度下,不同规格文蛤的排氨率并无显著差异。
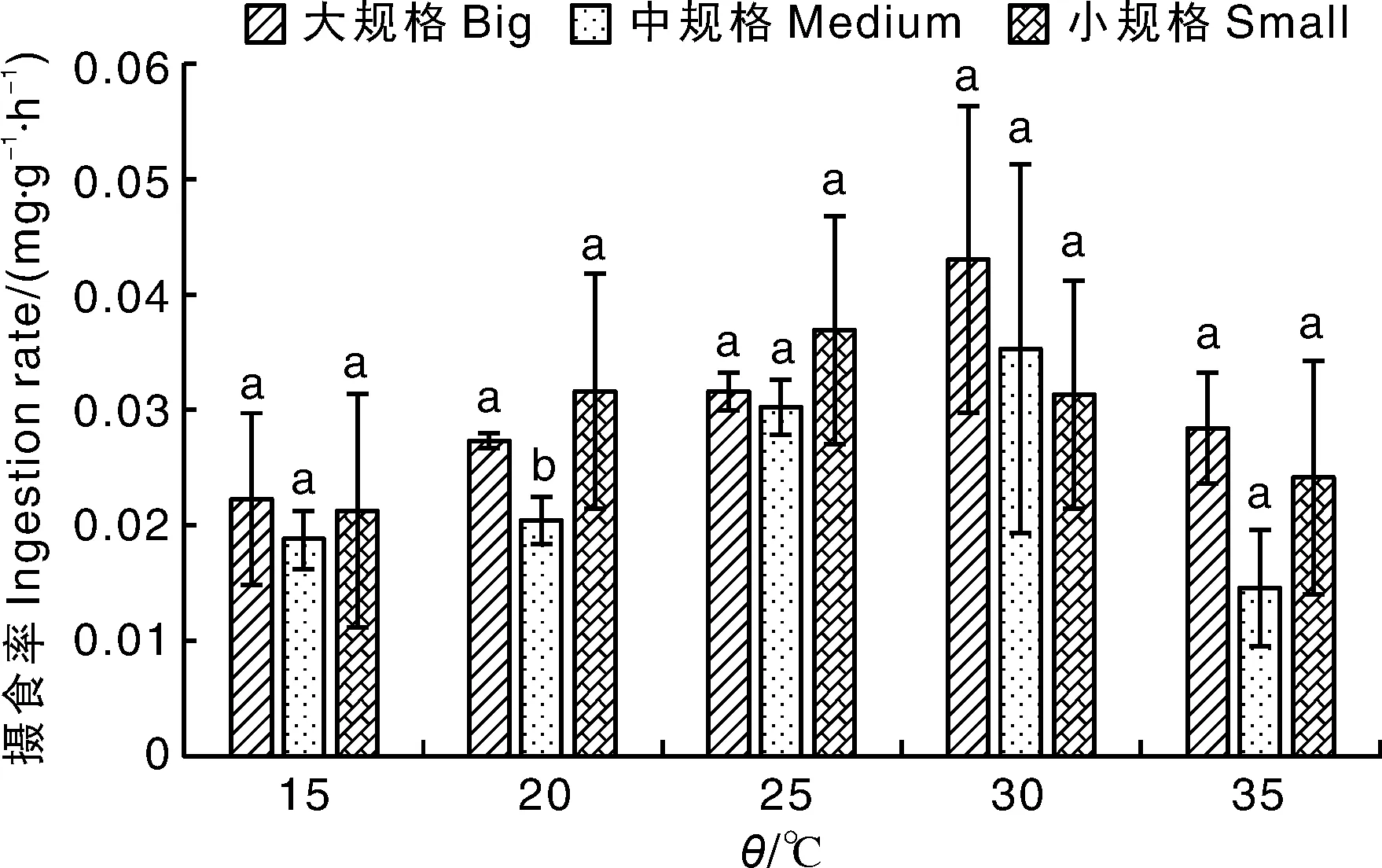
相同温度下柱上无相同字母的表示差异显著(P<0.05)。下同。Bars marked without the same letters indicated significant difference at P<0.05 under the same temperature. The same as below.图1 不同规格文蛤在不同温度下的摄食率Fig.1 Ingestion rate of Meretrix petechialis under different temperature
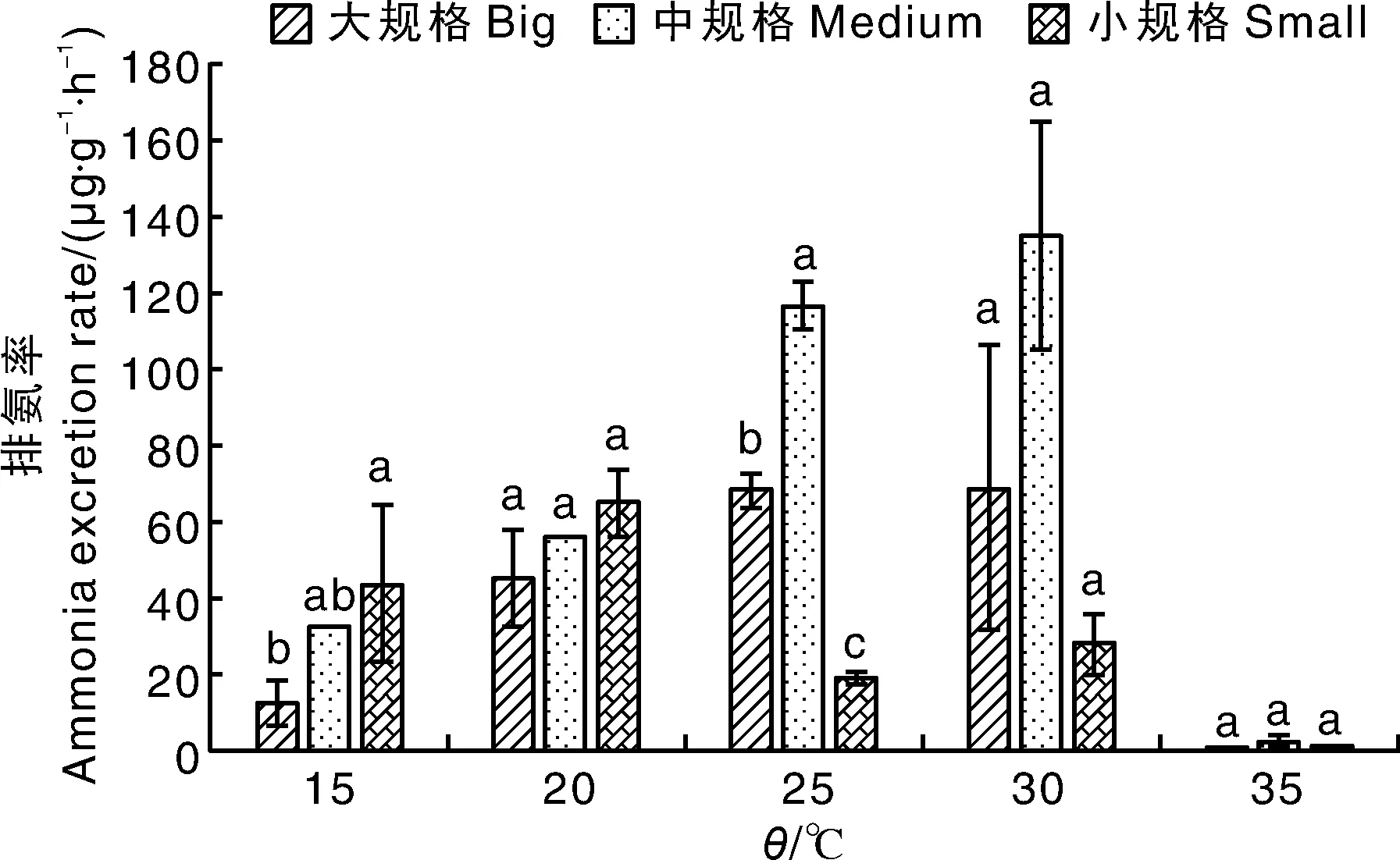
图2 不同规格文蛤在不同温度下的排氨率Fig.2 Ammonia excretion rate of Meretrix petechialis under different temperature

表4 不同温度区间下文蛤的排氨率变化率Table 4 Changes of ammonia excretion rate of Meretrix petechialis under different temperature ranges %
2.3 不同温度下文蛤的同化率和排粪率
如表5所示,在15~30 ℃,3种规格的文蛤同化率随温度升高而升高;但在30~35 ℃,大、小规格文蛤的同化率随温度升高而降低,而中规格文蛤的同化率随温度升高进一步升高。
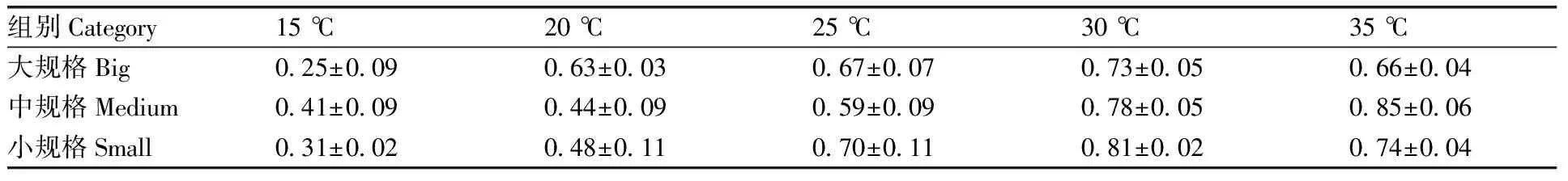
表5 不同温度区间下文蛤的同化率Table 5 Assimilation rate of Meretrix petechialis under different temperature
从表6和图4可以看出,3种规格文蛤的排粪率为2.173~15.908 μg·g-1·h-1。大规格文蛤在15~20 ℃排粪率变化最大,中规格文蛤在30~35 ℃排粪率变化最大,小规格文蛤在25~30 ℃排粪率变化最大。大、小规格的文蛤在15 ℃排粪率最大,分别为15.91、13.42 μg·g-1·h-1,中规格文蛤在20 ℃排粪率最大,为11.71 μg·g-1·h-1。在15 ℃下,大规格文蛤的排粪率显著(P<0.05)高于中规格文蛤,但小规格文蛤的排粪率分别与大、中规格文蛤相比差异均不显著;在其他温度件下,3种规格文蛤的排粪率均无显著差异。

表6 不同温度区间下文蛤的排粪率变化率Table 6 Changes of faecal egestion rate of Meretrix petechialis under different temperature ranges %
2.4 不同温度下文蛤的耗氧率和代谢效率
经测定,大、中、小规格文蛤的耗氧率均随温度升高而升高,范围为0.290~1.780 mg·g-1·h-1,且均在35 ℃达到最大值,分别为1.47、1.78、1.72 mg·g-1·h-1。如表7所示,3种规格的文蛤均在20~25 ℃区间耗氧率变化最大,但大、

表7 不同温度区间下文蛤的耗氧率变化率Table 7 Changes of oxygen consumption rate of Meretrix petechialis under different temperature ranges %

图3 不同规格文蛤在不同温度下的排粪率Fig.3 Faecal egestion rate of Meretrix petechialis under different temperature
中、小规格的文蛤分别在15~20、25~30、15~20 ℃耗氧率变化最小。从图4可以看出,3种规格文蛤的呼吸能为4.13~25.35 J·g-1·h-1。在35 ℃,3种规格文蛤的呼吸能无显著差异;在15、20 ℃,大规格文蛤的呼吸能显著(P<0.05)低于中、小规格文蛤;在25、30 ℃,中规格文蛤的呼吸能显著(P<0.05)高于大规格文蛤,但小规格文蛤的呼吸能分别与大、中规格文蛤相比差异均不显著。
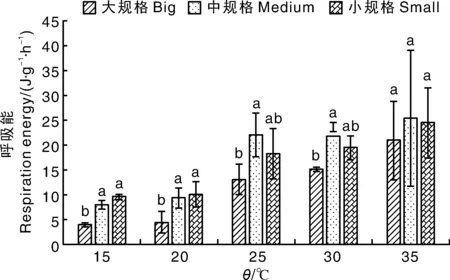
图4 不同规格文蛤在不同温度下的呼吸能Fig.4 Respiration energy of Meretrix petechialis under different temperature
如表8所示,在15~25 ℃,文蛤的氧氮比先降后升,在20 ℃达到最低值;在35 ℃,3种规格文蛤的氧氮比均剧增,远高于其他温度下。

表8 不同温度下文蛤的氧氮比Table 8 Ratio of oxygen to nitrogen of Meretrix petechialis under different temperature
3 讨论
3.1 不同温度对文蛤摄食、排泄、排氨的影响
文蛤是变温动物,外界环境温度变化对文蛤新陈代谢具有重大影响。在本研究中,随着水温升高,不同规格文蛤的摄食率先上升后下降,大、中规格文蛤在30 ℃,小规格文蛤在25 ℃,摄食率达到最大值。Jørgensen等[20]认为,在适宜的温度内,滤食性贝类摄食率随温度升高而增加。其原因一方面可能是升温导致海水黏滞性下降,加快了滤水速率;另一方面可能是温度升高导致滤食性贝类新陈代谢加快,鳃丝上纤毛摆动加速等,使得单位时间内滤水量加大。当环境温度处于适温外时,贝类生长状态会受到影响,生理代谢强度开始减弱,滤食率也逐渐下降[21]。
贝类的摄食、同化、排泄是相互影响的。van Erkom Schurink等[22]比较了4种南非贻贝(Aulacomyaater、Choromytilusmeridionalis、Mytilusgalloprovincialis、Pernaperna)的滤食、耗氧、同化等代谢指标,发现温度在10~20 ℃区间,贻贝的吸收率不受影响,但耗氧率和排泄率受影响。Wilbur等[23]将温度从15 ℃升到25 ℃,发现在贻贝(Geukensiademissa)上测得的生理参数除了吸收率外均随升温发生了显著变化。然而,董波等[24]在研究菲律宾蛤仔时发现,在9~22 ℃,其摄食率和同化率均有上升,但在26 ℃下,其生理代谢指标开始降低;栗志民等[25]在研究企鹅珍珠贝时发现,温度升高使该贝的同化率逐渐增加,并在29 ℃达到峰值,但当温度继续升高至32 ℃后,同化率降低了13.5%。本研究结果与董波等[24]和栗志民等[25]的结果有相似的变化趋势:在15~30 ℃,随着文蛤摄食率逐渐增加,饵料的同化率上升,而排粪率持续降低;在30~35 ℃,伴随着摄食率的下降,文蛤的同化率和排粪率均降低。这意味着,在适宜的环境里,水温升高通过增强贝类体内消化酶活性可以增加贝类对饵料的摄食强度,促进摄食吸收,同时减少排泄;但当水温超过贝类的适宜范围之后,酶活受到抑制,摄食同化率开始下降。关于造成不同研究结果差异性的原因:Laing等[26]认为,可能是实验温度范围选取问题,如某研究温度下实验贝类的吸收率与之无相关性;但也有可能是实验条件的不同所致,如饵料种类和浓度[27]、海水盐度和pH值等。
贝类排泄物主要有氨、尿素、尿酸等,其中氨占比达到总排泄量的70%以上[28]。排氨率是反映贝类代谢强度的重要指标之一,排氨率升高意味着,当贝类遭受到温度胁迫时,会通过提高代谢来加快能量物质消耗以应对不适条件。本研究中,温度从15 ℃升高至30 ℃时,大、中规格文蛤的排氨率均有上升趋势,而小规格文蛤的排氨率曲线处于波动状态。这可能是因为3种规格的文蛤对温度的敏感性不一样,小规格文蛤对温度变化较为敏感,体内的内环境平衡调节能力弱。
3.2 不同温度对文蛤呼吸代谢的影响
耗氧率是指单位体重的生物在单位时间内通过生物氧化所消耗的氧气量,是用来衡量生物体新陈代谢强度的基本指标之一。文蛤这类变温动物极易受到环境温度的影响,从而改变自身的代谢效率。一般认为,在正常的水温条件下,水体温度与耗氧率呈正比,一旦温度超过某一极限就会导致贝类自身生理功能出现异常,耗氧率下降。本研究中,文蛤耗氧率随温度上升而增加,在20~25 ℃增加最明显,之后增加速率逐渐减慢,因此25 ℃可能是文蛤的最适温度。该结果与在部分滤食性贝类上的结果具有相似性。刘英杰[29]发现,不同规格青蛤在20~35 ℃随温度升高耗氧率也逐渐升高;栉江珧在18~34 ℃,耗氧率随温度升高而增大,且在22~26 ℃增加率最大[30];泥蚶在15~25 ℃,耗氧率与温度成正比,在25 ℃时耗氧率达到最大值,之后随温度升高逐渐降低[31]。
在对文蛤开展的研究中,由于大部分结果是以静水法获得的,而本研究采用的是间歇式呼吸测量法,因此耗氧率的测量结果与其他研究存在差异。Bougrier等[32]认为,研究结果出现不同的原因可能在于贝类的驯化程度或者实验方法不同。范建勋[33]的结果与本研究相似,温度与耗氧率成正相关,但是其设置的最高水温仅为30 ℃,低于本研究的最高水温。王盛青等[34]和冯建彬等[35]在对文蛤的研究中发现,文蛤耗氧率随温度升高先上升后下降,但本研究中文蛤耗氧率随着温度升高并未出现下降的现象;因此,若要判断文蛤耗氧率的最高耐受温度,还需要进一步开展后续实验。
文蛤的规格对耗氧率也有较大影响。Bayne等[36]发现,贝类耗氧率随单位软体部干重的增大呈下降趋势;姜祖辉等[37]认为,毛蚶在生长过程中,脂肪和肌肉积累增多,造成重要脏器与肌肉等组织比重增大,从而导致其规格变大而代谢下降。本研究得出的结论与上述结果存在差异:在15~20 ℃升温过程中,各规格文蛤的耗氧率由大到小依次是小规格>中规格>大规格;在20~35 ℃,各规格文蛤的耗氧率由大到小依次是中规格>小规格>大规格,但是中、小规格文蛤耗氧率与温度的拟合曲线差异较小。
3.3 生物氧氮比
由于氮主要存在于蛋白质中,脂肪和糖类含氮较少,因此生物氧氮比可以用来判断生物体内营养物质的分解代谢比率。Widdows等[38]认为,氧氮比与环境胁迫有关,是判断生物体对环境适应性程度的指标之一。通常认为,当氧氮比接近7时,生物体能量需求较低,可以完全由蛋白质作为主要能源物质[39];当氧氮比约为24时,此时蛋白质供能不足,代谢物质转变为蛋白质和脂肪联合供能[40];当氧氮比超过24甚至无穷大时,碳水化合物替代蛋白质,与脂肪共同承担供能的职责[41]。本研究中,在15~30 ℃,文蛤的氧氮比值基本处于7~24,说明文蛤在能量代谢中基本上以蛋白质和脂肪供能为主,且小规格文蛤较大、中规格文蛤的变化率更大,推测小规格文蛤代谢较为剧烈,能量消耗较大。在对魁蚶(15~25 ℃)[9]、菲律宾蛤仔(9~22 ℃)[24]和墨西哥湾贝(10~31 ℃)[42]的研究中发现,其供能均以蛋白质和脂肪为主,而且环境温度升高可以提升蛋白质的利用率,降低碳水化合物和脂肪代谢率。上述结果与本研究中大、中规格文蛤(10~30 ℃)的结果类似。在35 ℃,3种规格文蛤的氧氮比均远超24,直接原因在于此时文蛤排氨率过低。可能的原因是,高温环境使文蛤酶活性降低,影响了正常的排氨机能,此时能量来源于碳水化合物和脂肪的分解。
猜你喜欢
红领巾·探索(2021年8期)2021-08-26
结直肠肛门外科(2021年1期)2021-04-22
初中生世界·七年级(2019年10期)2019-11-25
食品与生活(2019年8期)2019-10-30
食品与健康(2019年7期)2019-07-18
食品与生活(2017年11期)2017-12-18
山西果树(2015年5期)2015-11-14
小学生作文选刊·低年级版(2015年5期)2015-05-21
食品与生活(2014年8期)2014-10-20
