
橡胶树炭疽HK-36菌株α-淀粉酶基因家族生物信息学分析
2020-10-23 12:51蔡志英施玉萍戴利铭龙青姨毛常丽穆洪军
热带农业科技 2020年4期
蔡志英,施玉萍,戴利铭,高 锋,龙青姨,毛常丽,穆洪军
(云南省热带作物科学研究所,云南景洪666100)
天然橡胶是全球重要的战略性物资,也是关系我国国计民生的可再生资源,迄今为止已有100多年的栽培历史。1906年,斯里兰卡首次报道橡胶树炭疽病为害,目前该病已广泛分布于非洲、南美洲、亚洲等地的橡胶种植国[1-3]。该病害在我国早期仅在苗圃和幼树上发现,上世纪60年代初期在海南的开割胶树上首次发现少数橡胶品系整年因病落叶而导致停割。1992年,该病在海南畅好橡胶农场大面积爆发,发病面积达1 550.53 hm2,占开割胶树的75%,受害树30多万株,造成20多万株橡胶树重度落叶,损失干胶达250 t[4]。目前该病日趋严重。
炭疽菌分布广泛,其种内变异较多,寄主范围广,能引起谷类、蔬菜、果树、林木等多种作物炭疽病发生,导致严重产量损失[5-8]。该菌可侵染橡胶树的叶、花和果,严重时诱发落叶,嫩梢回枯以及橡胶树果实腐烂[4,6]。目前炭疽菌已成为研究植物寄生真菌的生长、发育、侵染过程、信号转导、寄主与微生物互作机理的模式菌株[9]。α-淀粉酶是生物体内参与淀粉水解的一类重要酶,了解橡胶树炭疽菌HK-36菌株α-淀粉酶家族基因,对深入研究该家族基因功能有着重要意义。
α-淀粉酶(α-1,4-D-葡萄糖苷水解酶)是生物体分泌的一种重要的淀粉水解酶,它水解淀粉和糖原中的α-1,4-D-糖苷键,产生短的麦芽低聚糖和麦芽糖。大多数α-淀粉酶(EC 3.2.1.1)属于糖苷水解酶(GH)家族13[10-11]。淀粉不能直接透过细胞膜,没有淀粉酶微生物就不能利用淀粉。微生物先分泌胞外α-淀粉酶内切α-1,4-糖苷键将淀粉水解成双糖和单糖,然后再摄入体内利用[12]。从微生物中提取的α-淀粉酶已大量应用于食品工业、酿造、纺织品工业和医药行业等,是应用最为广泛、最重要的工业酶之一。目前已知真菌来源的α-淀粉酶多数为曲霉属、青霉属、木霉属、根霉属和酵母属。对真菌来源的α-淀粉酶基因功能的研究多集中于工业上应用较广的曲霉属微生物。从黑曲霉基因组中发现6种α-淀粉酶,分 别 为aamA、amyA、amyB、amyC、amyD和amyE。其中amyD与amyE淀粉酶参与细胞壁α-葡聚糖的合成;amyC是一种前导酶,在异麦芽糖或麦芽糖诱导下激活AmyR(淀粉酶转录调控基因),以诱导其他淀粉酶水解酶表达,amyC的表达量非常低,目前研究相对少;aamA基因的表达产物是耐酸性α-淀粉酶水解酶,amyA和amyB两个基因的相似度非常高,与米曲霉的Taka-淀粉酶相似度大于98%[13-14]。
α-淀粉酶基因家族在炭疽菌中的分布、数量、基因结构以及其在炭疽菌中的生长发育过程中的作用知之甚少。通过对基因组的信息分析,以期望发现和解释具有普遍意义的生命现象和它们的变化、内在规律和相互关系。本文借助炭疽菌HK-36菌株基因组测序数据和生物信息学方法分析炭疽菌α-淀粉酶基因家族的种类、数量、结构特征和功能,为进一步深入研究炭疽菌中α-淀粉酶基因家族奠定基础。
1 材料和方法
1.1 α-淀粉酶家族成员鉴定
本项目组前期已对野生型橡胶树炭疽菌HK-36菌株进行全基因组测序(序列未公布),用已知真菌的α-淀粉酶氨基酸序列[15-17],利用BioEdit Sequence Alignment Editor软件比对HK-36菌株全基因组测序数据(E-Value≤1e-6),找出α-淀粉酶家族成员基因。
1.2 α-淀粉酶家族基因结构、基本特征分析
利用FGENESH在线工具,以Colletotrichum gloeosporioides、C.graminicola和C.higginsianum菌株为 参 比(http://www.softberry.com/berry.phtml?topic=fgenesh&group=programs&subgroup=gfind/)进 行 基 因结构分析[18]。使 用ProtParam(https://web.expasy.org/protparam/)预测蛋白质基本理化性质(分子式、分子量、等电点和亲水系数)。分别采用HMMTOP(http://www.enzim.hu/hmmtop/html/submit.html)、Euk-mPLoc(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)、SignalP 4.1 Server(http://www.cbs.dtu.dk/services/SignalP-4.1/)进行蛋白的跨膜螺旋[19]、亚细胞定位[20]和信号肽分析[21]。
1.3 α-淀粉酶家族蛋白系统进化分析
从NCBI中下载曲霉α-淀粉酶的氨基酸序列,用Clustal Omega软件(https://www.ebi.ac.uk/Tools/msa/clustalo/)进行氨基酸多序列比对,用MEGA 6.0软件邻位相连法(Neighbor-joining,NJ)构建系统发育树,进化树的可靠性采用Bootstrap-Method 1 000次重复检验,分析蛋白系统进化关系。
1.4 预测α-淀粉酶蛋白质家族二级与三级结构
根据同源预测和建模的原理,用在线工具PredictProtein(http://www.predictprotein.org/)蛋 白 质数据库进行α-淀粉酶蛋白质家族二级结构分析。使用工具SWISS-MODEL/SWISS-PdbViewer(http://swissmodel.expasy.org/)预测 蛋 白质 三 级结构分析。采用InterProScan(http://www.ebi.ac.uk/interpro)进行蛋白结构域的分析,应用程序选择HMMPfam、HMMSmart和SuperFamily。
1.5 α-淀粉酶蛋白质家族基因家族启动子顺式作用元件分析
用CgAmys家族成员基因开放阅读框起始密码子ATG前端的1450 bp的核酸序列,用在线软件PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析CgAmys家族成员基因的启动子区域顺式调控元件[22],并按功能进行统计分析。
2 结果与分析
2.1 α-淀粉酶蛋白质家族
用已知真菌的29个α-淀粉酶氨基酸序列,利用BioEdit Sequence Alignment Editor软件程序搜索橡胶树HK-36炭疽菌株全基因组核酸序列(EValue≤1e-6),共找出5个推定的α-淀粉酶基因,分别命名为CgAmy1~CgAmy5,基因序列已提交NCBI(登 录 号 分 别 为:MN637822、MN637823、MN637824、MN637825和MN637826)。该菌α-淀粉酶酶推导基因编码蛋白的氨基酸平均数目为529.8个,其中CgAmy2的最大的含有549个氨基酸,CgAmy3的最小含有500个氨基酸(表1)。
2.2 α-淀粉酶蛋白家族基因结构、理化性质分析及蛋白系统进化
分析橡胶树炭疽菌HK-36菌株α-淀粉酶蛋白质家族基因内含子、外显子发现,CgAmy3基因含有6个外显子,CgAmy1和CgAmy4基因外显子数量为3,CgAmy2基因含有2个外显子,CgAmy5基因仅含有1个外显子(图1)。α-淀粉酶基因编码序列长度介于1 593(CgAmy5)~1 805(CgAmy4)bp之间;蛋白分子量介于57.2(CgAmy3)~61.5(CgAmy1)kDa,平均分子量为59.48 kDa;等电点介于4.59(CgAmy4)~6.12(CgAmy1),均为亲水性蛋白,蛋白呈酸性;HK-36菌株α-淀粉酶蛋白质家族基因CgAmy1~CgAmy5都未预测到跨膜螺旋;通过在线工具Euk-mPLoc预测到CgAmy1定位于溶酶体和质膜,CgAmy2定位于内质网,CgAmy3定位于过氧物酶体,CgAmy4和CgAmy5定位于线粒 体;其 中CgAmy2、CgAmy4和CgAmy5酶 蛋 白具有信号肽(表1)。
2.3 α-淀粉酶蛋白家族系统进化分析
通过邻位相连法对曲霉属(Aspergillus niger、A.shirousamii、A.awamori、A.oryzae)踝节菌属埃默森蓝状菌(Rasamsonia emersonii)和罗克福尔青霉菌(Penicillium roqueforti)等其他23个α-amylase蛋白序列进行系统聚类分析(图2)。α-amylase蛋白被分成3个亚组,亚组I包含13个α-amylase蛋白;亚组II包含CgAmy2、CgAmy4和CgAmy5等9个αamylase蛋 白;亚 组III包 含CgAmy1和CgAmy3等6个α-amylase蛋白。
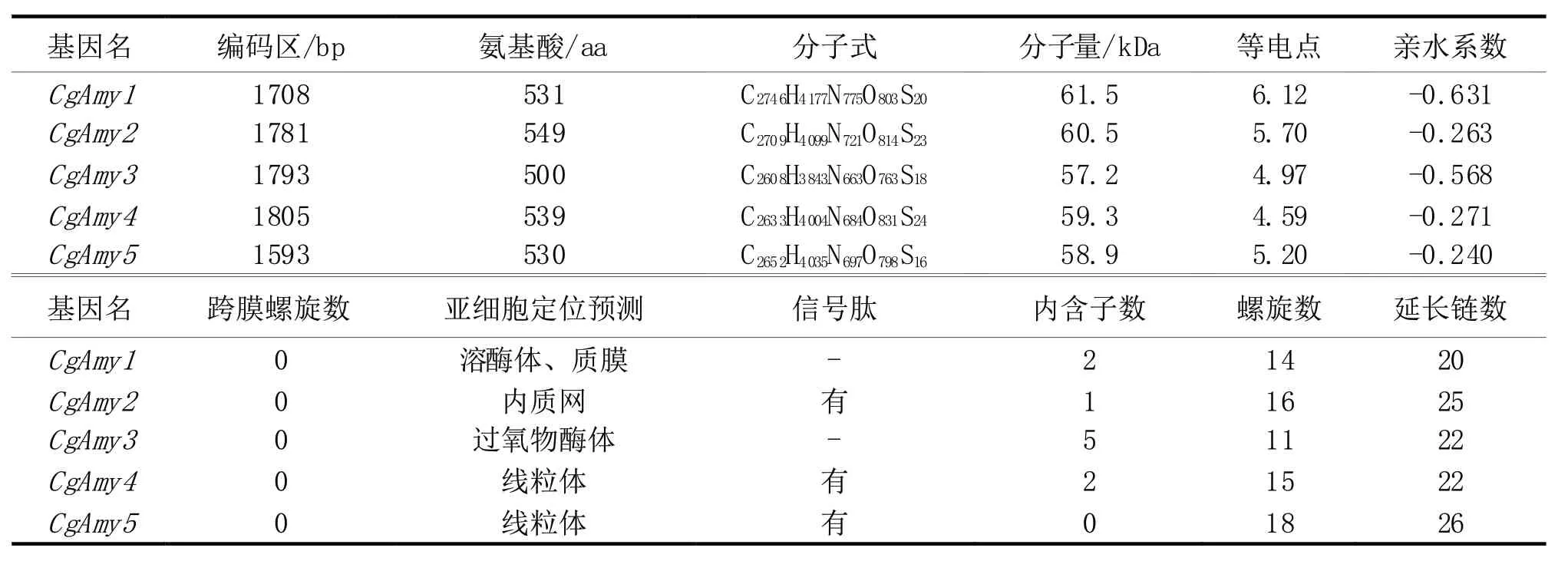
表1 CgAmy基因家族及其推导的蛋白的基本特征
2.4 α-淀粉酶蛋白质家族二级和三级结构预测
如图3-A所示,CgAmys蛋白N端由延长链和螺旋结构组成,C端基本由延长链组成。CgAmys蛋白的延长链数量在20(CgAmy1)~26(CgAmy5),其中CgAmy3和CgAmy4延长链数量均为22,CgAmy2蛋白延长链数量均为25;CgAmys蛋白的螺旋结构数量介于11(CgAmy3)~18(CgAmy5),CgAmy1、Cg-Amy2和CgAmy4蛋白的螺旋结构数量分别为14、16和15(图3-A和表1)。CgAmys三级结构也表明α-淀粉酶家族蛋白含有多个螺旋、延长链及回折结构。Swiss-Model分析结果发现,CgAmy1和CgAmy3结 构 几 乎 相 同,CgAmy2和CgAmy4结 构 相似。这一结果与CgAmys蛋白系统进化分析相吻合,CgAmy1和CgAmy3聚 类 在 一 个 亚 组,CgAmy2和CgAmy4位于同一进化枝上。
CgAmys蛋白具有同源α-淀粉酶蛋白家族指纹Glycoside hydrolase superfamily(IPR017853)和Glycosyl hydrolase,all-beta(IPR013780),同 时 含Glyco_hydro_13_cat_dom(IPR006047)结构域,含有该结构域的酶属于糖基水解酶家族13(GH 13)。α-淀粉酶是一种催化水解多糖中(1-4)-α-D-葡萄糖键的酶,在淀粉转化为麦芽糖的过程中,从糖链的非还原端去除α-麦芽糖残基。Glyco_hydro_13_cat_dom(IPR006047)的催化结构域中有一个(α/β)8桶状结构,其中包含活性位点。70氨基酸钙结合结构域位于第3个β链和第3个α螺旋之间,羧基端有个β折叠的Greek key桶状结构域。此外CgAmy4和CgAmy5还含有A_amylase_DUF1966_C(IPR015340)结构域,这种结构域存在于各种真菌α-淀粉酶蛋白中。到目前为止,它的确切功能还没有被定义。
2.5 CgAmys基因家族启动子顺式作用元件
通过CgAmys家族各成员基因的启动子元件分析结果表明(表2),除含有TATA box核心启动子元件和CAAT框等外,主要涉及激素响应和非生物逆境胁迫应答和生长调节。CgAmys基因家族各成员都具有茉莉酸应答顺式作用元件(TGACG-motif),此外还有赤霉素、水杨酸、脱落酸和生长素多种激素的响应元件,推测CgAmys对茉莉酸的响应还受其他激素共同调节,可能涉及一个较复杂的信号传递网络。
3 讨论
α-淀粉酶是一种催化水解多糖中(1-4)-α-D-葡萄糖键的酶,在淀粉转化为麦芽糖的过程中,从糖链的非还原端去除α-麦芽糖残基。目前关于α-淀粉酶基因功能研究多见于模式菌株黑曲霉(Aspergillus niger)[23-26],但是在橡胶树炭疽菌中该类基因的功能研究至今未见报道。本研究利用生物信息学方法鉴定HK-36菌株α-淀粉酶基因家族,对比分析该菌α-淀粉酶家族成员的基因结构、蛋白理化性质与结构、亚细胞定位、启动子元件,蛋白三维结构等,有助于进一步解析真菌CgAmy基因在生长发育及逆境胁迫响应中的功能。
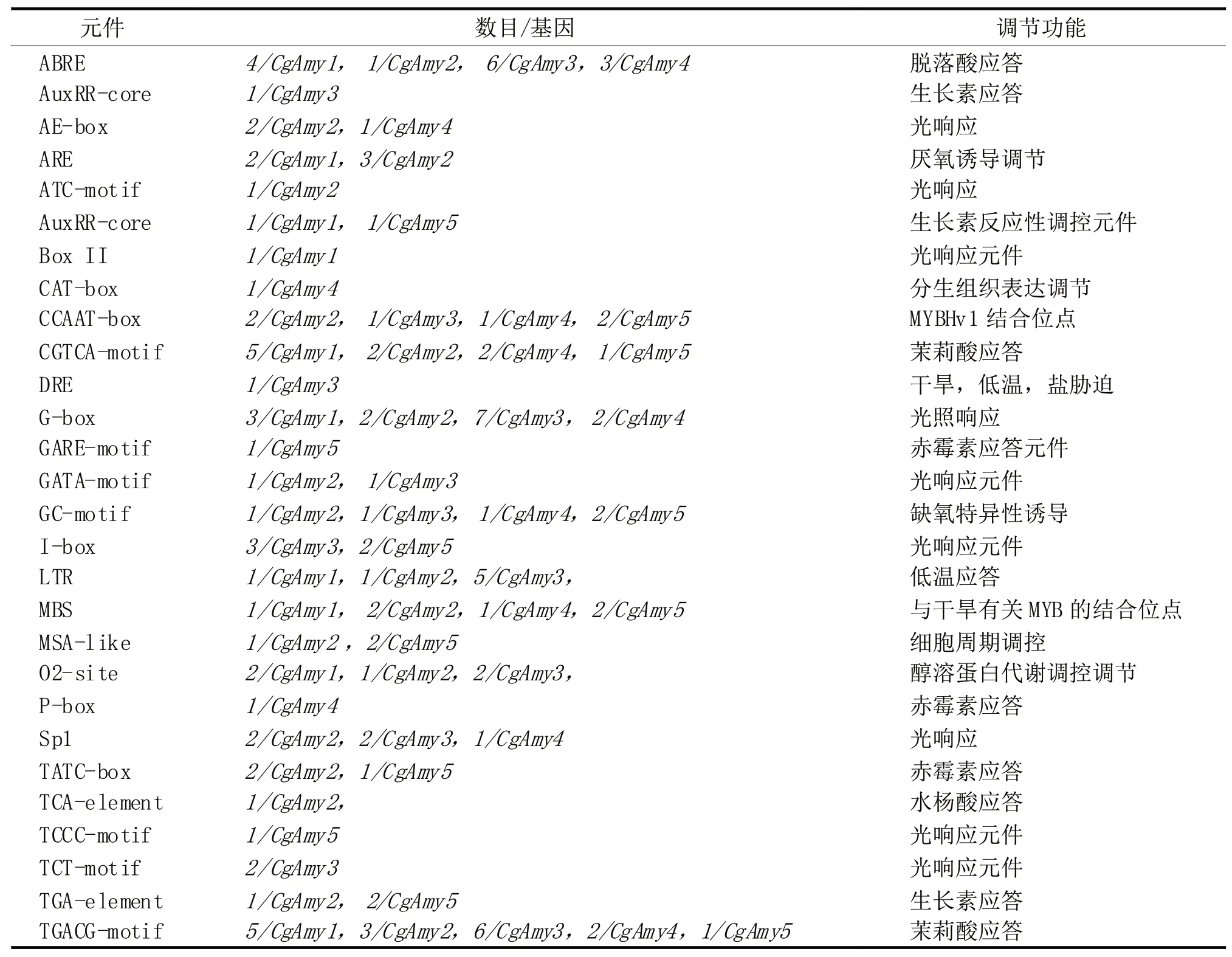
表2 Amy基因家族上游顺式作用元件
微生物α-淀粉酶通常含有3个结构域(被命名为A、B和C结 构 域)。A为α-淀粉酶催化反应中心结构域,其典型特征为(α/β)8 TIM桶 状 结 构,B和C结构域大多位于A结构域的两边。其中Ca2+的保守结合位点在A和B结构域之间,通常Ca2+对α-淀粉酶家族保持活性和稳定性是必不可少的[27]。本研究通过InterProScan分析炭疽菌HK-36菌株CgAmys家族蛋白结构域发现,CgAmys基因的推定蛋白都含Glyco_hydro_13_cat_dom(IPR006047)结构域,含有该结构域的蛋白酶属于糖基水解酶家族13(GH 13)。这个催化结构域有一个(α/β)8 TIM桶状结构,其中包含活性位点。70个氨基酸钙结合结构域位于第3个β链 和 第3个α螺 旋 之 间,羧 基 端 有 个β折 叠 的Greek key桶状结构域。这一结果与前人的研究报道相吻合,炭疽菌株HK-36的CgAmys家族的推导,它们的推导蛋白都含有(α/β)8 TIM桶状催化结构域和钙结合位点。此外,CgAmy4和CgAmy5还 含 有A_amylase_DUF1966_C(IPR015340)结 构域,暗示它们可能具有其他的功能。
基因的内显子/外含子结构数量可作为重要的进化印记,揭示家族基因各成员间的进化关系[28-29]。在 炭 疽 菌 株HK-36的CgAmys蛋 白 基 因 家族中,CgAmy5没 有 内 含 子。而其他4个CgAmys蛋白基因具有1~5个内含子,可能是在CgAmy5基础上进化而来。尤其是CgAmy3具有高达5个内含子,在进化中可能经历了多次外显子重排或内含子插入等,属于CgAmy家族基因中最新的。
系统进化分析推断炭疽菌株HK-36的5个α-淀粉酶可能来源于一个共同的祖先,但进化成二个亚组。现有研究结果表明亚组I对淀粉水解具有较高的活性[15];亚组II为糖基磷脂酰肌醇锚定α-淀粉酶,它们对淀粉的水解活性很低,但对麦芽糖和淀粉的α-葡聚糖转移酶活性很高[25];亚组III为胞内α-淀粉酶,对淀粉的水解活性相对较低,这些酶可能与细胞壁α-葡聚糖合成有关[26]。
通过CgAmys家族基因上游序列的启动子元件分析,发现该家族基因的启动子区域有不同的顺式元件,表明它们的表达很可能受多种转录因子调节。CgAmys基因家族各成员都具有茉莉酸响应元件(TGACG-motif),预示这些基因的表达可能受茉莉酸调控。此外,CgAmys基因上游序列的启动子区还具有赤霉素、水杨酸、脱落酸和生长素多种激素的响应元件,暗示CgAmys基因家族可能涉及一个较为复杂的信号传输网。关于赤霉素调控α-淀粉酶基因的表达在水稻糊粉层细胞中已经得到证实,受赤霉素刺激水稻糊粉层细胞中Ca2+-ATP酶和Ca2+通道共同调控胞质内的Ca2+浓度,以刺激α-淀粉酶基因的表达[30]。
4 结论
通过生物信息学分析,在炭疽菌株HK-36的基因组筛选到5个α-淀粉酶基因。通过氨基酸序列的多重比对分析,这5个α-淀粉酶基因的推导蛋白分别聚类在II和III亚组,亚组II包含CgAmy2、CgAmy4和CgAmy5等9个α-amylase蛋白;亚组III包含CgAmy1和CgAmy3等6个α-amylase蛋白。CgAmys基因上游序列的启动子区存在茉莉酸赤霉素、水杨酸、生长素和脱落酸多种激素的应答元件。但是,CgAmys基因所包含的顺式元件不尽相同,说明它们可能具有不同的功能。本文借助炭疽菌HK-36菌株基因组测序数据和生物信息学方法分析炭疽菌α-淀粉酶基因家族的种类、数量、结构特征和功能,为进一步深入研究炭疽菌中α-淀粉酶基因家族奠定基础。
猜你喜欢
中国合理用药探索(2022年7期)2022-08-16
蜜蜂杂志(2022年4期)2022-07-22
热带生物学报(2022年1期)2022-03-09
实用心脑肺血管病杂志(2022年2期)2022-02-22
农业科技与信息(2021年9期)2021-12-07
农业科技与信息(2021年8期)2021-12-06
中国烟草科学(2021年6期)2021-01-09
心电与循环(2020年1期)2020-02-27
科技与创新(2017年13期)2017-07-19
人人健康(2017年11期)2017-06-02
