
不同播种量对小麦泰科麦33干物质积累转运、旗叶光合特性及产量构成的影响
2021-03-25 13:50吕广德殷复伟王超李宁钱兆国吴科
江苏农业学报 2021年1期
吕广德 殷复伟 王超 李宁 钱兆国 吴科

摘要: 于2016-2019年小麦生长季进行小麦品种泰科麦33的田间播种量试验,2016-2017年播种量为75 kg/hm2(SA1)、150 kg/hm2(SA2)、225 kg/hm2(SA3)、300 kg/hm2(SA4)、375 kg/hm2(SA5)5个水平,2017-2018和2018-2019兩个小麦生长季调整播种量为75.0 kg/hm2(SA6)、112.5 kg/hm2(SA7)、150.0 kg/hm2(SA8)、187.5 kg/hm2(SA9)、225.0 kg/hm2(SA10)5个水平,同时在2018-2019年重复2016-2017年的试验。测定不同播种量条件下泰科麦33生育阶段的干物质累积量,并根据干物质累积量分析生物量积累特征以及开花前后干物质转运特性;通过测定开花后旗叶的蒸腾速率、气孔导度和净光合速率,分析不同播种量条件下泰科麦33的光合作用差异;对不同播种量条件下泰科麦33籽粒产量和产量结构的差异进行分析。结果表明,在所有播种量条件下,开花前干物质累积量高于开花后干物质累积量,但对籽粒贡献率却低于开花后;在SA1~SA5处理中,与SA1、SA3、SA4、SA5相比,SA2的干物质累积量显著提高,2016-2017年分别提高了5.9%、1.6%、8.6%、9.7%,2018-2019年分别提高了6.5%、1.5%、11.5%、13.6%;在SA6~SA10处理中,2017-2018年干物质累积量SA9处理最高,为14 779.17 kg/hm2,2018-2019年SA8处理最高为17 405.25 kg/hm2。不同播种量下泰科麦33干物质积累动态曲线均符合 Logistic 模型,在SA1~SA5处理中,两年干物质最大积累速率(Vm)均出现在SA2处理,与各处理平均值相比,干物质最大累积量分别提高了4.1%和6.7%, 快速累积持续时间虽然分别缩短了7.6 d和6.1 d,但最大累积速率分别提升了14.9%和15.6%;SA6~SA10处理中,2017-2018年SA9处理的干物质最大积累速率最高,与各处理平均值相比,干物质最大累积量提高了7.4%, 快速累积持续时间延长了0.8 d,最大累积速率提升了0.6%;2018-2019年,干物质最大累积量出现在SA8处理,与各处理平均值相比,干物质最大累积量提高了5.2%, 快速累积持续时间虽然缩短了1.8 d,但最大累积速率提升了7.9%。在所有播种量条件下,泰科麦33旗叶净光合速率和气孔导度均在开花后7 d达到最高,蒸腾速率在开花后14 d最高;SA1~SA5处理中,两年SA1处理的蒸腾速率比其他处理的平均值分别高9.1%和9.2%,气孔导度分别高15.0%和14.0%,净光合速率分别高15.1%和12.3%;SA6~SA10处理中,SA6处理在花后的蒸腾速率高于其他处理的平均值9.2%,气孔导度高17.3%,净光合速率高11.8%。但在SA1~SA3处理间旗叶光合参数差异不显著,SA6~SA10处理中SA6~SA8处理间差异也不显著。在SA1~SA5处理中,各处理籽粒产量的大小顺序为SA2>SA3>SA1>SA4>SA5,SA2两年产量分别为9 545.05 kg/hm2和9 439.50 kg/hm2。在SA6~SA10处理中,2017-2018年各播种量处理产量的大小顺序为SA9>SA8>SA10>SA7>SA6,SA9的产量为8 342.55 kg/hm2,2018-2019年各处理的产量顺序为SA8>SA9>SA7>SA10>SA6,SA8的产量为9 287.95 kg/hm2。通过拟合泰科麦33播种量与产量之间的方程曲线发现,在 SA1~SA5处理下产量达到理论最大值时最佳播种量两年分别为179.16 kg/hm2和 159.70 kg/hm2;在SA6~SA10处理下2017-2018年播种量为188.96 kg/hm2时产量最大,而2018-2019年播种量为153.70 kg/hm2时产量最大。以上分析结果说明,优质小麦新品种泰科麦33在播种量153.70~188.96 kg/hm2条件下产量、干物质累积量和光合速率最高。
关键词: 小麦;泰科麦33;播种量;干物质;光合;产量
中图分类号: S512.1+10.42 文献标识码: A 文章编号: 1000-4440(2021)01-0016-13
Effects of sowing amounts on dry matter accumulation and distribution, photosynthetic characteristics of flag leaves and yield composition of wheat TKM 33
LYU Guang-de1, YIN Fu-wei2, WANG Chao1, LI Ning3, QIAN Zhao-guo1, WU Ke1
(1.Taian Academy of Agricultural Sciences,Taian 271000,China;2.Taian Agricultural Technology Promotion Station,Taian 271000, China;3.Taian Heyuan Seed Technology Co., Ltd, Taian 271000,China)
Abstract: Sowing amount test of wheat variety TKM33 was carried out in wheat growing season during 2016-2019. The sowing amount in 2016-2017 was set in five levels containing 75 kg/hm2 (SA1), 150 kg/hm2 (SA2), 225 kg/hm2 (SA3), 300 kg/hm2 (SA4) and 375 kg/hm2 (SA5). In two wheat growing seasons of 2017-2018 and 2018-2019, the sowing amounts were adjusted to five levels containing 75.0 kg/hm2 (SA6), 112.5 kg/hm2 (SA7), 150.0 kg/hm2 (SA8), 187.5 kg/hm2 (SA9) and 225.0 kg/hm2 (SA10). The sowing amount test in 2018-2019 was conducted based on repeating the test in 2016-2017. Dry matter accumulation of TKM33 was measured at growth stage under conditions of different sowing amounts. Biomass accumulation characteristics and transport characteristics of dry matter before and after flowering were analyzed based on the dry matter accumulation. Differences of photosynthesis of TKM33 under different sowing amounts conditions were analyzed by determining the transpiration rate, stomatal conductivity and net photosynthetic rate of flag leaves after flowering. The differences of grain yield and yield structure of TKM33 under different sowing amounts conditions were analyzed. The results showed that under conditions of all sowing amounts, the dry matter accumulation before flowering was higher than that after flowering, but the contribution rate to grain before flowering was lower than that after flowering. In SA1-SA5 treatments, the dry matter accumulation of SA2 increased significantly compared with that of SA1, SA3, SA4 and SA5, which increased 5.9%, 1.6%, 8.6% and 9.7% respectively in 2016-2017 and increased 6.5%, 1.5%, 11.5% and 13.6% respectively in 2018-2019. In SA6-SA10 treatments, the dry matter accumulation in 2017-2018 was the highest under SA9 treatment, which was 14 779.17 kg/hm2, but the highest value in 2018-2019 was under SA8 trearment, which was 17 405.25 kg/hm2. The dynamic curve of dry matter accumulation under different sowing amounts all conformed to the Logistic model. In SA1-SA5 treatments, the largest dry matter accumulation rate (Vm) of two years all appeared in SA2 treatment, the largest dry matter accumulation increased by 4.1% and 6.7% compared with the average value of all treatments, although rapid accumulation duration shortened by 7.6 days and 6.1 days respectively, but the largest accumulation rate increased by 14.9% and 15.6% respectively. The max accumulation rate of dry matter was the highest in SA9 treatment among SA6-SA10 treatments in 2017-2018. Compared with the average value of all treatments, the max accumulation of dry matter in SA9 treatment increased by 7.4%, the duration of rapid accumulation increased by 0.8 day, and the max accumulation rate increased by 0.6%. In 2018-2019, the max accumulation of dry matter appeared in SA8 treatment. Compared with the average value of each treatment, the max accumulation of dry matter increased by 5.2%, the rapid accumulation duration decreased by 1.8 days, although the max accumulation rate increased by 7.9%. Under all sowing amounts conditions, the net photosynthetic rate and stomatal conductance of flag leaves of TKM33 all reached the highest values seven days after flowering, and the transpiration rate reached the highest value 14 days after flowering. In SA1-SA5 treatments, the transpiration rate of SA1 treatment during the two years was 9.1% and 9.2% higher than the average value of other treatments, the stomatal conductance was 15.0% and 14.0% higher, and the net photosynthetic rate was 15.1% and 12.3% higher respectively. In SA6-SA10 treatments, the transpiration rate of SA6 treatment after flowering was 9.2% higher than the average value of other treatments, the stomatal conductance increased by 17.3% and the net photosynthetic rate increased by 11.8%. However, there was no significant difference in photosynthetic parameters of flag leaves between SA1-SA3 treatments and between SA6-SA8 treatments within SA6-SA10 treatments. In SA1-SA5 treatments, the grain yield showed the trend of SA2>SA3>SA1>SA4>SA5, and the yield of SA2 treatment in two years was 9 545.05 kg/hm2 and 9 439.50 kg/hm2 respectively. In SA6-SA10 treatments, the yields of all sowing amouts treatments showed the trend of SA9>SA8>SA10>SA7>SA6 in 2017-2018, and the yield of SA9 was 8 342.55 kg/hm2. In 2018-2019, the yields of all treatments showed the trend of SA8>SA9>SA7>SA10>SA6, and the yield of SA8 was 9 287.95 kg/hm2. By fitting the equation curve between sowing amount and yield of TKM33, it was found that the theoretical optimal sowing amount in two years was 179.16 kg/hm2 and 159.70 kg/hm2 respectively when the theoretical max yield was reached under SA1-SA5 treatments. Under the treatments of SA6-SA10, the max yield appeared when the sowing amount was 188.96 kg/hm2 in 2017-2018, and in 2018-2019 the max yield was got when the sowing amount was 153.70 kg/hm2. The above analysis result indicated that, the high-quality new wheat variety TKM33 showed the best yield, dry matter accumulation and photosynthetic rate under the sowing amount of 153.70-188.96 kg/hm2.
Key words: wheat;TKM33;sowing amounts;dry matter;photosynthetic;yield
小麦是中国重要的粮食作物,其种植面积在中国粮食作物种植面积中占比高达20%~29%[1]。自上世纪90年代以来,中国小麦产量水平快速提高,其中小麦品种、栽培措施等因素起了重要作用。播种量是小麦生产中较易控制的栽培措施,也是影响小麦群体结构、产量形成的重要因子[2-3]。小麦合理群体结构有利于解决群体与个体之间的矛盾,改良叶层结构,提高光能利用效率,增加群体生物量积累,实现穗大、粒多、粒饱,达到高产、高效的目标[4-7]。播种量较小时,单株小麦所得到的营养物质较为丰富,小麦地上部的分蘖数量增多,但小麦单株分蘖能力和成穗能力由于播种量较小并不能满足高产群体的需要;播种量较大时,小麦单株分蘖能力下降,无效分蘖增多,成穗率降低,造成土壤养分和光能利用的浪费。所以合理的播种量在小麦群体数量、群体质量和资源利用等方面有重要意义。泰科麦33是2018年通过黄淮麦区和山东省审定的优质高产小麦品种,具有分蘖力中等、壮蘖多、分蘖成穗率高等特点。本研究通过连续3年播种量试验,研究泰科麦33在不同播种量下的干物质积累转运特性、旗叶光合特性以及籽粒产量的差异,为泰科麦33在黄淮麦区推广种植提供参考。
1 材料与方法
1.1 试验地概况与试验设计
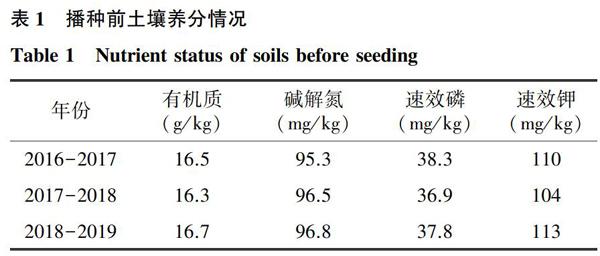
1.1.1 试验基本情况 试验地位于山东省泰安市马庄镇泰安市岳洋农作物专业合作社科技示范基地(36°00′N,117°01′E)。海拔86.8 m,2016-2017年小麦季降水量194.2 mm,平均气温10.1 ℃;2017-2018年小麦季降水量246.9 mm,平均气温9.7 ℃;2018-2019年小麦季降水量109.9 mm,平均气温9.3 ℃,具体数据如图1所示。试验地土壤为黏土,播种前试验田0~20 cm土壤养分见表1。
1.1.2 试验设计 小麦品种选用优质小麦新品种泰科麦33。2016-2017小麦生长季播种量为75 kg/hm2(SA1)、150 kg/hm2(SA2)、225 kg/hm2(SA3)、300 kg/hm2(SA4)、375 kg/hm2(SA5)5个水平,3次重复,播种日期为2016年10月1日;在获得泰科麦33合理播种量范围前提下,于2017-2018和2018-2019两个小麦生长季调整播种量为75.0 kg/hm2(SA6)、112.5 kg/hm2(SA7)、150.0 kg/hm2(SA8)、187.5 kg/hm2(SA9)、225.0 kg/hm2(SA10)5个水平,每个处理均为3次重复,播种日期分别是2017年10月11日和2018年10月11日。同时在2018-2019年设置与2016-2017年相同播种量的试验。以上所有试验的每个处理面积均为80.0 m×2.1 m,采用随机区组设计。肥料用量2/3基施,所用肥料为氮磷钾配比为14∶16∶15的复合肥750 kg/hm2,1/3拔节期追施,施用30∶0∶5(N∶P2O5∶K2O)的复合肥375 kg/hm2。其他栽培管理措施同一般大田。
1.2 测定项目与方法
1.2.1 干物质积累及分配 在泰科麦33三叶期调查基本苗,于小麦冬前期(2016-11-22、2017-12-07、2018-12-07)、起身期(2017-02-14、2018-02-21、2019-02-21)、拔节期(2017-03-23、2018-03-25、2019-03-25)、开花期(2017-04-27、2018-04-28、2019-04-28)以及成熟期(2017-06-04、2018-06-07、2019-06-05)取样,其中小麦冬前期、起身期和拔节期取10棵单株,开花期和成熟期连续取30个单茎,所有植株取地上部105 ℃殺青1 h,80 ℃烘至恒质量,称干质量。干物质积累转运计算公式如下[8]:冬前期、起身期和拔节期干物质积累量(kg/hm2)=不同时期单株干物质量(g)×种植密度(1 hm2株数)/1 000;开花期和成熟期干物质积累量(kg/hm2)=不同时期单茎干物质量(g)×穗数(1 hm2穗数)/1 000;开花前贮藏同化物转运量(kg/hm2)=开花前营养器官干物质量(kg/hm2)-[成熟期干物质重(kg/hm2)-籽粒干物质量(kg/hm2)];开花前同化物转运率=开花前贮藏同化物转运量(kg/hm2)/开花前干物质积累量(kg/hm2)×100%;开花前干物质对籽粒的贡献率=开花前贮藏同化物转运量(kg/hm2)/成熟期籽粒干质量(kg/hm2)×100%。
1.2.2 利用Logistic 曲线拟合泰科麦33干物质积累量 各处理生物量动态增长用Logistic模型Y=K/(1+aebt)拟合[9],其中Y 为积累量(kg/hm2),K为理论最大积累量(kg/hm2),t 为生长天数(d)。分别对模型公式求一阶、二阶、三阶导数, 得到相应生长曲线的快速积累期起始时期(t1)、快速积累期终止时期(t2)、最大积累速率(Vm)、最大积累速率出现时间(tm),t1=1bln2+ 3a,t2=1bln2- 3a,T=t2-t1,Vm=-bK4,tm=-lnab 。
1.2.3 旗叶光合参数测定 在开花期随机选取长势均匀的 10 株小麦挂牌标记,小麦开花后每隔7 d在9∶00-11∶00使用 CIRAS-2便携式光合作用系统测定旗叶的光合参数(净光合速率Pn、蒸腾速率Tr和气孔导度Gs)[10]。
1.2.4 群体基本苗、籽粒产量及产量构成因素调查 在出苗后三叶期选取1 m2调查基本苗。在成熟期每个小区选取1 m2调查穗数,随机取10穗调查穗粒数。脱粒后自然风干至含水量为13.0% 时测定千粒质量。实时收获籽粒产量。每个处理3次重复。
1.3 数据处理
采用 Excel 2017 和 SPSS 20 软件对数据进行统计分析和线性方程作图。采用R语言进行相关性分析并作图。采用单因素(one-way ANOVA)和LSD法进行方差分析和多重比较(α=0.05)。
2 结果与分析
2.1 不同播种量条件下泰科麦33干物质的积累转运特征
2.1.1 不同播种量条件下泰科麦33干物质积累特征 由表2可知,各个播种量处理下,R2均达到极显著水平,干物质累积符合Logistic模型。在SA1~SA5处理中,2016-2017年和2018-2019年干物质积累特征基本一致,随着播种量的增加,干物质最大累积量先升高后降低,两年结果均在SA2处理播种量下最高,分别达到19 716.36 kg/hm2和19 526.28 kg/hm2。随着播种量的增加,干物质快速积累期终止时间延后,干物质快速积累持续期延长,而SA2处理下干物质快速积累期开始时间(两年分别为156.9 d和149.5 d)和干物质最大累积速率出现时间(分别为第191.3 d和第182.5 d)最晚,但干物质最大累积速率最高,分别为188.3 kg/(hm2·d)和194.5 kg/(hm2·d),明显高于其他处理。与各处理平均值相比,干物质最大累积量两年分别提高了4.1%和6.7%, 快速累积持续时间虽然分别缩短了7.6 d和6.1 d,但最大累积速率分别提升了14.9%和15.6%。
在SA6~SA10处理中,2017-2018年和2018-2019年的干物质积累规律不一致。随着播种量的增加,2017-2018年的干物质最大积累量在SA9处理最高,为16 469.51 kg/hm2,在该播种量条件下,干物质快速积累开始时间和终止时间均延后,快速积累时间持续64.4 d,干物质最大累积速率最高,为168.5 kg/(hm2·d),出现在第179.7 d,与各处理平均值相比,干物质最大累积量提高了7.4%, 快速累积持续时间延长了0.8 d,最大累积速率提升了0.6%。而2018-2019年,干物质最大积累量却出现在SA8处理,为20 366.79 kg/hm2,在该播种量下,干物质快速积累开始时间延后,快速积累时间持续71.2 d,干物质最大累积速率出现在第182.9 d,为188.2 kg/(hm2·d),与各处理平均值相比,干物质最大累积量提高了5.2%, 快速累积持续时间虽然缩短了1.8 d,但最大累积速率提升了7.9%。
由图2可以看出,随着生育进程,不同播种量下泰科麦33生育期干物质累积量呈“慢–快–慢”的 S 型曲线变化。在SA1~SA5处理中,在拔节期之前,随着播种量的增加,干物质累积量增加;在拔节期之后,SA2处理的干物质累积量迅速增加;到收获期,干物质积累量表现为SA2>SA3>SA1>SA4>SA5,与SA5相比,SA2、SA5、SA1、SA4的两年平均干物质累积量分别增加了11.6%、9.9%、5.1%、1.4%。
在SA6~SA10处理中,在拔节期之前,随着播种量的增加,干物质积累量也呈现增加的趋势;在拔节期之后,2017-2018年,SA9处理的干物质积累量增加明显,而2018-2019年,SA8的干物质积累量增加明显,两年结果不一致。在2017-2018年,收获期干物质累积量表现为SA9>SA8>SA10>SA7>SA6,其中SA9处理较其他处理干物质积累量分别增加了305.54 kg/hm2(SA8)、329.62 kg/hm2(SA10)、1 132 .91 kg/hm2(SA7)和1 976.99 kg/hm2(SA6),增幅分别为2.1%、2.3%、8.3%、15.4%。而2018-2019年表现为SA8>SA9>SA10>SA7>SA6,SA8處理较其他处理分别增加了175.84 kg/hm2(SA9)、719.77 kg/hm2(SA10)、808.00 kg/hm2(SA7)和1 812.82 kg/hm2(SA6),增幅分别为1.0%、4.3%、4.9%、11.6%。
2.1.2 不同播种量条件下泰科麦33干物质分配转运 由表3可知,在SA1~SA5处理中,2016-2017年和2018-2019年的泰科麦33干物质分配转运规律基本一致。随着播种量的增加,泰科麦33的生物量和收获指数呈先增加后降低的趋势,两年均在SA2播种量条件下最高,分别为16 961.75 kg/hm2、48.1%和17 048.13 kg/hm2、47.3%。泰科麦33出苗到开花阶段的两年干物质累积量分别占整个生育期的71.5%~74.5%和71.8%~75.4%,说明出苗到开花阶段为干物质积累的主要时期。开花前干物质转运量对籽粒的贡献率分别为40.6%~44.0%和40.2%~45.3%,开花后干物质对籽粒的贡献率分别为56.0%~59.4%和54.7%~59.8%,说明开花后干物质生产对籽粒起重要作用。通过方差分析发现,除了开花到成熟阶段干物质累积量存在年度差异外,其余指标年度差异不显著,而播种量对各阶段干物质积累和分配转运有显著影响。试验年份和播种量的互作对泰科麦33干物质积累转运影响显著。
在SA6~SA10处理中,两年结果不一致,其中2017-2018年生物量和收获指数SA9处理最高,分别为14 779.17 kg/hm2和48.1%,生物量高于其他处理平均数的6.8%;2018-2019年SA8处理最高,分别为17 405.25 kg/hm2和45.5%,生物量比其他处理平均数高5.3%。干物质积累分配规律与SA1~SA5处理的积累分配规律基本一致。方差分析结果表明,SA6~SA10处理的年度差异和播种量显著,播种量和试验年份的互作对泰科麦33干物质积累转运有显著影响。
2.2 不同种植密度条件下泰科麦33的光合特性
由于天气原因,2017-2018年泰科麦33的光合特性未能测定,我们只分析SA1~SA5处理2016-2017年、2018-2019年光合数据以及SA6~SA10处理2018-2019年的光合数据。由图3可以看出,泰科麦33旗叶蒸腾速率在开花后 14 d 达到最高,之后呈下降趋势。SA1~SA5处理中,两年SA1处理的蒸腾速率比其他处理平均值分别高9.1%和9.2%,且最高值出现在SA1处理的第14 d,分别为8.99 mmol/(m2·s)和9.42 mmol/(m2·s);SA6~SA10处理中,SA6播种量的蒸腾速率比其他处理平均值高9.2%,最高值出现在SA6处理的第14 d,为9.56 mmol/(m2·s)。泰科麦33旗叶气孔导度在开花后第7 d 达到最高,在第14 d时缓慢下降,之后呈快速下降的趋势。SA1~SA5处理中,两年SA1处理的气孔导度比其他处理平均值分别高15.0%和14.0%,且最高值出现在SA1处理的第7 d,分别为0.91 mmol/(m2·s)和0.95 mmol/(m2·s);SA6~SA10处理中,SA6处理的气孔导度比其他处理平均值高17.3%,最高值出现在SA6处理的第7 d,为0.96 mmol/(m2·s)。泰科麦33旗叶净光合速率和气孔导度变化趋势一致,均在开花后第7 d 达到最高,之后呈下降趋势。SA1~SA5处理中,两年SA1处理的净光合速率比其他处理平均值分别高15.1%和12.3%,最高值同样出现在SA1处理的第7 d,分别为25.59 μmmol/(m2·s)和25.97 μmmol/(m2·s);SA6~SA10处理中SA6净光合速率高于其他处理平均值11.8%,SA6处理的第7 d净光合速率值为 25.43 μmmol/(m2·s)。
同一测定时期,随着播种量的增加,蒸腾速率、气孔导度和净光合速率均出现下降的趋势。但通过方差分析发现,SA1~SA5处理中,泰科麦33的花后旗叶蒸腾速率、气孔导度和净光合速率在SA1~SA3处理之间差异基本不显著;SA6~SA10处理中,泰科麦33的花后旗叶蒸腾速率、气孔导度和净光合速率在SA6~SA8处理之间差异也基本不显著。
2.3 不同播种量对小麦泰科麦33籽粒产量及产量结构的影响
由表4可以看出,SA1~SA5处理中,随着播种量的增加,2016-2017年和2018-2019年泰科麦33籽粒产量和产量结构的变化规律基本一致,穗数增加,穗粒数减少,千粒质量降低,籽粒产量先增加后降低。其中穗数变幅两年分别为1 hm25.607×106~8.133×106穗和4.805×106~7.035×106穗,SA5处理较SA1处理分别增加45.1%和46.4%,SA4处理与SA5处理差异不显著。SA5处理与SA1处理相比,穗粒数分别减少15.9粒和8.8粒,千粒质量分别减少5.8 g和5.9 g。籽粒产量的顺序为SA2>SA3>SA1>SA4>SA5, SA2处理两年分别为9 545.05 kg/hm2和9 439.50 kg/hm2,较SA5处理增幅分别为20.4%和21.6%。由方差分析可知,试验年份和播种量对泰科麦33籽粒产量及产量结构的影响显著,试验年份和播种量的互作除对籽粒产量影响不显著外,对产量结构的影响显著。
在SA6~SA10处理中,随着播种量的增加,2017-2018年和2018-2019年泰科麦33籽粒产量及其结构的变化规律基本一致,穗数增加,穗粒数减少,千粒质量降低,籽粒产量先增加后降低。其中穗数变幅两年分别为1 hm2 4.358×106~5.993×106穗和4.358×106~6.840×106穗,SA10处理较SA6处理分别增加37.5%和60.0%。SA10处理与SA6处理相比,穗粒数分别减少10.0粒和7.8粒,千粒质量分别减少2.2 g和5.5 g。两年籽粒产量的变化趋势存在差异,2017-2018年,各播种量处理的产量高低顺序为SA9>SA8>SA10>SA7>SA6,SA9处理产量为8 342.55 kg/hm2,较SA6处理增幅32.8%;2018-2019年,各播种量处理的产量高低顺序为SA8>SA9>SA7>SA10>SA6,SA8处理产量为9 287.95 kg/hm2,较SA6处理增幅18.7%。两年结果表现不一致,原因是201
2.4 不同播种量条件下小麦泰科麦33籽粒产量及产量结构的相关性
通过不同播种量处理下泰科麦33籽粒产量及产量结构的相关性分析(图4)可知,穗数与穗粒数呈负相关关系,其中穗数与穗粒数除在2018-2019年SA6~SA10处理中呈显著负相关外,其余处理均为极显著负相关;穗数与千粒质量呈负相关关系,在2018-2019年各处理中呈显著负相关,在2017-2018年处理中呈极显著负相关;穗粒数与千粒质量呈正相关关系,在2018-2019年处理中呈极显著相关,在2016-2017年和2017-2018年处理中呈显著相关。从产量与产量三要素的相关性分析来看,在SA1~SA5处理中,产量与穗数呈负相关,与千粒质量和穗粒数呈正相关,相关性均达到显著水平。在SA6~SA10处理中,2017-2018年产量与穗粒数和千粒质量呈显著负相关,与穗数呈正相关;2018-2019年,产量与穗数、穗粒数、千粒质量呈正相关。SA6~SA10处理中,两年度的产量与产量三要素之间的关系存在差异,原因可能与2017-2018年小麦拔节期遇到倒春寒天气有关,对泰科麦33产量三要素的影响较大。
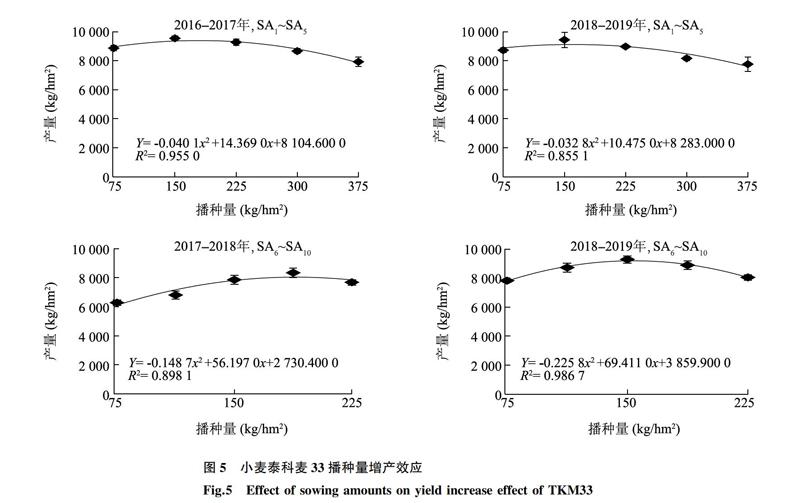
2.5 小麦泰科麦33播种量与产量的关系曲线分析
根据表4中结果,对不同播种量处理的泰科麦33产量进行比较分析,拟合出播種量与产量之间的关系曲线(图5)。其中2016-2017年SA1~SA5处理,播种量和产量的关系曲线为 Y=-0.040 1x2+14.369 0x+8 104.600 0, R2=0.955 0,由方程可得播种量从 75.00 kg/hm2增加到 179.16 kg/hm2时,泰科麦33产量逐渐增加并达到最大值9 391.80 kg/hm2;2018-2019年SA1~SA5处理,关系曲线为 Y=-0.032 8x2+10.475 0x+8 283.000 0,R2=0.855 1,由方程可得播种量为159.70 kg/hm2时,产量达到最大值9 119.32 kg/hm2;2017-2018年SA6~SA10处理,关系曲线为 Y=-0.148 7x2+56.197 0x+2 730.400 0,R2=0.898 1,由方程可得播种量为188.96 kg/hm2时,产量达到最大值8 039.92 kg/hm2;2018-2019年SA6~SA10处理关系曲线为 Y=-0.225 8x2+69.411 0x+3 859.900 0,R2=0.986 7,由方程可得播种量为153.70 kg/hm2时,产量达到最大值9 194.14 kg/hm2。
3 讨论
3.1 合理的播种量可以提高泰科麦33干物质累积量和改善干物质的分配转运
干物质积累转运是小麦产量形成的物质基础。有研究者发现,较高的干物质积累能够显著提高小麦的产量水平[11-12]。播种量是影响小麦干物质积累转运的重要因素[13-14]。本试验发现,随着播种量的增加,泰科麦33的生物量呈先增加后降低的趋势,这与胡卫丽等[15]的研究结果一致。在播种量SA1~SA5处理中,在150 kg/hm2播种量条件下干物质累积量最高。在SA6~SA10处理中,2017-2018年在187.5 kg/hm2播种量下干物质累积量最高,而2018-2019年在150 kg/hm2播种量下干物质累积量最高,两年结果差异可能与2018年泰科麦33拔节期遇到强烈倒春寒天气有关,使相同播种量条件下的穗数与往年相比减少。有研究结果表明,小麦籽粒灌浆物质1/3来自于开花前营养器官中贮藏的光合同化物向籽粒的转移,2/3来自于花后同化物的积累[16-20],因此,增加花后同化物的积累是获得高产的基础,适当增加播种量可以提高花后干物质累积量及其籽粒的贡献率[21]。本试验发现,所有播种量试验结果均显示花后同化物对籽粒的贡献率高于花前干物质积累对籽粒的贡献率。其中在SA1~SA5处理中,150 kg/hm2播种量的花后同化物累积量显著高于其他播种量水平;在SA6~SA10处理中,由于气候因素导致两年结果不一致,分别在187.5 kg/hm2和150.0 kg/hm2播种量下花后干物质累积量最高。试验中泰科麦33花后干物质对籽粒灌浆的贡献率均低于前人研究的66.7%的水平[16-20],但显著提高了开花前积累的干物质向籽粒转运。
3.2 合理的播种量可以提高泰科麦33单株花后旗叶的光合能力
合理的播种量可以构建合理的小麦群体结构,进而优化协调单株株型,提高小麦的产量和经济系数[22]。合理的单株株型对小麦光合能力的提高有显著作用,小麦光合作用产生的碳水化合物为干物质积累的基础,小麦营养器官碳同化的强弱和光合产物在营养器官积累的多寡,都直接影响后期小麦的籽粒质量和产量形成[23-24]。有研究结果表明,小麦产量的90%~95%与光合作用有关[25]。张向前等[26]研究发现,1 hm2 1.80×106株种植密度的光合参数(蒸腾速率、气孔导度和净光合速率)高于1 hm2 2.40×106株种植密度,这与本研究结果基本一致。我们发现,在SA1和SA6播种量水平(1 hm2 1.35×106株)下光合参数高于SA2和SA8播种量水平(1 hm2 2.64×106株)下的光合参数。通过方差分析发现,在同一环境下,以上各个播种量水平下的光合参数之间差异不显著,说明SA1~SA5处理中,75.0 kg/hm2和150.0 kg/hm2播种量之间的光合能力均可以满足籽粒灌浆期间碳同化积累的需求。但当播种量增加到187.5 kg/hm2以上时,各个播种量条件下的光合参数与75.0~150.0 kg/hm2播种量条件下的光合参数之间的差异显著,说明在187.5 kg/hm2以上播种量的单株光合能力显著降低,187.5 kg/hm2以上的播种量均显著降低了籽粒产量和干物质量。但该结果只是在产量和干物质量确定的情况下得到的,并且只说明了单株的光合能力,并未在群体结构上说明光合作用与产量的关系。所以今后有必要针对不同播种量水平下的泰科麦33群体光合作用进行研究,补充说明适宜播种量(合理群体)下的光合能力与产量的关系。
3.3 合理的播种量可以提高泰科麦33的籽粒产量
增加播种量可以提高小麦的穗数,但当播种量增加到一定的数量后,穗数并不会显著提高,而且产量三要素中穗粒数减少、千粒质量降低,最终导致产量下降。陈爱大等[27]发现,镇麦168穗数在1 hm2 1.80×106 ~3.30×106株播种量下,随着播种密度的增加而增加。刘莹等[28]发现,襄麦D31在1 hm21.80×106~3.15×106株播种密度下,随着播种密度的增加,有效穗数显著提高,穗粒数显著下降,千粒质量变化不显著,这与本试验研究结果不一致。泰科麦33播种量在75~300 kg/hm2 (1 hm21.33×106~4.07×106株)下,有效穗数显著提高,穗粒数和千粒质量显著下降。还有研究结果表明,一定范围内增加种植密度可以提高小麦产量,但如果继续增加密度则会显著减产[29-30],且通常获得最高产量的密度多在1 hm22.25×106~3.75×106株[31-33],相应穗数多集中在1 hm25.15×106~6.60×106穗[34-35],该结果与我们的研究结果基本一致。我们发现,泰科麦33在1 hm22.64×106株密度下,相应的穗数为1 hm26.13×106穗可以获得最高产量。但我们的研究结果却与郑飞娜等[36]对泰农18在常规条播下种植密度的研究结果有差异,其研究发现,泰农18在1 hm23.40×106株密度时才能获得最高产量。原因可能与所用小麦品种的特性不同有关,泰科麦33是一个多穗型品种,分蘖成穗强于大穗型品种泰农18,所以泰科麦33在低于泰农18的种植密度下即可获得高产。
在小麦产量的3个构成因素中,穗数是对产量补偿能力及自动调节能力最強的因素。田纪春等[37]研究结果表明,穗数对小麦产量的正效应最大。周延辉等[38]通过对稻茬小麦产量及其构成因素分析发现,产量构成因素与产量的相关程度为穗数>千粒质量>穗粒数,且都是正相关关系。周继泽等[39]通过对河南省五大主导品种适宜播种量研究发现,矮抗58、百农207和郑麦7698的穗数与产量呈负相关,西农979和周麦22的穗数与产量呈正相关关系,说明不同品种穗数和产量的相关性存在差异。增加穗数最直接的栽培措施是增加播种量。我们研究发现,泰科麦33在播种量为SA1~SA5条件下,产量与穗粒数、千粒质量呈正相关,却与穗数呈负相关,与周延辉等的研究结果不一致[38];在2018-2019年播种量为SA6~SA10条件下,产量与穗数、穗粒数及千粒质量均呈显著正相关,且相关程度为穗数>千粒质量>穗粒数,与周延辉等的研究结果一致[38]。分析原因,可能与我们所设置的播种量有密切关系,我们通过播种量与产量的相关性分析发现,在SA1~SA5条件下播种量与产量呈负相关,在SA6~SA10条件下播种量与产量呈正相关。
4 结论
增加播种量,小麦穗数增加,但穗粒数、千粒质量、蒸腾速率、气孔导度和净光合速率均呈下降的趋势。但合理的播种量,可以构建合理的群体结构,促使蒸腾速率、气孔导度和净光合速率下降不显著,从而增加干物质累积量和籽粒产量。在本试验条件下,对优质高产小麦品种泰科麦33来说,播种量153.70~188.96 kg/hm2是获得籽粒高产的播种量。
参考文献:
[1] 石江荣, 任永康, 王 芳. 氮素营养对超高产小麦调控的研究进展[J]. 山西农业科学,2010, 38(3): 80-82.
[2] TOKATLIDIS I S. Addressing the yield by density interaction is a prerequisite to bridge the yield gap of rain-fed wheat[J]. Annals of Applied Biology, 2014, 165: 27-42.
[3] DAI X L, ZHOU X H, JIA D Y, et al. Managing the seeding rate to improve nitrogen-use efficiency of winter wheat[J]. Field Crops Research, 2013, 154: 100-109.
[4] 姜 东,于振文,李永庚,等. 高产小麦营养器官临时贮存物质积运及其对粒重的贡献[J]. 作物学报, 2003, 29(1): 31-36.
[5] 刘丽平,胡焕焕,李瑞奇,等. 行距配置和密度对冬小麦品种河农 822 群体质量及产量的影响[J].华北农学报, 2008, 23(2): 125-131.
[6] 李 宁,段留生,李建民,等. 播期与密度组合对不同穗型小麦品种花后旗叶光合特性、籽粒库容能力及产量的影响[J].麦类作物学报, 2010, 30(2): 296-302.
[7] 蒋会利. 播期密度对不同小麦品种群体茎数及产量的影响[J]. 西北农业学报, 2012, 21(6): 67-73.
[8] 姜 东,谢祝捷,曹卫星,等. 花后干旱和渍水对冬小麦光合特性和物质运转的影响[J]. 作物学报, 2004, 30(2): 175-182.
[9] 赵 姣,郑志芳,方艳茹,等. 基于动态模拟模型分析冬小麦干物质积累特征对产量的影响[J]. 作物学报, 2013, 39(2): 300-308.
[10]郭培武,赵俊晔,石 玉,等. 水肥一体化对小麦水分利用和光合特性的影响[J]. 应用生态學报, 2019, 30(4): 1170-1178.
[11]张维城,王志和,任永信,等. 有效分蘖终止期控制措施对小麦群体质量影响的研究[J]. 作物学报, 1998, 24(6): 903-907.
[12]田中伟,王方瑞,戴廷波,等. 小麦品种改良过程中物质积累转运特性与产量的关系[J]. 中国农业科学, 2012, 45(4):801-808.
[13]HILTBRUNNER J, STREIT B, LIEDGENS M. Are seeding densities an opportunity to increase grain yield of winter wheat in a living mulch of white clover[J]. Field Crops Research, 2007, 102: 163-171.
[14]张 敏,王岩岩,蔡瑞国,等. 播期推迟对冬小麦产量形成和籽粒品质的调控效应[J]. 麦类作物学报, 2013, 33(2):325-330.
[15]胡卫丽,王永华,李刘霞,等. 氮密调控对两种穗型冬小麦品种茎蘖干物质积累与转运的影响[J]. 麦类作物学报, 2014, 34(6): 808-815.
[16]李世清,邵明安,李紫燕,等. 小麦籽粒灌浆特征及影响因素的研究进展[J]. 西北植物学报, 2003, 23(11): 2031-2039.
[17]牟会荣,姜 东,戴廷波,等. 遮荫对小麦旗叶光合及叶绿素荧光特性的影响[J]. 中国农业科学, 2008, 41(2): 599-606.
[18]李东升,温明星,蔡金华,等. 播期和密氮组合对镇麦10号干物质积累及产量的调控效应[J]. 麦类作物学报, 2015, 35(10): 1426-1432.
[19]吴 祯,张保军,海江波,等. 不同种植方式对冬小麦花后干物质积累与分配特征及产量的影响[J]. 麦类作物学报, 2017, 37(10): 1377-1382.
[20]马尚宇,王艳艳,刘雅男,等. 播期、播种量和施氮量对小麦干物质积累、转运和分配及产量的影响[J]. 中国生态农业学报(中英文), 2020, 28(3): 375-385.
[21]张小涛,黄玉芳,马晓晶,等. 播种量和施氮量对不同基因型冬小麦干物质累积、转运及产量的影响[J]. 植物生理学报, 2017, 53(6): 1067-1076.
[22]刘万代,陈现勇,尹 钧,等. 播期和密度对冬小麦豫麦49-198群体性状和产量的影响[J].麦类作物学报, 2009, 29(3): 464-469.
[23]柴守玺,赵德明,常 磊. 西北绿洲种植密度对冬小麦产量及生理指标的影响[J]. 生态学报, 2008, 28(1): 292-301.
[24]房 锋,张朝贤,黄红娟,等. 麦田节节麦发生动态及其对小麦产量的影响[J]. 生态学报, 2014, 34(14):3917-3923.
[25]卓武燕,张正茂,刘苗苗,等. 不同类型小麦光合特性及农艺性状的差异[J]. 西北农业学报, 2016, 25(4): 538-546.
[26]张向前,陈 欢,赵 竹,等. 密度和行距对早播小麦生长、光合及产量的影响[J]. 麦类作物学报, 2015, 35(1): 86-92.
[27]陈爱大,蔡金华,温明星,等. 追施氮肥对强筋小麦‘镇麦168产量和品质的影响[J]. 西南农业学报, 2014, 27(3): 1154-1158.
[28]刘 莹,唐 清,王立峰,等. 播期和密度对襄麦 D31籽粒产量及品质的影响[J]. 麦类作物学报, 2017, 37(3): 376-381.
[29]CARR P M, HORSLEY R D, POLAND W W. Tillage and seeding rate effects on wheat cultivars: I. Grain production[J]. Crop Science, 2003, 43: 202-209.
[30]LLOVERAS J, MANENT J, VIUDAS J, et al. Seeding rate influence on yield and yield components of irrigated winter wheat in a Mediterranean climate[J]. Agron J, 2004, 96: 1258-1265.
[31]陈雨海,余松烈,于振文. 小麦生长后期群体光截获量及其分布与产量的关系[J]. 作物学报, 2003, 29(5):730-734.
[32]曹 倩,贺明荣,代兴龙,等. 密度、氮肥互作对小麦产量及氮素利用效率的影响[J]. 植物营养学与肥料学报, 2011,17(4): 815-822.
[33]YANG D Q, CAI T, LUO Y L, et al. Optimizing plant density and nitrogen application to manipulate tiller growth and increase grain yield and nitrogen-use efficiency in winter wheat[J]. Peer J, 2019, 7: e6468.
[34]MA S C, WANG T C, GUAN X K, et al. Effect of sowing time and seeding rate on yield components and water use efficiency of winter wheat by regulating the growth redundancy and physiological traits of root and shoot[J]. Field Crops Research, 2018, 221:166-174.
[35]張永丽,肖 凯,李雁鸣. 种植密度对杂种小麦 C6-38/Py85-1 旗叶光合特性和产量的调控效应及其生理机制[J]. 作物学报, 2005, 31(4): 498-505.
[36]郑飞娜,初金鹏,张 秀,等. 播种方式与种植密度互作对大穗型小麦品种产量和氮素利用率的调控效应[J]. 作物学报, 2020, 46(3): 423-431.
[37]田纪春,邓志英,胡瑞波,等. 不同类型超级小麦产量构成因素及籽粒产量的通径分析[J]. 作物学报, 2006, 32(11): 1699-1705.
[38]周延辉,朱新开,郭文善,等. 稻茬小麦中高产水平下产量及其构成因素分析[J]. 麦类作物学报, 2018, 38(3): 293-297.
[39]周继泽,欧行奇,王永霞,等. 河南省五大主导小麦品种适宜播种量研究[J]. 农学学报, 2019, 9(2): 1-6.
(责任编辑:张震林)
收稿日期:2020-07-05
基金项目:泰安市科技计划引导计划项目(2019NS094);山东省重点研发计划项目(2016GNC113004);国家小麦现代农业产业技术体系项目(CARS-3-2-21)
作者简介:吕广德(1987-),山东滨州人,硕士,农艺师,主要从事小麦遗传育种与高产栽培技术研究。(E-mail)2007guangd@163.com
通讯作者:吴 科,(E-mail)sdtawuke1964@126.com;钱兆国,(E-mail)qianzhaoguo@163.com
猜你喜欢
河南农业·综合版(2021年11期)2021-11-30
河南农业·综合版(2021年4期)2021-06-08
河南农业·综合版(2021年2期)2021-03-24
河南农业·综合版(2021年2期)2021-03-24
农民致富之友(2019年32期)2019-11-23
清明(2019年6期)2019-11-21
中国经贸导刊(2018年16期)2018-08-02
新农村(2017年17期)2017-08-23
世界热带农业信息(2014年12期)2015-01-21
世界热带农业信息(2014年8期)2014-09-23
