
水母胶原蛋白的提取及性能研究
2020-12-04 06:36仇雷雷邹帅军王倩倩张黎明海军军医大学海军特色医学中心海洋生物医药与极地医学研究室上海200433
药学实践杂志 2020年6期
仇雷雷,王 博,邹帅军,王倩倩,张黎明 (海军军医大学海军特色医学中心海洋生物医药与极地医学研究室,上海200433)
0 引言
胶原蛋白主要存在于动物皮肤、骨骼和结缔组织中,是主要的结构蛋白质。胶原蛋白具有较低的免疫原性、较好的生物相容性、良好的生物降解性等生物学优点,广泛应用于生物制药、医疗保健、组织工程、食品加工、美容化妆品等领域。目前胶原蛋白主要来源于陆生牲畜(牛、羊和猪等),但某些疾病(口蹄疫、疯牛病)以及宗教相关问题,使得这些牲畜来源的胶原蛋白应用受到一定约束[1]。海洋来源的胶原蛋白资源丰富,且不存在传染性病菌和伦理等问题,因而,对海洋生物来源的胶原蛋白进行开发利用具有良好的应用前景[2]。
目前,从海洋生物提取胶原蛋白的研究主要集中在鳕鱼、罗非鱼、海参以及水母等,这些海洋生物的皮肤、肌肉、软骨组织及中胶层等部位中含有丰富的胶原蛋白[3]。水母是一类胶质状的浮游生物,属刺胞动物门,种类多,分布广,我国近海海域水母资源丰富。大型水母伞部直径可达2 m,中胶层非常厚实,其中胶原蛋白(122.64~693.92 mg/g,干重)约占总蛋白含量的50%[4]。本研究选用青岛海域采集的越前水母(Nemopilema nomurai),对其伞部胶原蛋白进行分离纯化和表征,为水母胶原蛋白的开发应用提供理论基础。
1 材料与仪器
1.1 仪器
SCIENTZ-12N 冷冻干燥机(宁波新芝生物科技股份有限公司);LA8080 氨基酸自动分析仪(日本株式会社日立高新技术科学);Nicolet iS50 傅里叶变换红外光谱仪(美国Thermo Scientific 公司);X′Pert3 Powder X 射线衍射仪(荷兰帕纳科公司);VARIOSKAN LUX 多功能酶标仪(美国Thermo Scientific 公 司);Mini-PROTEAN Tetra System 垂直电泳系统(美国BIO-RAD 公司)。
1.2 试剂
越前水母样品采集于青岛海域,腌制后于-20 ℃保存;牛I 型胶原蛋白标准品(河北考力森公司);胃蛋白酶(上海生工生物工程公司);标准蛋白Marker(日本TaKaRa 公司);其余试剂均为国产分析纯。
2 实验方法
2.1 胶原蛋白的提取与纯化
提取:经腌制的水母样品用自来水清洗,去除表面盐分,4 ℃下用去离子水浸泡、洗净后,再用异丙醇去除脂肪、NaOH 去除杂蛋白、过氧化氢去除色素。称取去杂水母伞盖样品,组织匀浆后加入相应体积不同浓度(0.3~0.6 mol/L)的乙酸溶液,胃蛋白酶的添加量为水母伞盖质量的0.1%~0.5%,酶消化时间为24~96 h,反应后的液体使用200 目网过滤,离心(4 ℃,10 000 r/min)30 min 取上清即得粗提液。
纯化:①超滤:分别取粗提液5 ml 至分子量10 万和 5 万的超滤管中,4 ℃ 离心(4 000 r/min,30 min),收集滤液进行SDS-PAGE 分析。②透析:分别取粗提液5 ml 到分子量5 万和10 万的透析袋中,4 ℃透析3 d,每8 h 换一次透析液,取样进行电泳分析。③盐析:取粗提液5 ml,添加NaCl 至浓度为 0.9 mol/L,4 ℃ 下 10 000 r/min 离心 10 min,收集沉淀,复溶于0.5 mol/L 乙酸溶液,重复上述过程2 次,取溶液进行电泳分析。仅改变NaCl 浓度(0.9 mol/L→0.9 mol/L→2.0 mol/L 或 2.0 mol/L→0.9 mol/L→0.9 mol/L),重复实验。
粗提液纯化后,在-80 ℃冷冻干燥得水母酶溶性胶原蛋白(PSC)。
2.2 单因素分析和正交实验设计
以水母胶原蛋白得率(冻干胶原蛋白质量/水母干重)为指标,确定基本反应条件(乙酸浓度、料液比、胃蛋白酶用量、提取时间),每次改变其中一个条件进行水母胶原蛋白提取的单因素实验。
获得单因素数据后,设计正交实验(表1)来获取最佳的实验方案。

表1 水母胶原蛋白提取正交实验因素水平表
2.3 天狼星红法测定胶原蛋白浓度
参照Coentro 等[5]的方法检测胶原蛋白浓度,将100 μl 胶原蛋白溶液添加至1 ml 天狼星红色染料中静置30 min,然后以12 000 r/min 离心30 min。去除上清,沉淀用碱性试剂溶解30 min,取200 μl胶原蛋白液于540 nm 处读数。
2.4 X 射线衍射
用X 射线衍射仪分析胶原蛋白样品的晶体结构,X 射线源为 Cu 靶 Ka 辐射(λ=0.154 nm),电流为 40 mA,电压为 40 kV。扫描角度 2θ 为 5°~40°,扫描速率为 3.6°/min,并根据公式 λ=2d sinθ,计算d 值。
2.5 紫外光谱分析
胶原蛋白溶解于0.5 mol/L 乙酸中,用多功能酶标仪检测其紫外光谱。扫描波段为200~400 nm,波长间隔为1 nm。以0.5 mol/L 乙酸溶液作为空白对照。
2.6 傅里叶变换红外光谱分析
取胶原蛋白5 mg 和适量KBr 置于研钵中,研磨均匀。取适量的混合样品于压片磨具中,用压片机制成试样薄片。将压片放入傅里叶变换红外光谱仪内进行扫描(500~4 000 cm-1,分辨率 2 cm-1)。
2.7 十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDSPAGE)
胶原蛋白溶液和上样缓冲液混匀后水浴(100 ℃)加热 5 min,取 15 μl 上样,使用 8% 的分离胶和5%的浓缩胶,80 V 电泳100 min。
2.8 氨基酸组成分析
胶原蛋白样品在110 ℃下盐酸水解24 h,水解产物12 000 r/min 离心20 min,上清液用氨基酸自动分析仪进行氨基酸含量测定。
本文采用MIDAS-GTSNX岩土有限元软件来模拟双排水盲沟的渗流计算。一般计算模型如图2所示。计算时,在模型两侧以及排水盲沟处设置给定水头边界;在模型底面、顶面,以及排水盲沟上的开挖面设置给定流量边界,流量为0,即为隔水边界。
2.9 胶原蛋白的溶解性
将胶原蛋白样品溶于0.5 mol/L 乙酸溶液中,配成1 mg/ml 的胶原蛋白溶液,分别取10 ml 溶液调配成不同 pH 值(3~9 共 7 个梯度)和 NaCl 浓度(0.5、0.6、0.7、0.8、0.9、1.0、1.1 mol/L),10 000 r/min离心,取上清液进行胶原蛋白浓度测定,并绘制变化曲线。
2.10 数据处理
3 结果与分析
3.1 胶原蛋白提取的单因素结果
从图1A 可见,当酶的用量从0.1%升到0.3%时,胶原蛋白得率逐渐增高并达到最大值。继续增加酶使用量,胶原蛋白得率反而减少,可能是过量酶对胶原蛋白具有一定破坏作用。从图1B 可见,当乙酸浓度为0.4 mol/L 时,胶原蛋白的得率最高。而乙酸过量,会破坏已生成胶原蛋白。从图1C可见,料液比在1∶2 g/ml 时,水母胶原蛋白的得率最大。可能是料液比低于1∶2 g/ml 时,胶原溶胀不充分,而料液比继续增加并没有多余胶原以供溶胀。从图1D 可见,随着时间推移,胶原蛋白的得率先升高再下降:48 h 和24 h 相比,增加了8.8%;72 h和 48 h 相比,增加了 15.9%;96 h 和 72 h 相比,得率反而下降,说明72 h 是胶原蛋白提取的最佳时间点。
3.2 胶原蛋白提取的正交实验
表 2 正交实验极差 R 分析中,RA>RD>RC>RB,即在胶原蛋白提取中,酶的使用量起着最关键的作用,提取时间排第二,料液比和乙酸浓度的影响比较小。综合这9 组数据,确定最优理论水平为A3B2C2D3,即乙酸(0.4 mol/L)、料液比(1∶2 g/ml)、胃蛋白酶(0.3%),提取 72 h,此时的得率为(33±0.63)%。
3.3 胶原蛋白的纯化
图2 中水母胶原蛋白粗提物(泳道8)在分子量13 万处含有大量的目标胶原蛋白,分子量10 万以下有杂条带。不同规格透析袋处理(泳道1、2)后,分子量10 万以下的杂蛋白部分去除;盐析结果显示,0.9 mol/L→0.9 mol/L→2.0 mol/L NaCl 的盐析(泳道4)可以去除大量杂蛋白,但目标蛋白损失较多,而 0.9 mol/L→0.9 mol/L→0.9 mol/L NaCl(泳道3)处理既可以保留目标蛋白又能够去除部分杂蛋白。最后超滤结果显示,分子量10 万(泳道6)的超滤管可以明显减少杂蛋白含量。
3.4 SDS-PAGE 电泳分析
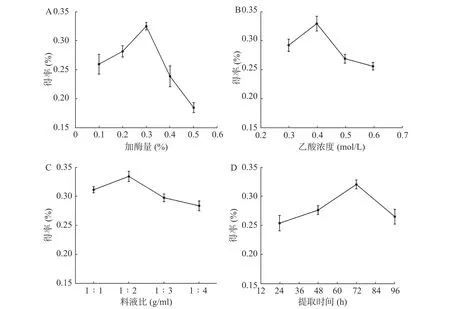
图1 水母胶原蛋白提取的单因素分析

表2 水母胶原蛋白提取正交实验结果
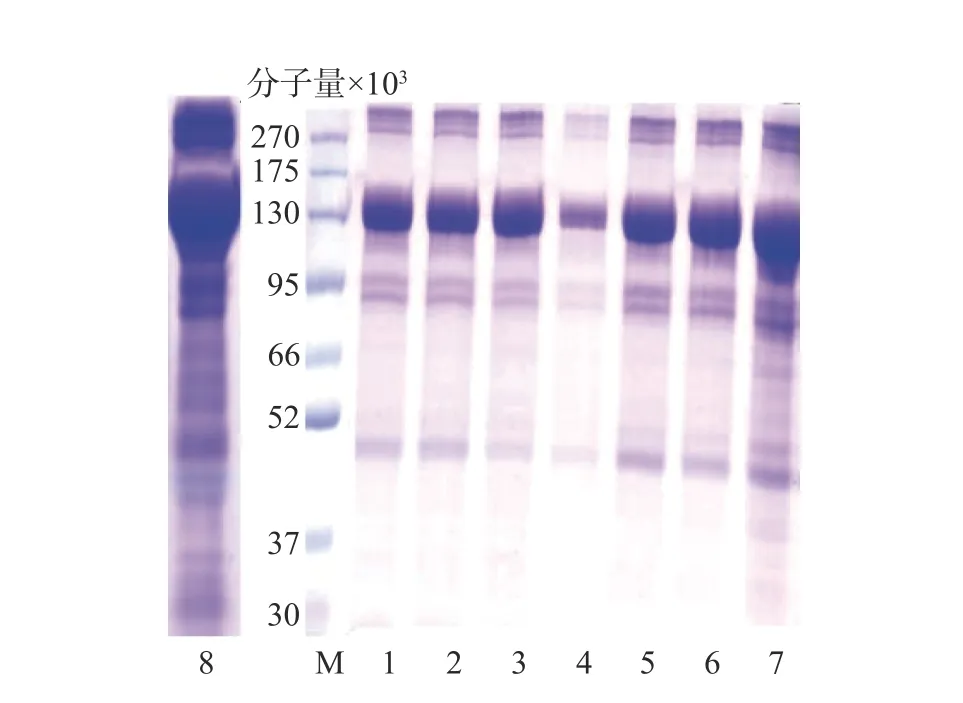
图2 水母胶原蛋白纯化过程 SDS-PAGE 电泳图
纯化实验结果提示,单一纯化方法不能完全去除杂蛋白,传统透析处理耗时长,而反复盐析会增加胶原蛋白溶液NaCl 含量。综合权衡后确定纯化条件为0.9 mol/L→0.9 mol/L→0.9 mol/L NaCl 盐析和100 kDa 超滤组合。图3 是组合纯化后胶原蛋白电泳图:泳道b 为牛I 型胶原蛋白标准品,在分子量 13 万左右有 α1、α2链,而 β 链 γ 链的分子量都在20 万以上。本实验提取的水母胶原蛋白(泳道a),分子量10 万下基本没有杂蛋白,但α 链只发现了一条,其分子可能是(α1)3[6-7],分子量在13 万左右,符合I 型胶原蛋白的特征。
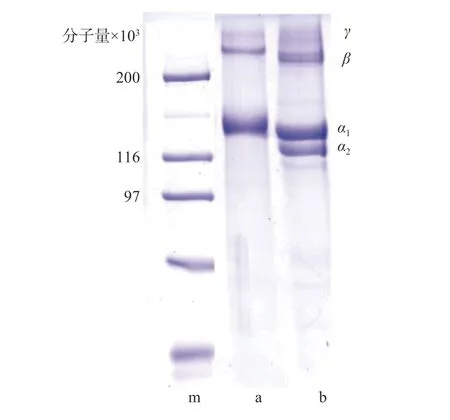
图3 纯化后水母胶原蛋白 SDS-PAGE 电泳图
3.5 胶原蛋白的X 射线衍射图
胶原蛋白X 射线衍射分析结果如图4、表3 所示。水母胶原蛋白和牛胶原蛋白都有3 个衍射峰。峰1 形状尖锐与胶原蛋白三螺旋结构有关,d 值反映胶原蛋白分子链间距离[8],水母和牛胶原蛋白的d1值分别为 1.244 nm 和 1.226 nm。在20°附近出现一个宽大峰(峰2),代表胶原蛋白纤维内部众多结构层次所引起的漫散射[9]。在30°附近出现了第3 个峰(峰3),该峰较小,其d 值显示三股螺旋结构中沿螺旋中心轴相邻氨基酸残基之间的距离[10],水母和牛胶原蛋白峰3 的d3值分别为0.295 nm 和0.296 nm,极为接近。说明水母胶原蛋白和牛胶原蛋白标准品一样,保持了完整的三股螺旋结构。
3.6 胶原蛋白的紫外吸收光谱
蛋白通常含有芳香族氨基酸酪氨酸、色氨酸和苯丙氨酸,这些氨基酸含有苯环共轭双键系统,在特定波长(280 nm 和251 nm)处有最大吸收峰。但在胶原蛋白中这些氨基酸含量少,其紫外吸收峰主要集中在220~230 nm,主要是由羰基C=O 的n→π跃迁所致。水母胶原蛋白的紫外光谱如图5 所示,其最强吸收峰在220 nm 处,与牛胶原蛋白吸收峰出现的位置类似,接近Ab Aziz 等[11]报道的水母Rhopilema hispidum 胶原蛋白紫外吸收峰(218 nm),提示本实验中提取的水母胶原蛋白纯度较高。
3.7 胶原蛋白的傅里叶变换红外光谱
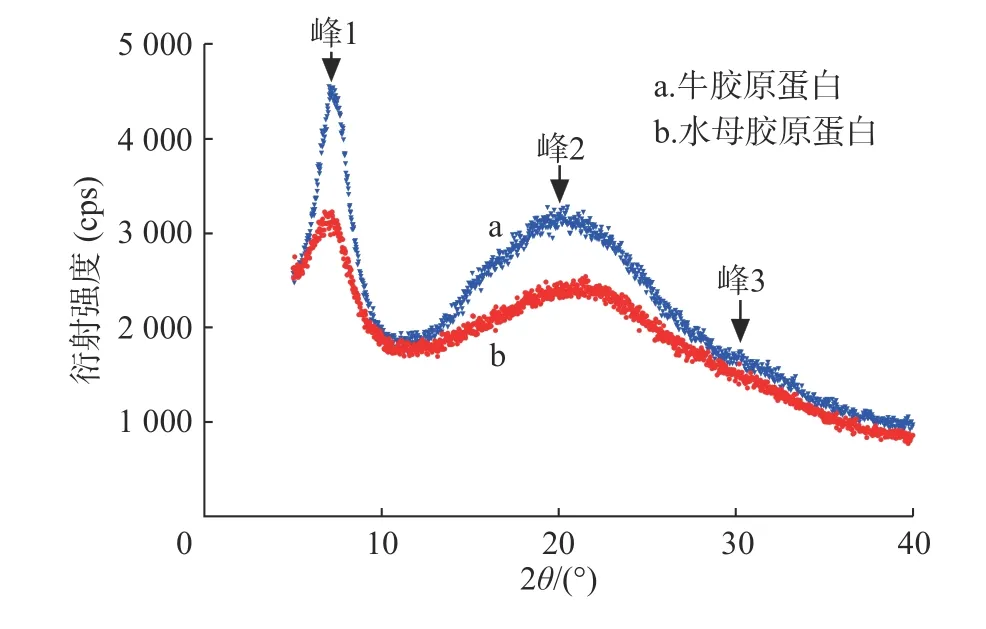
图4 胶原蛋白的 X 射线衍射图谱
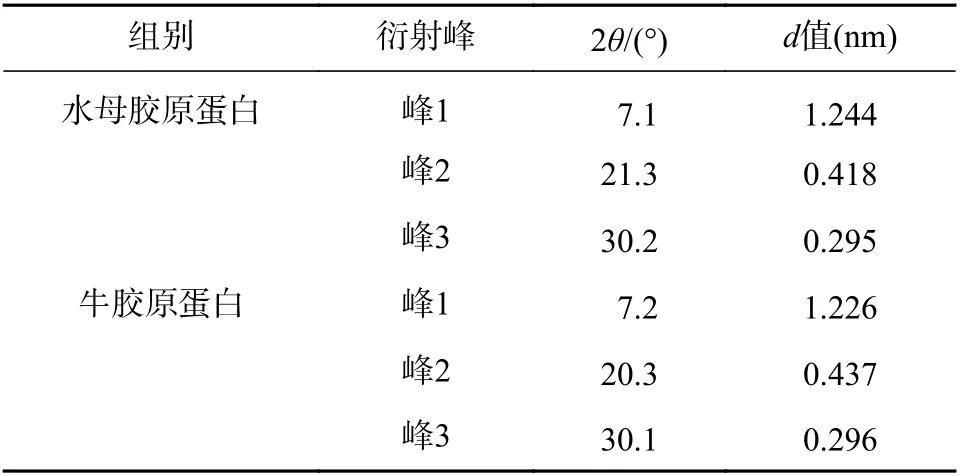
表3 胶原蛋白 X 射线衍射峰对应的 d 值
从图6 可见,水母胶原蛋白酰胺A 吸收峰为3 288 cm-1,和 Rastian 等[12]检测的酰胺 A(3 292 cm-1)吸收峰接近,表明氢键存在于胶原蛋白结构中。胶原蛋白分子中CH2基团的不对称伸缩振动产生了酰胺B 特征吸收峰[13-14],水母胶原蛋白的酰胺B 在2 933 cm-1处,和牛胶原蛋白的酰胺 B(2 936 cm-1)吸收峰类似。作为蛋白质二级结构变化的特征区域——酰胺Ⅰ带(1 625~1 690 cm-1),是由蛋白质肽链骨架的C=O 伸缩振动产生[15],水母胶原蛋白的酰胺I 带出现在1 641 cm-1。水母胶原蛋白酰胺II 带吸收峰出现在1 543 cm-1,在胶原蛋白酰胺II 带(1 500~1 600 cm-1)的范围内,可能是 C—N 键的伸缩振动和N—H 弯曲振动产生。水母胶原蛋白酰胺Ш位于1 236 cm-1处,主要是脯氨酸侧链和甘氨酸骨架的—CH2摇摆振动产生,提示所提取的水母胶原蛋白三螺旋结构完整性较好。

图6 胶原蛋白的红外光谱

图5 胶原蛋白紫外光谱
3.8 天狼星红法检测胶原蛋白
电泳图谱表明提取的水母胶原蛋白达到电泳纯(95%)。文献报道[16-18]天狼星红染料与胶原蛋白中的特征性结构Gly-X-Y 结合,可以使用天狼星红染料对胶原蛋白进行快速检测。本实验测得每100 mg 冻干的水母胶原蛋白样品中含有胶原蛋白96.85 mg,其纯度为96.85%。
3.9 氨基酸组成分析
如表4 所示,水母胶原蛋白的氨基酸组成中,甘氨酸占34.8%,与云南深水鲷鱼皮胶原蛋白甘氨酸含量(35%)接近,比热带淡水鲤鱼鳞中胶原蛋白甘氨酸含量(30.6%)高[19]。水母的胶原蛋白中羟脯氨酸占6.3%,脯氨酸占8.2%,亚氨酸比例为14.5%。其脯氨酸羟基化度(羟脯氨酸含量/亚氨酸含量)为43.68%,比牛胶原蛋白标准品的羟基化度(39.92%)和Zhang 等[7]提取的水母胶原蛋白的羟基化度(32.2%)都高。据文献报道[20],羟基化度越高,胶原蛋白的三螺旋结构越稳定。检测结果中还发现蛋氨酸、酪氨酸和组氨酸的含量比较低,缺乏色氨酸,这些都符合I 型胶原蛋白的特点[21]。
3.10 胶原蛋白的溶解性
从图7A 可见,当NaCl 浓度在0.5~0.8 mol/L时,水母胶原蛋白相对溶解度比较稳定,保持着最大值。其原因可能是低浓度的盐离子可以与胶原蛋白分子结合,使得胶原蛋白分子带有更多的电荷,相互排斥,溶解度较大。当NaCl 浓度达到0.9 mol/L时,水母胶原蛋白的溶解度急剧下降,相对溶解度不到23%,当NaCl 浓度超过1.0 mol/L 后,其相对溶解度在15%左右。可能因为NaCl 的浓度较高时,极性较强的盐离子夺走了胶原蛋白结合的水分子,破坏了其周围的水化膜,盐析作用显现出来,溶解度下降。

表4 二种胶原蛋白中不同氨基酸组分的百分比
从图7B 可见,当溶液的pH 在3~5 时,水母胶原蛋白的相对溶解度比较大,维持在80%左右。其中pH 为4 时,溶解度最大,与海参胶原蛋白的溶解性类似[3]。当pH 在6~8 时,胶原蛋白的相对溶解度维持在较低值,尤以pH 7 时溶解度最小,此时就是胶原蛋白的等电点。当pH 超过8 后,胶原蛋白溶液的溶解度又有所上升。
4 讨论
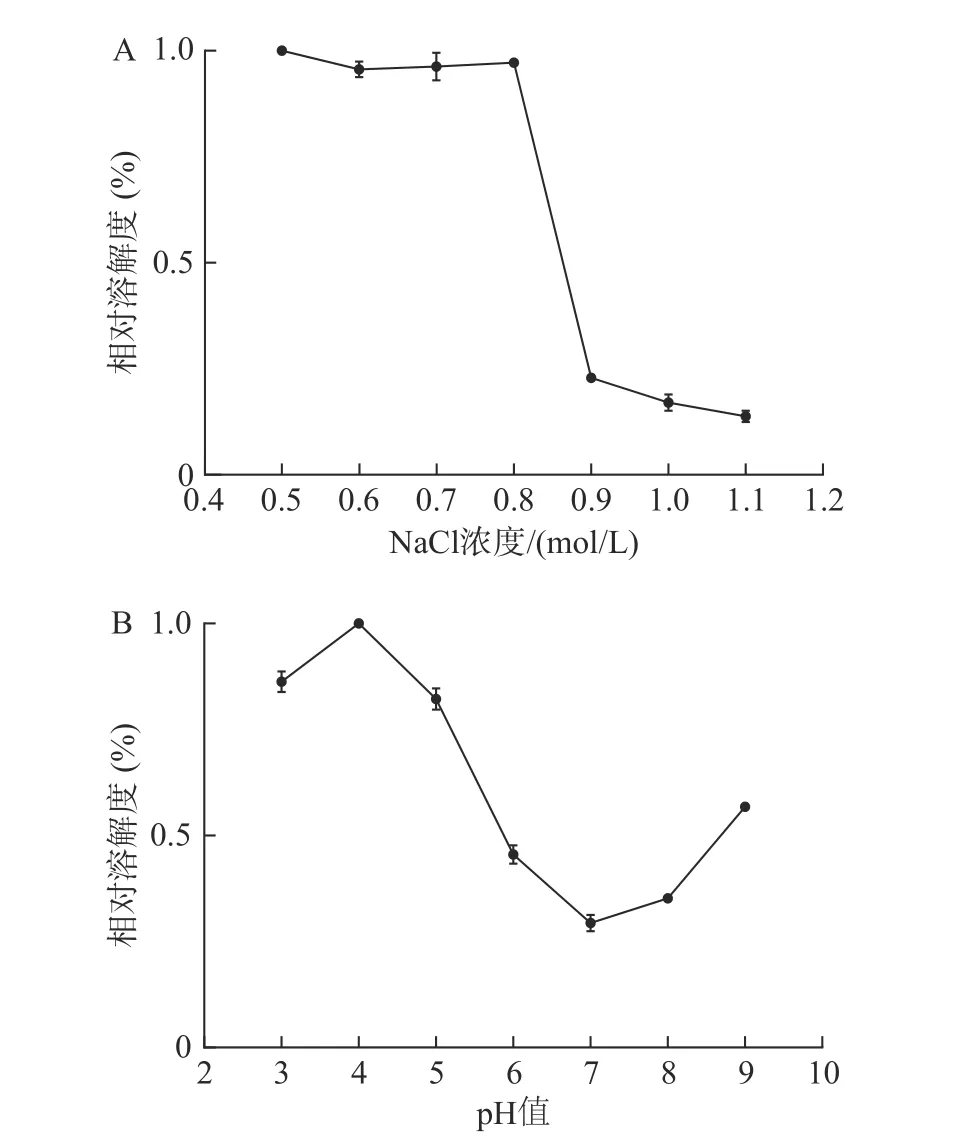
图7 水母胶原蛋白在不同 pH 值及NaCl 浓度条件下溶解度特性
本实验通过单因素分析和正交实验优化了水母胶原蛋白的提取条件,确定最适条件为:乙酸0.4 mol/L、料液比1∶2 g/mL、胃蛋白酶0.3%、反应72 h。水母胶原蛋白的纯化方法优化为0.9 mol/L-0.9 mol/L-0.9 mol/L NaCl 盐析和分子量10 万超滤的组合方法。相比阳离子交换层析法和Sephacryl S-100 凝胶过滤纯化方法,超滤不仅可以去除盐分和杂蛋白,还能快速富集胶原蛋白,缩短提取时间。
纯化的水母胶原蛋白通过天狼星红法检测其纯度为97%,高于从其他多种海洋生物提取的I 型胶原蛋白纯度[22-23]。本实验中越前水母胶原蛋白干重得率为33% 左右,相比于水母Chrysaora quinquecirrha 胶原蛋白提取得率(1.2%干重)显著提高[24]。凝胶电泳显示提取的水母胶原蛋白构型可能是 (α1)3,α1链分子量约为 13 万和 10 万以下基本没有杂条带,比Jankangram 等[25]提取的水母胶原蛋白纯度更高。在氨基酸组成分析中,越前水母胶原蛋白甘氨酸占34.82%,羟脯氨酸占6.33%,脯氨酸占8.16%,没有检测到色氨酸。以上结果均表明水母胶原蛋白符合I 型胶原蛋白的特点。光谱分析发现,分离纯化过程中水母胶原蛋白三螺旋结构保存较好。溶解度试验表明其在pH=4 时溶解性最佳,当NaCl 浓度升高至0.9 mol/L 时溶解度快速下降。
总之,本研究通过优化提取纯化过程,缩短了水母胶原蛋白制备时间,提高了所得胶原蛋白的纯度,且水母胶原蛋白三螺旋结构保持完整,与牛I 型胶原蛋白特征相似度很高,提示水母胶原蛋白可望成为哺乳动物胶原蛋白的替代品。鉴于水母胶原蛋白具有资源丰富、提取方便、生物相容性好等优点,可望成为生物医药、食品、化妆品等多领域的理想生物材料,具有良好的应用前景。
猜你喜欢
发明与创新·小学生(2022年5期)2022-05-12
动漫星空(兴趣百科)(2020年12期)2020-12-12
儿童故事画报(2020年8期)2020-10-30
世界农药(2019年3期)2019-09-10
世界农药(2019年3期)2019-09-10
试题与研究·中考化学(2016年4期)2017-03-28
中学化学(2016年10期)2017-01-07
科普童话·百科探秘(2016年10期)2016-11-23
中国学术期刊文摘(2016年8期)2016-02-13
中学化学(2015年5期)2015-07-13
