
增温增CO2对间作玉米光合特性的影响
2021-01-14 03:03王飞孙增光尹飞郭彬彬刘领焦念元
中国农业科学 2021年1期
王飞,孙增光,尹飞,郭彬彬,刘领,焦念元
河南科技大学农学院/河南省旱地农业工程技术研究中心,河南洛阳 471023
0 引言
【研究意义】受人类活动和土地利用方式的影响,大气中CO2浓度由工业革命前的280 μmol·mol-1上升到当前的 400 μmol·mol-1,并每年以 1.5—2.0 μmol·mol-1的速率继续增加。根据IPCC第五次评估报告,21世纪末大气中 CO2浓度达到 700 μmol·mol-1,气温升高2.0℃左右[1]。CO2和温度是影响植物光合作用的2个关键因子。其中,CO2作为光合暗反应的原料,不仅影响光合同化物的合成,还调节植物生长和气孔开关等;温度与植物代谢酶活性密切相关,直接影响植物光合和呼吸作用,进而影响植物光合性能和产量[2-3]。全球正面临CO2浓度升高及其带来的温室效应,给农业生产力及粮食和营养安全带来极大挑战。研究作物对CO2浓度升高和增温的响应,对于理解和预测未来全球气候变化对作物适应性的影响,以及提高农作物产量至关重要。【前人研究进展】CO2浓度升高和温度升高能够影响植物的光合作用。研究表明,CO2浓度升高能够提高植物光合效率,促进作物产量的提高[2-3]。理论上短期CO2浓度升高能够使得植物净光合速率提高35%—60%,且多数研究表明,长期的CO2浓度升高使得植物出现净光合速率促进作用下调甚至消失的趋势,即光适应现象[4-6],但也有研究报道未出现光适应现象[7]。普遍认为,温度升高会减弱CO2浓度升高所带来的正效应,降低植物净光合速率、干物质量和产量[8-9]。而在高纬度地区,增温会提高净光合速率,促进产量的形成[10]。增温增CO2对作物净光合速率所带来的促进或抑制均有报道[8,11]。AINSWORTH等[12]和 DARBAH等[13]研究发现,CO2浓度升高能够提高水稻剑叶Rubisco的最大羧化速率(Vc,max),最大电子传递速率(Jmax)和最大净光合速率(LSPn)。SAGE等[14]研究C3作物在高CO2浓度下的响应,发现作物出现光合下调现象。这可能与氮浓度降低,C-N代谢不平衡有关[15]。在玉米||花生间作复合体系中,具有明显地上、地下种间作用。地上部,间作玉米具有明显的光竞争优势,增强间作玉米Rubisco最大羧化速率(Vc,max)、最大电子传递速率(Jmax)和磷酸丙糖利用率(TPU),从而促进对 CO2的羧化固定和强光利用能力,提高净光合速率;而地下部,玉米能改善花生铁营养和竞争吸收花生根区氮,促进花生共生固氮,固定的氮能被间作玉米吸收[16];进而,间作玉米表现出显著的产量间作优势。焦念元等[17]研究发现,施磷能够提高玉米||花生体系中间作玉米羧化效率(CE)、最大电子传递速率(Jmax)和磷酸丙糖利用率(TPU),提高净光合速率。【本研究切入点】前人研究大多集中在单作体系中,在玉米||花生间作复合体系中鲜有研究。那么,CO2作为光合作用基本底物,其浓度增加是否能进一步提高间作玉米光合速率,提高对强光的利用能力,增加产量,其光合机理是什么,施磷在增加 CO2浓度后对其有什么调控效应,还有待进一步研究。【拟解决的关键问题】本研究利用开顶式气室,以玉米||花生2﹕4模式为研究对象,研究了增温增CO2对间作玉米光合色素含量、SPAD值、光合-光强、光合-CO2响应曲线及其相关参数的影响,为应对将来气候变化,实现玉米||花生绿色高产高效提供理论依据和技术指导。
1 材料与方法
1.1 试验地概况
本试验于 2018—2019年在河南科技大学试验农场进行。试验点地处温带,属于半湿润、半干旱大陆性季风气候,年平均气温 12.1—14.6℃,年平均降雨量约600 mm,年平均蒸发量约2 114 mm,年日照时数2 300—2 600 h,无霜期215—219 d,年平均辐射量约492 kJ·cm-2。试验地土壤为黄潮土,质地为中壤。0—20 cm土层基础理化性质为土壤容重1.35 g·cm-3、pH 7.66、有机质 10.74 g·kg-1、碱解氮 33.96 mg·kg-1和速效磷 6.84 mg·kg-1。
1.2 试验设计
以玉米“豫单9953”、花生“花育16”为供试材料,玉米‖花生2﹕4模式为研究对象,设置气候因子和施磷水平双因素不完全随机区组试验。施磷设 P0(0)和 P180(180 kg P2O5·hm-2)2 个水平,基施。气候因子在2018年分别设常温常CO2(环境温度和CO2,TC)和增温增 CO2(环境温度+(2±0.5)℃和(700±50)μmol CO2·mol-1,+T+C)2 个处理,共4个处理;在2019年分别设常温常CO2(环境温度和 CO2,TC)、增温常 CO2(环境温度+(2±0.5)℃,+TC)和增温增 CO2(环境温度+(2±0.5)℃和(700±50)μmol CO2·mol-1,+T+C)3 个处理,共6个处理。间作体系中,玉米宽窄行种植,宽行行距160 cm,窄行行距40 cm,株距20 cm,花生播种于宽行之中,行距30 cm,株距20 cm,玉米花生间距35 cm。南北向种植,小区宽6 m,长10 m。基施氮肥90 kg N·hm-2,于玉米大喇叭口期追施 90 kg N·hm-2,其他管理同大田生产。2018年于6月4日播种,9月22日收获;2019年于6月18日播种,10月5日收获。生长季气象数据如表1所示。
增温、增CO2均采用半封闭开顶式气室(open-topchamber,OTC),以罐装液态 CO2为 CO2气源。试验期间,采用德国LOCKE公司生产的减压阀控制增CO2气室内 CO2浓度在(700±50)μmol·mol-1,依靠气室增温效应,气室内温度比外界环境气温平均升高(2±0.5)℃。OTC为长方体钢架结构,长8 m,宽6 m,高2.5 m,面积48 m2,为减少内部气体外溢,上部框架边缘增加0.5 m宽45°收缩口,安装电风扇确保气室内空气流动、CO2浓度均匀。室壁采用阳光板,透光率达90%以上。2018年和2019年均在玉米拔节期至收获期升高CO2浓度。
1.3 测定项目与方法
1.3.1 叶绿素相对含量 使用 SPAD-502型叶绿素仪进行测定。2018年分别于苗后39 d(大喇叭口期)、58 d(灌浆期)、70 d(乳熟期)和90 d(蜡熟期)测定各处理间作玉米穗位叶SPAD值,2019年分别于苗后33 d(大喇叭口期)、55 d(灌浆期)、66 d(乳熟期)和85 d(蜡熟期)测定各处理间作玉米的SPAD值,每个小区测15片间作玉米功能叶,取其平均值。
1.3.2 光合色素含量 于2019年苗后35 d和85 d取间作玉米穗位叶,采用乙醇提取法测定叶绿素 a(Chla)、叶绿素b(Chlb)含量,并计算叶绿素a+b(Chla+b)和叶绿素a/b(Chla/b)。
1.3.3 光合-光强、光合-CO2响应曲线及其相关参数参照焦念元等[17]方法,使用 LI-6400XT型光合仪(LI-COR,美国),在2018年间作玉米苗后59 d(乳熟期)和2019年间作玉米苗后34 d(大喇叭口期)、55 d(灌浆期)、65 d(乳熟期)和82 d(蜡熟期),选择晴天 9:30—12:00,测定代表性玉米植株穗位叶的光合-光强和光合-CO2响应曲线。运用Photosyn assistant软件分析计算光饱和点(LSP)、光饱和时最大净光合速率(LSPn)、表观量子效率(AQY)、光补偿点(LCP)、羧化效率(CE)、CO2饱和时净光合速率(Amax)、Rubisco最大羧化速率(Vc,max)、最大电子传递速率(Jmax)和磷酸丙糖利用率(TPU)。

表1 生长季气象环境数据Table 1 Environmental conditions during the whole growing seasons
1.3.4 产量 在收获期,于各小区选取代表性植株2 m双行的果穗,脱粒风干后称其籽粒重量,并计算产量。
1.4 数据处理
用 Excel 2016和 SPSS 22.0软件分别进行数据整理、统计分析与作图,显著性检验采用LSD法,显著水平是0.05。
2 结果
2.1 增温增CO2对间作玉米SPAD值的影响
与常温常CO2(TC)相比,增温常CO2(+TC)后,间作玉米苗后 33和 55 d SPAD值分别提高了3.37%—8.52%和7.80%—10.96%,苗后66 d和85 d分别降低了7.33%—16.77%和15.76%—23.73%,除施磷条件下苗后33 d外均达到显著差异(P<0.05);增温(+TC)后,再升高CO2浓度(+T+C),间作玉米苗后33、55、66和85 d分别提高了3.26%—4.04%、3.22%—6.76%、18.84%—33.39%和8.46%—16.49%,除施磷条件下苗后33和55 d外均达到显著差异(P<0.05)。与TC处理相比,+T+C处理的间作玉米大喇叭口期、灌浆期和乳熟期分别提高了 4.68%—12.91%、7.88%—18.37%和 4.02%—11.43%,蜡熟期降低了8.63%—12.72%,除2019年苗后66 d外均达到显著差异水平(P<0.05)(图1)。这说明增温和增温增CO2均能促进间作玉米前期叶绿素含量,施磷表现出一定的协同效应。
2.2 增温增CO2对间作玉米叶绿素构成的影响
与常温常CO2(TC)相比,增温(+TC)后,间作玉米苗后35 d叶绿素b、a+b和a/b在不施磷条件下达到显著差异水平(P<0.05),叶绿素a和叶绿素b分别提高了 26.70%和 10.0%,叶绿素 a/b降低了17.03%;增温(+TC)后增加CO2(+T+C),叶绿素a、b和a+b在2种磷水平下均达到显著差异水平(P<0.05),分别提高了 13.74%—15.47%、26.36%—36.69%和16.95%—23.91%;与TC相比,+T+C的间作玉米叶绿素a、b和a+b分别提高17.58%—19.54%、52.55%—59.55%和26.08%—28.47%,叶绿素a/b降低了 23.04%—25.18%,均达到显著差异水平(P<0.05);增温常CO2(+TC)和增温增CO2(+T+C)间作玉米苗后85 d的叶绿素a、b和a+b均有所降低,与 TC相比,+TC的分别降低了 7.81%—8.57%、22.05%—25.50%和12.10%—14.39%,均达到显著差异水平(P<0.05);与不施磷相比,施磷条件下苗后85 d各处理叶绿素b含量均达到显著差异水平(P<0.05),TC、+TC和+T+C处理分别提高了24.15%、18.64%和22.04%(表2)。说明增温和增温增CO2能够提高生育前期叶绿素含量,同时也加速了生育后期叶绿素降解,施加磷肥能够提高叶绿素b含量,延缓叶绿素降解。
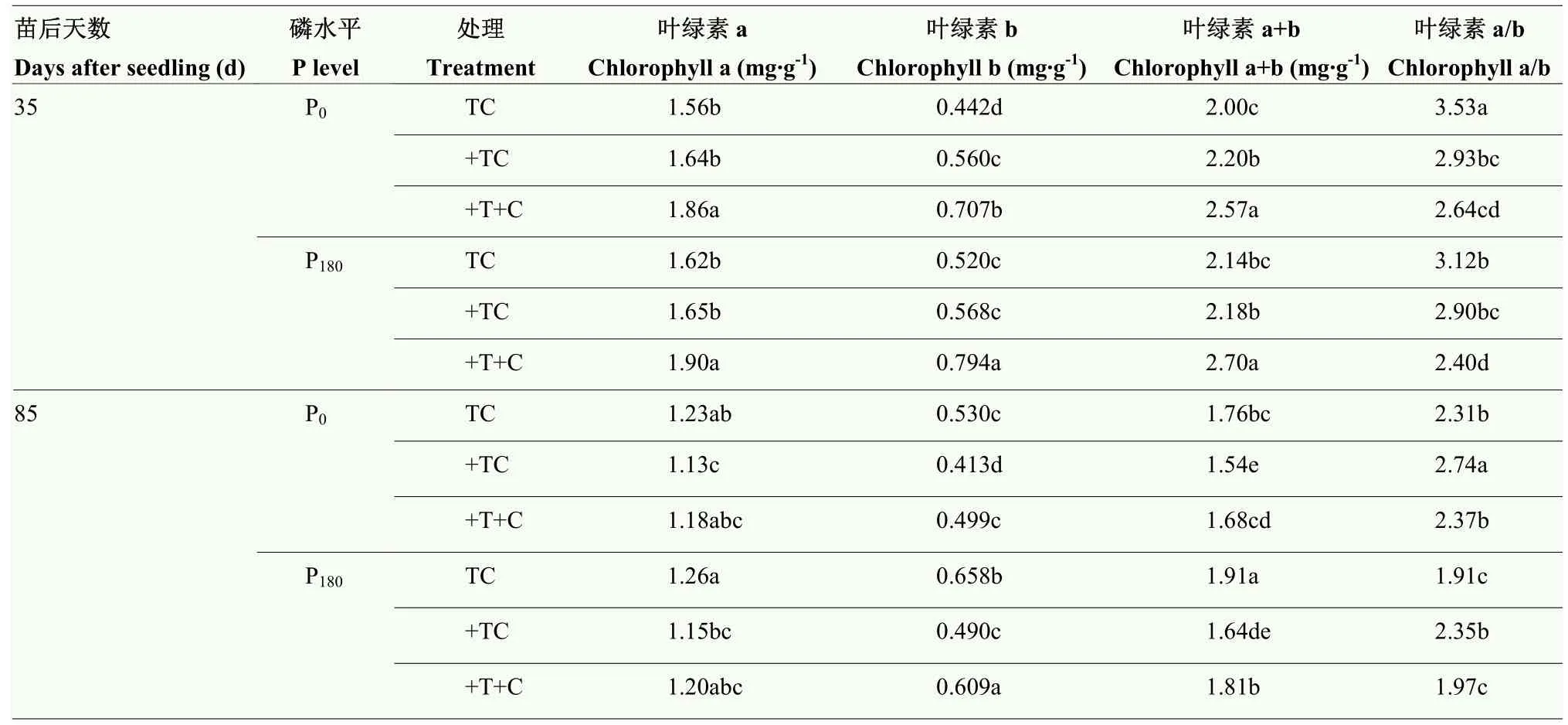
表2 增温增CO2对间作玉米叶绿素构成的影响(2019)Table 2 Effects of elevated temperature and CO2 in chlorophyll component on intercropping maize (2019)
2.3 增温增CO2对间作玉米光合-光强响应曲线的影响
玉米净光合速率随着光照强度的增强而逐渐升高,达到饱和点以后趋于平缓。在光照强度低于500 μmol·m-2·s-1时呈线性增长(图 2)。2018 年相同磷水平下间作玉米净光合速率在达到饱和点均表现为+T+C>TC。2019年间作玉米净光合速率随着苗后天数的延长逐渐降低,在苗后34 d达到最大值。苗后34、55和65 d均表现为+T+C>+TC>TC;苗后82 d表现为 TC>+T+C>+TC。与不施磷相比,施磷条件下玉米穗位叶净光合速率和光饱和点均有所升高。说明增温能够提高间作玉米大喇叭口期和灌浆期净光合速率,降低乳熟期和蜡熟期净光合速率;增温增CO2能够提高间作玉米大喇叭口期、灌浆期和乳熟期净光合速率,降低蜡熟期净光合速率,施磷能够提高各处理最大净光合速率和光饱和点。
2.4 增温增 CO2对间作玉米光合-光强响应曲线参数的影响
在生育前期(苗后 55 d),间作玉米AQY、LSPn和LSP均表现为+TC>TC,随后表现为+TC<TC,其中,苗后55 d和82 d施磷条件下的AQY达到显著差异水平(P<0.05),施磷条件下间作玉米AQY在苗后55 d提高了18.18%,在苗后82 d降低了33.33%。增温后(+TC),再增加CO2浓度(+T+C),AQY、LSPn和LSP各生育时期均表现为+T+C>+TC;LCP均表现为+T+C<+TC,其中苗后55和65 d的AQY提高了19.44%—48.72%和80.0%—82.75%,LCP降低了4.80%—21.99%和29.55%—5.47%,均达到显著差异水平(P<0.05)。与常温常 CO2(TC)相比,增温增 CO2(+T+C)AQY、LSPn和LSP在苗后35、55和65 d表现为+T+C>TC,在苗后 55 d,AQY和LSPn达到显著差异水平(P<0.05),分别提高了 30.30%—75.76%和 16.87%—19.44%。与不施磷相比,施磷条件下间作玉米苗后34 d,TC、+TC和+T+C处理的LSPn分别提高了13.30%、17.0%和9.86%,均达到显著差异水平(P<0.05)(表3)。说明增温能够提高间作玉米大喇叭口期和灌浆期的AQY、LSPn和LSP,降低乳熟期和蜡熟期的相关指标。增温增CO2能够提高间作玉米大喇叭口期、灌浆期和乳熟期的AQY、LSPn和LSP,降低蜡熟期AQY、LSPn和LSP,施磷能够提高各个生育时期AQY、LSPn、LSP和LCP(蜡熟期LCP除外)。
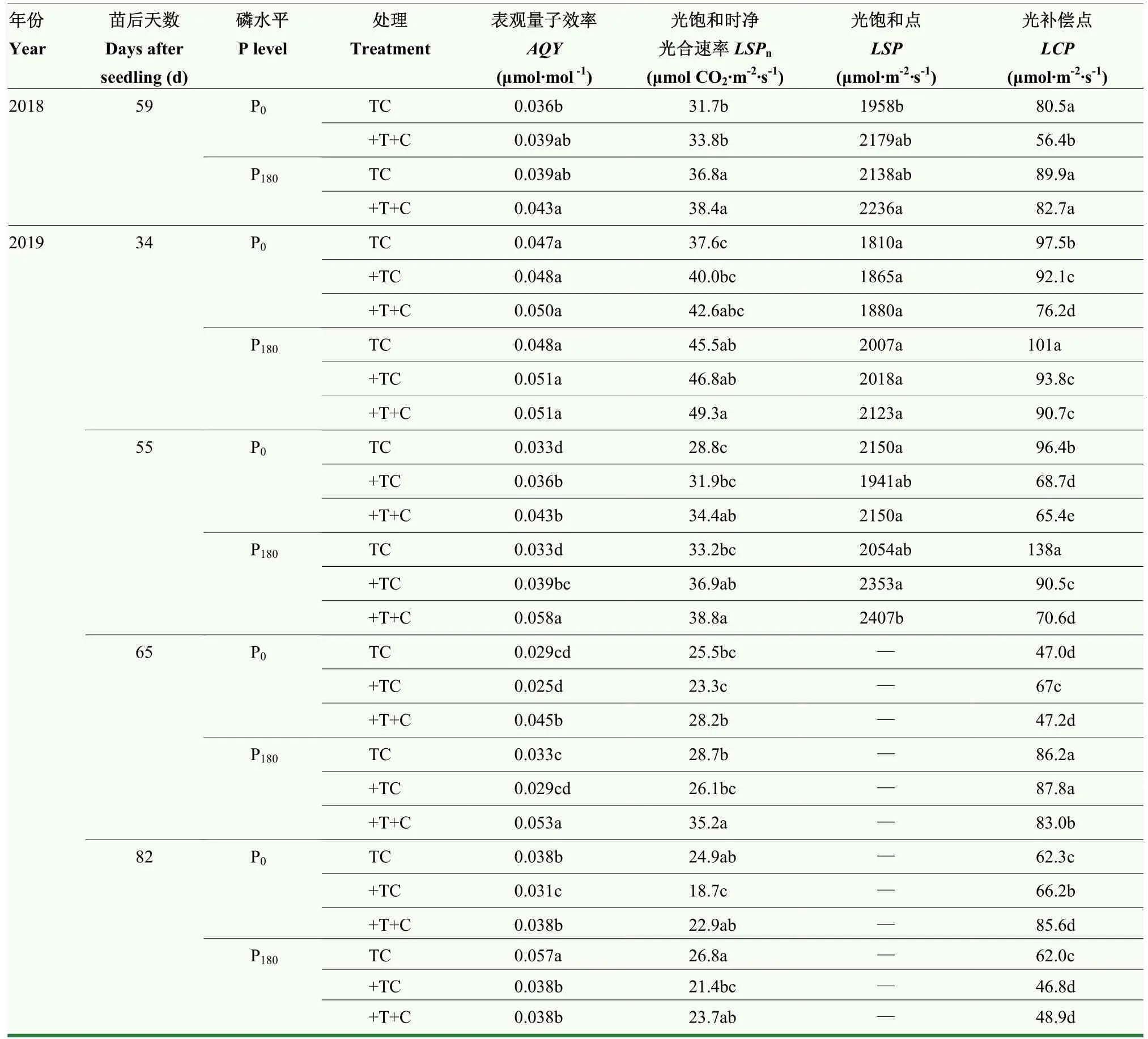
表3 增温增CO2对间作玉米光合-光强响应曲线参数的影响Table 3 Effects of elevated temperature and CO2 on the parameters of photosynthetic response curve to light intensity of intercropping maize
2.5 增温增CO2对间作玉米光合-CO2响应曲线的影响
各处理间作玉米穗位叶光合-CO2响应曲线显示,玉米净光合速率随着CO2浓度的升高而逐渐升高,达到饱和点以后趋于平缓。在光照强度低于200 μmol·m-2·s-1时呈线性增长(图 3)。2018 年相同磷水平下,间作玉米净光合速率在达到CO2饱和时均表现为+T+C>TC。2019年间作玉米净光合速率随着苗后天数的延长逐渐降低,在苗后 34 d达到最大值。苗后34、55和65 d均表现为+T+C>+TC>TC;苗后82 d表现为TC>+T+C>T+C。与不施磷相比,施磷条件下间作玉米穗位叶净光合速率和CO2饱和点均有所升高。说明增温能够提高大喇叭口期和灌浆期间作玉米 CO2饱和时净光合速率,降低乳熟期和蜡熟期净光合速率;增温增 CO2能够提高间作玉米大喇叭口期、灌浆期和乳熟期CO2饱和时净光合速率,降低蜡熟期净光合速率,施磷能够提高各处理 CO2饱和时净光合速率和 CO2饱和点。
2.6 增温增CO2对间作玉米光合-CO2响应曲线参数的影响
在间作玉米生育前期(苗后 55 d),羧化效率(CE)、CO2饱和时净光合速率(Amax)、Rubisco最大羧化速率(Vc,max)、最大电子传递速率(Jmax)和磷酸丙糖利用速率(TPU)均表现为+TC>TC,随后表现为+TC<TC,在苗后55 d施磷条件下,间作玉米Amax、Vc,max、Jmax和TPU均达到显著水平(P<0.05),分别提高了18.86%、13.34%、13.33%和20.14%。增温后(+TC),再增加CO2浓度(+T+C),CE、Amax、Vc,max、Jmax和TPU各生育时期均表现为+T+C>+TC;间作玉米苗后55 d和65 d的CE、Amax、Vc,max、Jmax和TPU均达到显著差异水平(P<0.05),其中苗后55 d分别提高了13.58%—32.96%、21.31%—11.61%、9.35%—14.55%、9.52%—15.13%和8.82%— 26.16%。增温增 CO2(+T+C)较常温常 CO2(TC),CE、Amax、Vc,max、Jmax和TPU均在苗后34、55和65 d表现为+T+C>TC,在苗后 55 d分别提高了 15.72%—36.78%、24.91%— 32.66%、20.77%—29.83%、20.93%—30.48%和27.16%— 30.74%,均达到显著差异水平(P<0.05);在苗后82 d表现为+T+C<TC,其中不施磷条件下CE和施磷条件下Vc,max达到显著水平(P<0.05),分别降低了6.67%和19.38%。与不施磷相比,施磷条件下CE、Amax、Vc,max、Jmax和TPU均有所提高,于 2018年苗后59 d均达到显著差异水平(P<0.05)(表4)。说明增温能够提高间作玉米大喇叭口期和灌浆期的CE、Amax、Vc,max、Jmax和TPU,降低乳熟期和蜡熟期的相关指标。增温增CO2能够提高间作玉米大喇叭口期、灌浆期和乳熟期的CE、Amax、Vc,max、Jmax和TPU,降低蜡熟期的相关指标,施磷能够提高各个生育时期CE、Amax、Vc,max、Jmax和TPU。
2.7 增温增CO2对间作玉米产量的影响
与常温常CO2(TC)相比,增温(+TC)后,间作玉米产量提高了19.2%—28.1%,达到显著差异水平(P<0.05);增温(+TC)后,再升高 CO2浓度(+T+C),间作玉米产量提高了5.25%—18.70%,达到显著差异水平(P<0.05)。与TC处理相比,+T+C处理的间作玉米产量提高了7.24%—52.0%,除2018年不施磷条件下外均达到显著差异水平(P<0.05)。与不施磷相比,施磷后TC、+TC和+T+C处理间作玉米分别提高了24.2%—67.2%、55.6%和27.8%—38.0%,达到显著差异水平(P<0.05)(图4)。这说明增温、增温增CO2均能提高间作玉米产量,施磷表现出一定的协同效应。
3 讨论
叶绿素是绿色植物进行光合作用的主要场所[18],而叶绿素a(Chl a)和叶绿素b(Chl b)是捕光色素复合体(LHcII)的重要组成部分,负责光能的吸收,吸收的光能转化为电能,进入电子传递链进行电子传递。周宁等[19]研究发现,增加CO2浓度明显提高了水稻生育前中期Chl a、Chl b和Chl a+b含量,降低了生育后期Chl a、Chl b和Chl a+b含量。本研究发现,增温增CO2后,间作玉米苗后35 d Chl a、Chl b和Chl a+b含量显著提高,苗后85 d Chl a、Chl b和Chl a+b含量有所降低,但未达到显著水平(除施磷条件下Chl a+b外),这与前人所得结果相一致[20-21]。本研究还发现,相比于Chl a,增温增CO2后Chl b提高幅度更大,表现更为敏感。Chl b含量虽然低,基本不影响Chl a+b总量,但可能对Chl a/b有显著影响。本研究表明,增温增 CO2后,在间作玉米生育前期 Chl a/b表现为下降趋势,在生育后期表现为上升的趋势。这与周宁等[19]研究结果一致而与谢立勇等[22]不一致。彭博等[23]研究表明,Chla/b值的下降能够提高叶片光合速率。这与本研究中间作玉米生育前期最大净光合速率上升和生育后期的下降相吻合。DONG等[24]研究发现,增温提高了水稻叶绿素含量,却降低了净光合速率,这与本研究结果不完全一致。本研究中增温提高了间作玉米生育前期叶绿素含量和最大净光合速率,生育后期增温加速了叶绿素的降解,最大净光合速率有所降低。
碳同化效率主要是Rubisco羧化效率决定的[25]。普遍认为,CO2浓度升高,提高了植物Rubisco羧化/加氧酶的羧化速率,抑制了 Rubp的氧化,从而提高了植物的净光合速率[26]。但在长期通入CO2的情况下,植物净光合速率的促进作用会出现下调甚至消失,即光合适应性现象[27]。本研究发现,增温增CO2显著提高了间作玉米生育前中期羧化效率(CE)和CO2饱和时净光合速率(Amax),降低了生育后期的CE和Amax,可能间作玉米在苗后 82 d出现适应性光合下调现象。最大羧化速率(Vc,max)、最大 RuBP再生的电子传递速率(Jmax)、最大磷酸丙糖利用率(TPU)被广泛认为是影响最大光合速率的主要限制因素[25],能反映光合作用暗反应过程的 CO2羧化固定能力。本研究中,增温增CO2提高了间作玉米前中期Vc,max、Jmax和TPU,降低了苗后82 d的Vc,max、Jmax和TPU。这与AINSWORTH等[12]和DARBAH等[13]研究结果相一致[12-13]。王建林等[28]通过盆栽试验发现玉米、水稻、花生、大豆等8种作物在苗后60 d的Vc,max均有所降低,出现光适应现象,这与本研究结果不一致。有研究指出,Vc,max的大小主要受到 Rubisco 蛋白酶的影响,光适应现象出现的主要原因可能是CO2浓度提高,加速了植物生物量的积累,从而降低了植物叶片中的N含量[29-30],导致叶片蛋白质含量整体发生下降,造成了植物单位叶面积上的Rubisco蛋白数量或活性的降低[31]。与王建林等[28]研究结果不一致原因可能在于,本研究立足于玉米花生间作体系,间作玉米可利用花生共生固定的氮作为氮源,提高间作玉米叶片中氮含量,从而保证Rubisco 蛋白酶活性和数量。LAM等[32]通过对不同固氮能力的大豆研究发现,叶片中氮含量高的品种未出现光适应性现象,这也证实了本研究的结果。磷肥是直接参与作物功能叶光合作用的光合磷酸化和碳同化过程的原料,本研究发现,施磷和增温增 CO2在间作玉米生育前中期具有明显的正效应,这可能是磷肥作为光反应电子传递链载体的重要组成部分,提高了最大电子传递速率(Jmax),促进了光反应产物ATP和NADPH的产生,从而促进了羧化效率的提高,进一步提高了净光合速率。但在间作玉米生育后期施磷和增温增 CO2具有明显负效应,可能是增温增 CO2产生的高温加速了叶绿素降解的结果。
4 结论
在玉米||花生体系中,增温和增温增 CO2能够提高间作玉米生育前期叶绿素含量和净光合速率,两者表现出明显的正向协同作用,增温降低了生育中后期叶绿素含量和净光合速率,增 CO2能够明显缓解增温在间作玉米生育中后期带来的负效应;增温增CO2提高了间作玉米生育前中期叶绿素含量、羧化效率、最大电子传递速率和磷酸丙糖利用率,从而进一步促进了对强光的利用能力,提高净光合速率。增温和增温增 CO2都促进了间作玉米产量的提高,施磷具有明显的正效应。
猜你喜欢
农业工程学报(2022年4期)2022-04-24
北方果树(2022年1期)2022-01-21
河北农业大学学报(2021年3期)2021-07-24
阅读(科学探秘)(2020年8期)2020-11-06
农业与技术(2020年12期)2020-07-04
绿色科技(2019年2期)2019-05-21
新课程·下旬(2017年7期)2017-08-14
女性天地(2016年10期)2017-04-25
新农业(2016年11期)2016-12-27
食品工业科技(2014年6期)2014-05-10
