
两个乌桕新品种苗木光合特性比较
2021-02-02 07:11许中秋隋德宗谢寅峰王俊毅
南京林业大学学报(自然科学版) 2021年1期
许中秋,隋德宗,谢寅峰*,王俊毅
(1.南京林业大学,南方现代林业协同创新中心, 南京林业大学生物与环境学院, 江苏 南京 210037;2.江苏省林业科学研究院, 江苏 南京 210014)
乌桕(Triadicasebifera)别名腊子树(浙江温州)、桕子树(四川)、木子树(湖北兴山、江西武宁),为大戟科(Euphorbiaceae)乌桕属(Triadica)落叶乔木,是中国特有的经济树种,生于旷野、塘边或疏林中,主要分布在我国黄河以南各省区,北达陕西、甘肃[1]。乌桕是集油用、材用、药用和观赏等多种价值于一身的多用树种,是我国亚热带重要的木本油料树种,其种皮可榨取桕脂(皮油),种仁可提取青油[2],总出油率可达41%以上,榨取的桕脂和青油是重要的工业原料[3]。其材坚硬且白,纹理致密清晰,是雕刻、家具和造船的良材[4]。乌桕还具有广泛的药用价值,根、树皮、叶皆可入药,用于治疗毒蛇咬伤、痈疮和通便等[5-6]。乌桕树形优美,叶形秀丽,秋叶深红、紫红或杏黄,具有较高的观赏价值[7]。目前,关于乌桕的研究报道现主要集中在化学成分分析、食品工艺、变色生理机制和育苗造林等方面[8-11]。
‘海滨绯红’和‘海滨紫晶’为近年江苏省林业科学研究院所选育的两个乌桕新品种,两个新品种当年生枝均为灰绿色,春季叶绿色,‘海滨绯红’秋叶鲜红,‘海滨紫晶’秋叶呈现紫红色,是观赏价值较高的彩叶类新品种,并已获得新品种授权保护。与现有市场常规应用品种相比,两个新品种均表现出较强的彩叶观赏特性。与彩叶品种‘红翡翠’相比,新品种叶色变化更为丰富,且现出较好的遗传稳定性,但有关其生长与生理适应性的研究鲜见报道。光合作用作为植物生长的生理基础,是评判植物生长适应性的重要指标[12]。其中,植物光合日变化是直接反应植物光合作用日生产能力的重要指标[13]。光照和CO2是植物光合作用的物质基础、能量能源和主要底物[14],研究植物光合作用对光合有效辐射(PAR)和胞间CO2浓度(Ci)的响应特征对研究植物光合能力、光合潜力及生态适应性具有重要意义[15]。鉴于此,本研究以2个乌桕新品种为研究对象,通过光合色素含量、光合日变化、光响应和CO2响应曲线的测定和比较,探讨2个乌桕新品种夏季光合性能及其差异机制,为乌桕新品种的栽培与推广应用提供理论依据。
1 材料与方法
1.1 试验材料
试验地点在南京市江宁区东善桥江苏省林业科学研究院(118°31′ E,30°38′ N),属亚热带季风气候,雨量充沛,年降水量1 034~1 276 mm,年平均温度15~15.9 ℃,年日照时间2 212.8 h,相对湿度76%,无霜期237 d。
供试材料为已获得新品种保护授权的乌桕新品种‘海滨绯红’和‘海滨紫晶’的2年生嫁接苗,砧木为同一家系实生乌桕苗。室外种植于直径40 cm、高50 cm的花盆中,每盆1株,常规水肥管理。每品种挑选6盆生长良好、无病虫害且长势相对一致的苗木,于2019年8月进行相关测定。
1.2 试验方法
1.2.1 光合色素含量测定
叶绿素含量的测定采用乙醇浸提法[16]。新鲜叶片去除中脉,剪碎混匀,称取0.2 g,重复3次,放入10 mL的试管中。加入10 mL 95%的乙醇,用保鲜膜封口,避光放置24 h,直到叶片变白。取上清液,测定波长665、649、470 nm处的吸光值,参照改良的Arnon方法[17]计算叶绿素a (Chl a)、叶绿素b (Chl b)和类胡萝卜素 (Car)的含量。
1.2.2 光合日变化的测定
利用Li-6400便携式光合测定仪(LI-COR公司,美国)于2019年8月中旬选择天气晴朗的白天,测定光合作用日变化。分别选取当年新梢相同部位的成熟叶进行测量,并做好标记。从6:00—18:00,每隔2 h测定1次,每个品种重复6次,测定的参数包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)及环境因子中的光合有效辐射(PAR)、大气CO2浓度(Ca)、气温(Ta)、空气相对湿度(φ)等。
1.2.3 光响应曲线测定
利用Li-6400便携式光合测定仪于2019年8月中旬选择晴朗天气的9:00—11:00进行光响应曲线测定,分别选取3株标准株,从植株上部的第8~10片叶进行测定。参比室CO2浓度设定为400 μmol/mol,温度为25 ℃,设定光合有效辐射(PAR)分别为2 000、1 600、1 200、1 000、800、600、400、300、200、100、80、50、20和0 μmol/(m2·s),采用自动测量模式,重复测量3次,取平均值。光响应曲线根据叶子飘新模型[18-19]进行拟合,模型的数学表达式为:
式中:在光响应曲线拟合中,Pn为净光合速率;I为光强;α为I=0时,光响应曲线的初始斜率即初始量子效率;β为修正系数;γ=α/Pn,max,Pn,max为最大净光合速率。根据公式可以得出相应的表观量子效率(AQY)、暗呼吸速率(Rd)、最大净光合速率(Pn,max)、光饱和点(LSP)、光补偿点(LCP)及模拟方程的决定系数(R2)等指标。
1.2.4 CO2响应曲线测定
在进行CO2响应曲线测定时,叶片选取与测定光响应相同的部位,采用Li-6400便携式光合测定仪的红蓝光源进行光强设定,设置光强为1 400 μmol/(m2·s),用CO2钢瓶将CO2浓度设定为50、100、200、300、400、600、800、1 000、1 200、1 500、1 800 μmol/mol,测定不同CO2浓度时的净光合速率。在设定测量程序时,为保证测定结果的准确性,每次变换CO2浓度后都进行仪器的匹配。CO2响应曲线也根据叶子飘新模型[18-19]进行拟合,模型的数学表达式为:
在CO2响应曲线拟合中,Ci为CO2浓度,Rp为光呼吸速率,其他参数涵义同光响应曲线。根据公式可以得出相应的最大光合能力(An,max)、羧化效率(CE)、CO2饱和点(CSP)、CO2补偿点(CCP)、光呼吸速率(Rp)及模拟方程的决定系数(R2)等指标。
1.2.5 数据分析
采用DPS 7.05软件对数据进行统计分析,采用多重比较Duncan新复极差法进行方差显著性检验(α=0.05)。利用Excel 2013软件作图。
2 结果与分析
2.1 两个乌桕新品种叶片光合色素含量
两个乌桕新品种的叶片光合色素含量如表1所示,‘海滨紫晶’的叶绿素a、叶绿素b含量、叶绿素a/b以及类胡萝卜素含量均高于‘海滨绯红’,其中叶绿素a、叶绿素b和类胡萝卜素含量的差异显著(P<0.05)。
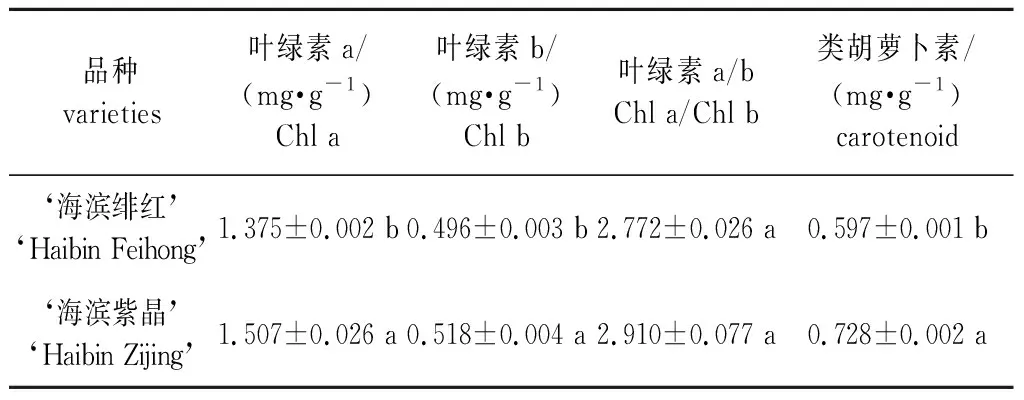
表1 两个乌桕新品种叶片光合色素含量
2.2 环境因子日变化特征
从环境因子日变化(图1)可知,测定当天的光合有效辐射(PAR)和大气温度(Ta)变化呈先上升后下降的趋势,大气CO2浓度(Ca)和相对湿度(φ)总体呈先下降后有所升高的趋势。PAR在10:00达到峰值,为1 299 μmol/(m2·s),然后下降,到18:00降至100.31 μmol/(m2·s);Ta在6:00—12:00间由27.25 ℃升至39.93 ℃,之后逐渐下降,18:00降至31.18 ℃;Ca值在6:00时最大,为439.09 μmol/mol,之后降低并上下波动;φ值在6:00时最大,为81.13%,在12:00降至最低值43.69%,之后逐渐升高,18:00时升至65.43%。
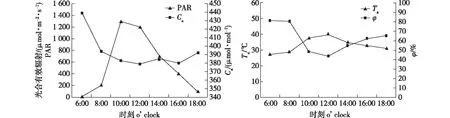
图1 环境因子光合有效辐射、大气CO2浓度、大气温度和大气湿度日变化Fig.1 Diurnal variations of photosynthetic effective radiation (PAR), atmospheric CO2 concentration (Ca), atmospheric temperature (Ta) and atmospheric humidity (φ)
2.3 不同品种光合气体交换参数日变化
两个新品种的净光合速率(Pn)日变化均呈典型的单峰型曲线(图2A、2B),无“午休”现象,两者的峰值皆出现在12:00,‘海滨紫晶’的Pn日均值[6.60 μmol/(m2·s)]高于‘海滨绯红’[5.39 μmol/(m2·s)],比‘海滨绯红’高22.45%;两者的气孔导度(Gs)日变化趋势及峰值出现的时间与净光合速率基本一致,‘海滨紫晶’的Gs日均值[0.16 mol/(m2·s)]比‘海滨绯红’[0.12 mol/(m2·s)]高33.33%。
如图2C、2D所示,两个新品种的CO2浓度(Ci)日变化与Pn、Gs的日变化趋势相反,呈先下降后上升的趋势,谷值出现在上午10:00,之后逐渐回升,‘海滨绯红’Ci日均值(335.38 μmol/mol)高于‘海滨紫晶’(305.84 μmol/mol),比‘海滨紫晶’高9.66%。蒸腾速率(Tr)日变化趋势与Pn、Gs日变化相似,‘海滨紫晶’Tr日均值[3.62 mmol/(m2·s)]比‘海滨绯红’[2.83 mmol/(m2·s)]高27.92%。

图2 两个乌桕新品种光合气体交换参数的日变化Fig.2 Diurnal variations in photosynthetic gas exchange parameters of T. sebifera in two new varieties
2.4 不同品种净光合速率与相关生理生态因子的灰色关联分析
为了进一步探讨内外环境因素对乌桕光合作用的影响程度,对两个乌桕新品种日变化过程中的净光合速率(Pn)和相关生理生态因子进行灰色关联分析,结果表明,Pn与生理因子的关联度为Tr>Gs>Ci;与环境因子的灰色关联度为Ta>PAR>Ca>φ(表2)。Ta和PAR是影响这两个品种净光合速率的主要环境因子,与‘海滨绯红’相比,‘海滨紫晶’与Tr关联度更大,而与Ca的关联度相对较小。
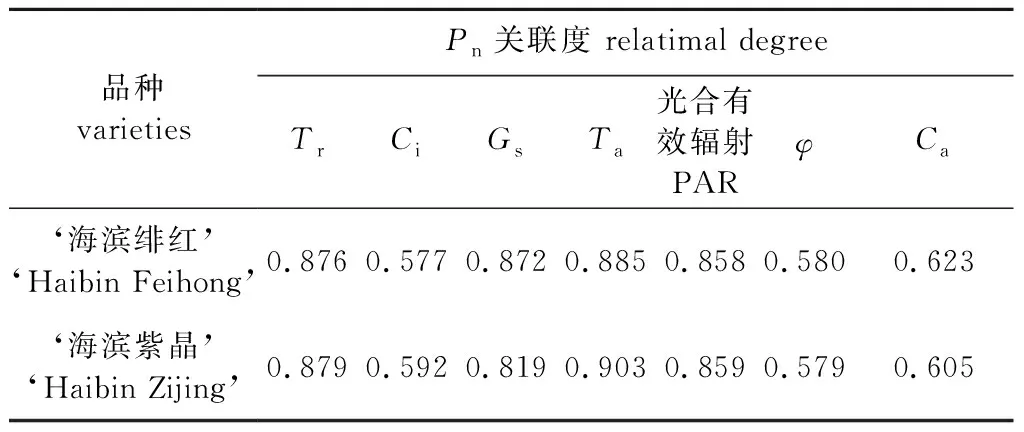
表2 净光合速率与生理、生态因子的灰色关联度
2.5 不同品种光响应曲线及特征参数比较
光响应曲线反映了植物光合速率随光照强度改变而变化的规律。通过叶子飘新模型的拟合,得到的Pn-PAR曲线(图3A),决定系数R2>0.99,表明该方法拟合程度较高。由图3A可知,2个品种对光合有效辐射的响应曲线基本一致,净光合速率随着光合有效辐射的升高而增加,当光照强度达到饱和点后,净光合速率增长曲线变化趋于平缓。
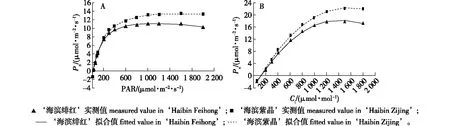
图3 两个乌桕新品种Pn-PAR和Pn-CO2响应曲线的拟合Fig.3 Simulation of Pn-PAR, and Pn-CO2 response curves of T. sebifera in two new varieties
通过计算得到两个品种的最大净光合速率(Pn,max)、表观量子效率(AQY)、暗呼吸速率(Rd)、光补偿点(LCP)、光饱和点(LSP)等相关光合作用参数(表3)。结果表明,‘海滨紫晶’的最大净光合速率(Pn,max)、光饱和点(LSP)、暗呼吸速率(Rd)和光补偿点(LCP)均高于‘海滨绯红’,表观量子效率(AQY)则低于‘海滨绯红’,其中Pn,max、LSP和Rd两个品种间存在显著差异(P< 0.05)。
2.6 不同品种CO2响应曲线及特征参数比较
CO2响应曲线反映了植物光合速率随胞间CO2浓度改变而变化的规律。通过叶子飘新模型的拟合,得到的Pn-CO2曲线(图3B),R2>0.98,表明该方法拟合程度较高。由图3B可知,两个品种对CO2浓度的响应曲线基本一致,净光合速率随着CO2浓度的升高而增加,当CO2浓度达到饱和点,净光合速率稳定在一定水平。

表3 两个乌桕新品种光响应曲线相关参数
通过计算得到两个品种的最大光合能力(An,max)、羧化效率(CE)、光呼吸速率(Rp)、CO2补偿点(CCP)、CO2饱和点(CSP)等相关参数(表4)。结果表明,‘海滨紫晶’最大光合能力(An,max)、CO2饱和点(CSP)、羧化效率(CE)和光呼吸速率(Rp)均高于‘海滨绯红’,CO2补偿点(CCP)则低于‘海滨绯红’,其中An,max和CSP存在显著性差异(P<0.05)。

表4 两个乌桕品种CO2响应曲线相关参数
3 讨 论
植物叶片光合色素含量和植物光合能力密切相关[20-21],可直接反映植物叶片光合能力的大小[22]。在光合作用中,叶绿素a和叶绿素b具有捕获和吸收光能的作用[23],类胡萝卜素作为一种重要的抗氧化剂,能够吸收过多的光能防止脂质过氧化并稳定细胞膜结构[24]。本研究中,‘海滨紫晶’的叶绿素a、叶绿素b、类胡萝卜素含量和叶绿素a/b均大于‘海滨绯红’,其中叶绿素a、叶绿素b和类胡萝卜素差异显著,表明‘海滨紫晶’对光能的吸收能力以及强光下的光保护能力较‘海滨绯红’强,同时其叶绿素a/b值的也部分反映了‘海滨紫晶’对强光环境具有更强的适应能力。
净光合速率可直接反应植物光合作用的强弱[25],晴朗天气下植物的光合作用日变化模式一般分为两种曲线类型,单峰型和双峰型,大多数植物在夏季表现为双峰型变化[26]。本研究中,两个乌桕新品种的Pn日变化曲线均为典型的单峰型,无“午休”现象,说明该两个新品种均具有较强的对夏季高温强光的适应能力。但‘海滨紫晶’Pn日均值和峰值均明显高于‘海滨绯红’,说明‘海滨紫晶’具有更强的净光合能力。从日变化趋势结合Pn与生态因子的灰色关联分析结果,即‘海滨紫晶’与Tr关联度更大,而与Ca的关联度相对较小,表明‘海滨紫晶’随着一天中环境气温的增高更有利于其净光合速率的增强,而对大气CO2浓度的下降其Pn受影响程度相对较小,也反映出‘海滨紫晶’具有更强的夏季高温环境的适应能力。从不同气体交换参数的日变化趋势看,尽管两个品种的变化趋势相同,即Pn、Gs和Tr呈单峰型,而Ci呈相反的“V”型,但‘海滨绯红’的Pn、Gs值整体明显低于‘海滨紫晶’的;同时Ci却要高于‘海滨紫晶’。根据光合作用气孔与非气孔限制的理论推断[27],‘海滨绯红’净光合能力低于‘海滨紫晶’主要是受非气孔因素的限制,应与其光能利用效率或CO2利用能力相对较低有关。
光响应曲线及其特征参数能较好地判定植物对光的利用和适应能力[28-29],在同一栽培环境下植物的光合能力不同,光响应特征差异显著[30-31]。在本研究中,‘海滨紫晶’Pn,max显著高于‘海滨绯红’,这与Pn日变化的测定结果一致。AQY可反映植物在光合作用过程中对光能的利用效率,特别是弱光条件下对光能的利用效率[32],Rd也与植物的耐弱光性相关。‘海滨绯红’AQY较大,Rd显著低于‘海滨紫晶’,表明其叶片吸收和转换光能的色素蛋白复合体可能较多,利用弱光的能力较强,同时能通过降低暗呼吸作用来间接提高光合性能。此外,植物对光照条件的需求可通过LSP和LCP反映[33]。一般情况下,植物叶片LSP和LCP均较高的植物,能够很好地在强光下进行光合作用[21],本研究中‘海滨紫晶’对强光有着更强的适应性。
CO2响应曲线及其特征参数能反映植物对CO2的利用能力[34]。An,max是植物在CO2饱和点处达到的最大净光合速率[35],CE与RuBPCase酶的活性呈正相关[36]。测定结果显示,‘海滨紫晶’An,max与CE均较高,有研究指出,An,max和CE较高的植物其叶片1,5-二磷酸核酮糖(RuBP)的再生能力及其对CO2的同化能力与利用效率也高[37]。光呼吸作为一种消耗过剩光能、保护光合机构免于强光破坏的途径[38],‘海滨紫晶’光呼吸速率高以及饱和CO2时的净光合速率也较高,表明其能在强光条件下保持较高的光呼吸以免于光合机构破坏。CCP低的植物通常具有较高的净光合速率和产量,CSP表示植物对低浓度CO2的利用能力。‘海滨紫晶’CCP较低,而CSP较高,表明其CO2同化能力强且利用幅度较宽。此外,两个品种在CO2饱和浓度下的最大净光合速率较光饱和时的高,这表明了乌桕在光照条件饱和的情况下,光合速率在更大程度上受CO2供应的限制。因此,在夏季强光热条件基础上[39]研究发现,‘海滨紫晶’较强的羧化效率和RUBP再生能力所表现出的强CO2利用能力可能是其比‘海滨绯红’具有更强光合性能的主要原因。
综上,两个乌桕新品种在光合特性上均表现出较强的夏季生长适应性。‘海滨紫晶’光合性能最优,其CSP和LSP均较高,可通过较优的强光适应能力和较宽的CO2利用幅度来提高光合性能;‘海滨绯红’次之,其主要通过降低暗呼吸作用减少消耗和提高弱光利用能力来积累更多的有机物质用于生长。‘海滨紫晶’较强的光合能力与其所受的非气孔限制相对较小有关,较强的CO2利用能力可能是其改善非气孔限制的主要原因,光呼吸可能是其在高温强光下避免光抑制和光破坏的一种机制。在栽培生产中,可通过调节光强或CO2浓度等手段来提高这两个新品种的光合速率,促进其生长发育。研究结果为乌桕新品种推广应用提供理论依据,由于本研究只从光合特性的角度探讨了两个乌桕新品种的夏季生长适应性,生长实际应用尚需结合生长指标等的测定并进行长期的观测。
猜你喜欢
花卉(2022年17期)2022-09-06
林业与生态(2022年5期)2022-05-23
连云港文学(2022年2期)2022-05-10
散文诗(2017年17期)2018-01-31
现代园艺(2017年21期)2018-01-03
新课程·下旬(2017年7期)2017-08-14
浙江农业科学(2017年5期)2017-06-21
西北林学院学报(2015年5期)2015-01-03
植物营养与肥料学报(2011年2期)2011-10-26
