
祁连山南麓高寒灌丛GPP 变化特征及对生长季积温的响应
2021-03-26 06:53祝景彬贺慧丹李红琴张法伟李英年杨永胜张光茹王春雨罗方林
草业科学 2021年2期
祝景彬 ,贺慧丹,李红琴,张法伟, ,李英年, ,杨永胜, ,张光茹, ,王春雨, ,罗方林,
(1. 枣庄学院旅游与资源环境学院,山东 枣庄 277160;2. 中国科学院西北高原生物研究所,青海 西宁 810001;3. 洛阳师范学院生命科学学院,河南 洛阳 471934;4. 中国科学院高原生物适应与进化重点实验室,青海 西宁 810001;5. 中国科学院大学,北京 100049)
由于人类对化石燃料的过度使用,致使空气中温室气体含量逐渐增加,导致全球气温升高[1],“温室效应”将会对陆地生态系统的碳循环过程产生深刻的影响[2-4]。青藏高原是世界上最年轻的高原,并且是我国草地分布最广的区域[5-6]。分布在青藏高原的大面积天然高寒草地,对于整个青藏高原生态系统的碳平衡具有至关重要的作用,对全球陆地生态系统碳循环也具有重要的影响[7]。由于青藏高原低温、高海拔的特殊环境,使青藏高原草地生态系统极其脆弱,对气候变化和环境因素改变的响应极其敏感[8]。青藏高原高寒灌丛生态系统分布面积为0.106 × 106km2,是青藏高原高寒草地的重要组成部分[9]。因此,研究高寒灌丛生态系统的碳平衡动态,对于准确认识青藏高原高寒草地碳循环过程具有重要意义[10-11]。
生态系统总初级生产力(gross primary productivity,GPP)是指植被通过光合作用制造的全部有机质总量,是生态系统碳循环的基础[9]。大气中CO2浓度会对GPP 产生重要影响,而GPP 的改变也会对大气中CO2浓度造成反馈影响,二者相互作用影响着气候变暖的过程[7-8]。GPP 受植物本身的光合作用潜力、叶面积指数、群落结构,以及光合有效辐射、温度、有效土壤水分、养分等诸多因子的共同影响[12]。植被生长发育需要一定的温度(热量)条件,植物进行光合作用过程中相关酶的生理活性以及酶促反应速度受到温度调控[13],温度是影响植物光合作用的关键因子[14-16]。但是,温度与总初级生产力的关系较为复杂,一定程度上温度的升高有利于光合速率的提高,但是超过其最适反应温度又开始下降[17]。此外,有研究表明,群落生物量对生态系统热量条件变化的反应具有一定的“时滞”性,在植被生长发育所需要的其他条件均得到满足时,温度越高越有利于植被的生长发育,并且当温度累积到一定程度时,植被才能完成其整个生长发育过程,在生长季的这个温度的累积数称为生长季积温(growing season degree days, GDD)。因此,分析生态系统气温的累加值与植被光合生产能力的关系在一定程度上更能客观地反映其真实的响应模式[18]。
国内外对陆地生态系统碳循环方面的研究主要集中在低海拔地区,而对于高海拔地区青藏高原高寒生态系统碳循环方面的研究相对较少[19-20];此外,目前对青藏高原高寒生态系统碳平衡的研究局限于短期内的变化动态和影响机制的研究[21],而对于较长时间尺度的年际变化动态和响应机制研究相对较少[4,11]。在高寒地区的研究表明,气候变暖提前了融雪期和植被返青期,延长了生长季长度,并促进高寒生态系统的CO2同化和植被生产[22]。近几十年来,青藏高原经历了显著的变暖,但有关高寒灌丛生态系统碳平衡的研究相对较少[23]。同时,全球气候变化的背景下,永冻层和冰川融化可能导致青藏高原不同类型草地生态系统分配格局的改变[24]。此外,研究表明在不同的时间尺度上,气候环境因子可能对碳循环有不同的影响[11]。在不同时间尺度上分析气候环境因子对碳通量的影响,更能准确地反映其客观关系。因此,本研究在不同时间尺度上分析了2003−2016 年生长季利用涡度相关系统在青藏高原东北部高寒灌丛获得的14 年连续通量数据,旨在:1)量化高寒灌丛生态系统生长季GPP 与热量条件[GDD、Ta(气温)]在不同时间尺度上的变化特征;2)明确不同时间尺度上高寒灌丛生态系统生长季的GPP 对热量条件(GDD、Ta)的响应。
1 材料与方法
1.1 试验地概况
采集数据的涡度相关系统位于青海省海北藏族自治州门源县境内的高寒灌丛生态系统,研究地点的具体位置属于高寒草甸生态系统国家野外科学观测研究站(海北站)。海北野外观测站地处青藏高原的东北边缘,地理位置为37°29′−37°45′ N,101°12′−101°23′ E,海拔为3 200~3 600 m。该地无明显的四季之分,一年之中只分为冷热两季,在11 月至次年4 月明显干燥寒冷,而在5 月至10 月则明显温暖湿润。全年平均气温约为−1.0 ℃,最热月为7 月,而最冷月为1 月。全年平均降水量为580 mm,但是降水在一年之中分布极不均衡,大约有80%的降水集中在植被生长季[25]。青藏高原高寒灌丛生态系统的植被主要由两层组成,分别为灌木层和禾本科层。青藏高原高寒灌丛生态系统的建群种为金露梅(Potentilla fruticosa),其他群落组成物种主要包括矮嵩草(Kobreia humilis)、垂穗披碱草(Elymus nutans)、异针茅(Stipa aliena)和麻花艽(Gentiana straminea)等。青藏高原高寒灌丛生态系统的土壤为一种粘性壤土[26-27]。
1.2 研究材料及分析方法
1.2.1 通量和相关监测
利用涡度相关系统对研究区CO2通量进行长期连续的观测(2003−2016 年)。通量观测的主要仪器为开路式涡度相关观测系统和常规气象观测系统,其中涡度相关系统包括三维超声风速仪(CSAT3, Campbell Scientific, Logan, USA)、红 外 气体分析仪(LI-7500, LI-COR Lincoln, USA)和数据采集器(CR5000, Campbell Scientific, Logan, USA)。常 规气象观测系统观测项目主要包括温度、湿度、辐射、风向、风速等。光合有效辐射( photosynthetic photon flux density, PPFD)传感器(L1190SB, LI-COR, Lincoln,USA)安装在高1.2 m 的支架上。空气温度/湿度传感器(Model HMP45C, Vaisala, Helsinki, Finland)安装在 防 辐 射 罩 内(Model 41002, RM Young Company,Michigan, USA)。土壤温度(soil temperature, Ts)主要是利用热电偶温度传感器(107-L, Campbell Scientific,Logan, USA)分别测定5、10、20、40 和80 cm 处的Ts。5、20 和50 cm 处的土壤含水量(soil water content, SWC)利用时域反射计(Model CS615-L, Campbell Scientific,Logan, USA)测定。
生长季有效积温(GDD)为大于5 ℃日平均气温的和[28]。参考以往研究经验,将生长季定义为高寒灌丛生态系统的5 月 − 9 月[11,14]。本研究中日尺度GPP、Ta为当天的GPP、Ta;月尺度GPP、GDD 为当月每天GPP、Ta的和;生长季尺度GPP 、GDD 为整个生长季每天GPP、Ta的和。
1.2.2 数据处理和插补
由于野外各种因素的不确定性,导致仪器运行会出现异常,导致通量塔记录数据缺失[29]。本文利用研究时段内的辐射有效能(净辐射与土壤热通量之差)与湍流热通量之比的能量闭合率来简单评价高寒灌丛通量数据的数据质量。高寒灌丛通量数据在生长季的能量闭合率为0.78。有关能量不闭合的研究和相关的结论较多,因此本研究没有深入讨论能量不闭合的原因[4,11]。但从数据分析来看,高寒灌丛通量观测的数据有效性较高,完全可以满足后续的数据分析。
本研究严格采用ChinaFLUX 推荐的通量数据处理方法来进行数据分析[30-31]。对于缺失的通量数据,一般通过非线性经验方程进行插补。本研究夜间通量数据(Reco,n)采用其与5 cm 深处土壤温度(Ts)的Van′t Hoff 函数[公式(1)]进行插补,生长季白天的碳通量数据(NEE)则利用与PPFD 的Michaelis-Menten 函数[公式(2)]进行插补[32-33]。

式中:Reco,ref为生态系统在10 ℃时的参考呼吸速率,Q10为生态系统呼吸熵(呼吸随温度升高10 ℃所增加的倍数),反映了生态系统呼吸对温度的敏感性。

式中:Reco,d为生态系统白天的呼吸速率,a 和Pmax分别为生态系统表观光量子效率和潜在最大CO2同化速率(即PPFD 趋向极大时生态系统净光合速率)。以上参数均为方程拟合值。
涡度相关系统无法直接测定生态系统总初级生产力(GPP)和生态系统呼吸(Re),需利用公式外推得到。涡度相关系统观测的CO2通量为NEE,将夜间呼吸函数外延至白天(Reco,d),即可得生态系统的总呼吸量(Re),进而可得生态系统总初级生产力[GPP,公式(3)],日GPP 计算如公式(3)所示,日Re为白天呼吸(Reco,d)和夜间呼吸(Reco,n)之和[27]。本研究中GPP 和GDD 的关系采用线性回归分析,利用Excel 和SPSS 20.0 软件对数据进行统计分析。

2 结果与分析
2.1 高寒灌丛生态系统的日GPP 变化特征及其对气温的响应
高寒灌丛生态系统的日GPP 在2003−2016 年生长季(5 月 − 9 月)的变化趋势基本一致,表现为先增加后减小的单峰变化趋势(图1)。14 年的数据表明,日GPP 在每年7 月达到峰值,日GPP 的最大值为2009 年7 月17 日的8.02 g·(m2·d)−1。高寒灌丛在2003−2016 年生长季(5 月 − 9 月)日平均气温(Ta)变化趋势基本一致,表现为先升高后降低的单峰变化趋势(图1)。最高气温出现在2010 年7 月29 日,为17.26 ℃;最低气温出现在2009 年5 月1 日,为−1.93 ℃。线性回归分析表明(图2),日GPP 与Ta呈极显著正相关关系(GPP = 0.41Ta+ 0.42,R2= 0.50,P < 0.001),由此表明在日尺度上气温越高,高寒灌丛植被的光合生产能力越强。

图1 高寒灌丛日总初级生产力(GPP)与气温(Ta)的变化特征Figure 1 Variation characteristics of daily gross primary productivity (GPP) and temperature (Ta) of alpine shrubland
2.2 高寒灌丛生态系统的月GPP 变化特征及其对GDD 的响应
高 寒 灌丛 在2003−2016 年生 长 季(5 月 − 9 月)GPP 月变化趋势基本一致,表现为先增大后减小的单峰变化趋势,2003−2016 年生长季的月GPP 基本都在7 月达到峰值,只有2003 年在8 月达到峰值,为130.54 g·m−2;月GPP 的最大值为2015 年7 月的值,为191.49 g·m−2,7 月GPP 的均值为165.49 g·m−2(图3)。2003−2016 年每年5 月的GPP 为当年生长季的月GPP 最小值,5 月GPP 的均值为23.02 g·m−2。6、8、9 月GPP 的均值分别为93.47、146.68 和75.66 g·m−2。在2003−2016 年生长季(5 月 − 9 月)的月GDD 变化趋势基本一致(图3),在7、8 月达到峰值,7、8 月GDD的均值分别为302.80 和302.75 ℃。5 月GDD 最小,其均值为70.70 ℃;6、9 月GDD 均值分别为206.43和107.54 ℃。

图2 高寒灌丛日总初级生产力(GPP)与气温(Ta)的关系Figure 2 The relationship between daily gross primary productivity (GPP) and temperature (Ta) of alpine shrubland
线性回归分析表明(图4),在整个生长季的月GPP 与 月GDD 极 显 著 正 相 关(GPP = 0.49GDD +5.93,R2= 0.84,P < 0.001),此外,分别分析了2003−2016 年生长季每个月的GPP 与GDD 的关系(表1),研究发现在5、9 月GPP 与GDD 没有显著相关性(P >0.05),在6、7、8 月GPP 与GDD 显著相关(P < 0.05),并且在生长旺盛期(7、8 月)的相关性更显著(P <0.01),在7 月GPP 与GDD 的相关性最强(R2= 0.54,P = 0.003)。这种不同月份GPP 与GDD 的相互关系存在差异(表1)可能是由不同月份之间气候环境因子的差异造成的。因此,在生长季旺盛期(7、8 月)这种相关性最强,说明在高寒灌丛植被生长最旺盛时期,热量条件对植被光合生产能力有更显著的影响。整体上看,在月尺度上,生长季每个月积温越大,越有利于生态系统总初级生产力的提高。

图3 高寒灌丛的月总初级生产力(GPP)与生长季有效积温(GDD)的变化特征Figure 3 Variation characteristics of monthly gross primary productivity (GPP) and growing season degree days (GDD) over alpine shrubland

图4 高寒灌丛的月总初级生产力(GPP)与生长季有效积温(GDD)的关系Figure 4 The relationship between monthly gross primary productivity (GPP) and growing season degree days (GDD) over alpine shrubland
2.3 高寒灌丛生态系统的生长季GPP 变化特征及其对GDD 的响应
如图5 所示,高寒灌丛在2003−2016 年整个生长季(5 月 − 9 月) GPP (R2= 0.64,P = 0.001)的值和GDD(R2= 0.44,P = 0.01)的值具有一定的变异性,但整体上表现为逐渐增加的趋势。生长季GPP 的最大值出现在2014 年,为562.13 g·m−2;生长季GPP 的最小值出现在2003 年,为406.32 g·m−2;2003−2016 年的生长季GPP 均值为507.11 g·m−2。2004−2016 年整个生长季总GDD 的最大值为2016 年的1 119.15 ℃,最小值为2004 年的826.35 ℃,整个生长季GDD 的均值为975.93 ℃。线性回归分析表明(图6),生长季GPP 与GDD 显著正相关(GPP = 0.35GDD + 164.99,R2= 0.39,P = 0.016),由此说明在生长季尺度上,高寒灌丛生态系统生长季有效积温(GDD)的增大有利于提高整个生长季的总初级生产力(GPP)。
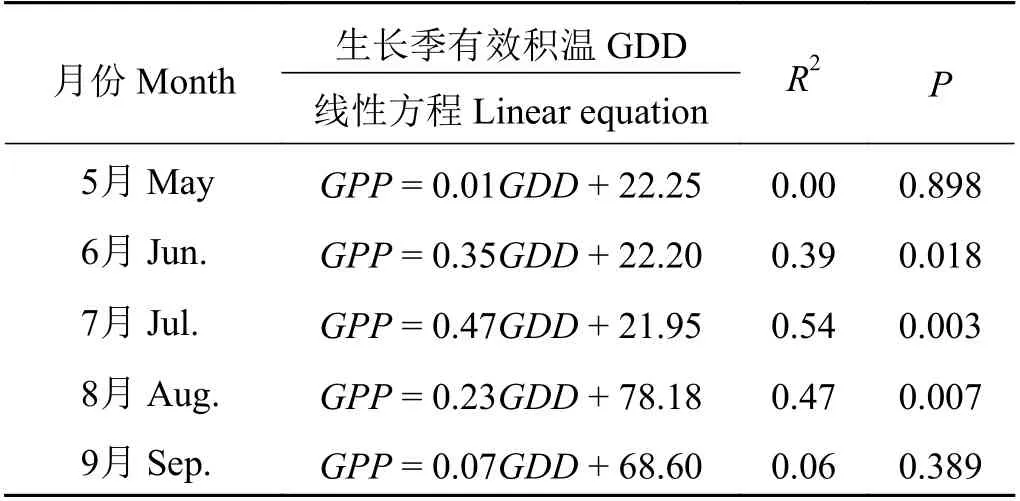
表1 高寒灌丛在生长季每个月总初级生产力(GPP)与生长季有效积温(GDD)的相关性Table 1 Correlation between gross primary productivity(GPP) and growing season degree days (GDD) of each month in growing season over alpine shrubland

图5 高寒灌丛的生长季(5 月 − 9 月)总初级生产力(GPP)与生长季有效积温(GDD)变化特征Figure 5 Variation characteristics of gross primary productivity (GPP) and growing season degree days(GDD) in growing season (May-Sep.) over alpine shrubland
3 讨论

图6 高寒灌丛的生长季(5 月 − 9 月)总初级生产力(GPP)与生长季有效积温(GDD)关系Figure 6 The relationship between seasonal gross primary productivity (GPP) and growing season degree days (GDD) in growing season (May-Sep.) over alpine shrubland
本研究中,GPP 和热量条件(Ta、GDD)在日尺度、月尺度的变化趋势都表现为先升高后降低的单峰变化趋势,都是在7、8 月达到峰值,这和前人有关 青 藏 高 原 高 寒 灌 丛 的 研 究 结 果 基 本 一 致[4,6,10,14]。在生长季尺度上,2003−2016 年高寒灌丛的生长季GPP 和GDD 具有一定波动性,但整体上表现为逐渐增加的趋势,这与前人对于青藏高原全球气候变化背景下气温升高的研究结论相一致[7,13]。研究表明,随着温度的升高,生态系统GPP 有增大趋势,但增大缓慢且具有较高异质性[13]。降水、温度、CO2浓度和地表覆盖变化等都会改变植物蒸腾、光合等生理活动,从而引起陆地生态系统GPP 的变化,进而影响其碳平衡。大量研究表明,热量条件对于高寒生态系统植被的光合生产能力具有至关重要的作用,但也发现群落生物量对生态系统热量条件变化的反应具有一定的“时滞”性。因此,分析生态系统气温的累加值与植被光合生产能力的关系在一定程度上更能客观地反映其真实的响应模式[34]。所以,在全球气候变化的背景下,研究总初级生产力对热量条件的响应极其重要。
大量研究表明,不同生态系统的碳平衡状态存在较大差异,这是由不同生态系统生物和非生物因素之间的差异造成的[18-19]。同时,年均温的年际差异也导致了碳循环各过程的波动[20]。但是,过去的研究大多局限于短期的研究,研究表明在不同的时间尺度上环境因子可能对生态系统生产力有不同的影响[21,26],因此本研究在不同时间尺度上探讨了GPP 对热量条件(Ta、GDD)的响应。研究结果发现,在不同时间尺度上GPP 与热量条件(Ta、GDD)都呈显著正相关关系,这与柴曦等[4]、Zhao 等[13]和Li等[11]对青藏高原高寒灌丛的研究结果一致,并且Kato 等[21]、Saito 等[32]对本研究地点附近的高寒矮嵩草草甸的研究也有类似的研究结果。由此说明,高寒灌丛生态系统植被的光合生产能力直接受到热量条件的控制,这可能是因为温度的累积效应是打破植被休眠、植被返青和生长代谢的主要控制因素[9,21,35]。因此,高寒植被在生长代谢方面受到温度的重要影响[36]。此外,温度能够影响土壤中微生物的生物量,以及相关酶的活性,由于高寒生态系统土壤中含有大量未分解的土壤有机质,并且微生物分解土壤有机质的过程对温度极其敏感,较高的温度能够刺激土壤有机质的分解,为植被的生长提供更多的养分,因此间接有利于提高植被的光合生产能力[21,23]。此外,青藏高原东部高寒灌丛草毡土的水分含量达到30%以上,远高于高原腹地的草原土,而且生长季开始就处于比较稳定的高值,受降水量的影响没有草原土那么大[4];另一方面,灌丛植被根系要深于草甸植被根系,降低了植被根系对表层土壤干旱的敏感性,缓解表层土壤水分的缺失对根系的影响[11,13]。因此,热量条件是高寒生态系统最为普遍的环境影响因子[21,32],它通过影响生态系统的生态过程,例如冠层发育、蒸散速率和土壤水分等来影响植被光合生产能力及其对气候变化的响应[6, 11, 21, 32]。
但值得注意的是,本研究对2003−2016 年生长季中每个月份的GPP 与GDD 作了线性回归分析,发现在5、9 月GPP 与GDD 没有显著相关性,在植被生长旺盛期(7、8 月)相关性较强,在7 月GPP 与GDD 的相关性最显著。这可能是由在生长季中不同月份的气候环境因子之间的差异造成的[11-14]。研究结果显示,在5、9 月GPP 与GDD 没有显著相关性,这可能是因为在生长季初期和末期植被生长缓慢[6,14],光合生产能力较弱,而此时有效积温也较小,所以有效积温的改变对植被光合生产能力的影响并不显著。但是,7 月份雨热条件最好[4,19],植被生长最旺盛,植被在这个时期的光合生产能力最强,因此在这个时期有效积温的变化,会对生态系统的总初级生产力产生极显著的影响,说明此时热量条件对GPP 变化的控制作用更强。总体上看,高寒灌丛生态系统的总初级生产力受到热量条件的显著影响,即有效积温越大,越有利于植被光合生产能力的提高。本研究的结果表明,在全球变暖的背景下,有利于青藏高原高寒灌丛总初级生产能力的提高,但是有研究也表明温度的升高也会增加生态系统呼吸[11],因此对于高寒灌丛生态系统的碳平衡状态还需要进一步深入的研究。
本研究结果表明热量条件是高寒灌丛植物生长的重要限制因子,温度升高会促进湿润地区植物光合作用和碳固定。但是,温度升高会导致土壤水分蒸发的增加,加剧植被受干旱胁迫的程度,可能导致植被生产力降低[13,21],因此应加强青藏高原不同植被类型生态系统植被光合生产能力对气候变化响应的研究。Wang 等[37]利用CENTURY 模型对松嫩平原的预测表明,一定幅度温度的增加,有利于提高植被光合生产能力,当温度升高超过一定阈值,将会造成植被光合生产能力的下降。所以,高寒灌丛GPP 对温度升高是否存在阈值效应,以及具体阈值如何等问题仍需进一步深入分析。并且,高寒灌丛生态系统的GPP 不仅受到气候变化的影响,还会受到人类活动的影响,因此应该加强对气候变化和人类活动对高寒灌丛GPP 变化相对贡献率的研究。此外,本研究仅探讨有效积温对高寒灌丛生态系统总初级生产力的研究,仍有一定的局限性。在今后的研究中应加强对实测生物量、碳储量、碳通量等资料的收集和整理,为生态系统模拟模型的验证和评价工作提供充分的依据。
4 结论
1)高寒灌丛在生长季的日GPP、Ta与月GPP、GDD 都表现为单峰变化趋势;而生长季总GPP 与GDD 具有较高变异性,但整体上表现为逐渐增加的趋势(P < 0.05)。
2)在日尺度、月尺度、生长季尺度上,GPP 与热量条件(Ta、GDD)都呈显著正相关关系(P < 0.05)。在7 月份的月GPP 与GDD 相关性最显著(P < 0.01)。整体上看,高寒灌丛GDD 的增加有利于促进GPP的提高,暗示在全球气候变暖的背景下将会提高青藏高原高寒灌丛生态系统植被的光合生产能力。
猜你喜欢
化工管理(2022年14期)2022-12-02
水土保持学报(2022年5期)2022-10-10
中国农业科学(2022年15期)2022-08-09
中国马铃薯(2022年2期)2022-07-05
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01
现代畜牧科技(2021年9期)2021-10-13
四川农业科技(2019年10期)2020-01-06
黑龙江粮食(2019年10期)2019-11-23
草业科学(2019年5期)2019-06-04
天津师范大学学报(自然科学版)(2016年4期)2016-12-14
