
不同生态恢复阶段种内竞争影响下的马尾松种群死亡率研究
2021-03-28 02:57王晓宁崔煜婕张滋隆
福建农业科技 2021年1期
王晓宁 崔煜婕 张滋隆

摘 要:种内竞争作为长期存在且不断调整的生态关系,是影响种群的死亡率变化不可忽略的因素。为明确种内竞争在生态恢复中对马尾松种群死亡率的影响,在福建省长汀县设立了3处样地,对先锋种马尾松的存活状况进行调查,并使用Hegyi竞争指数量化了种内竞争强度。结果表明:先锋阶段后期,马尾松个体大小分布分散,种内竞争最为激烈,先锋阶段中期与早期,马尾松个体大小变异性低,且早期的种内竞争强度高于中期。伴随种内竞争强度的增加,幼苗的死亡风险提升。种内竞争在生态恢复先锋阶段的早期与中期,显著影响马尾松种群死亡率,而先锋阶段后期,种内竞争不是影响马尾松种群死亡率的主要因素。研究结果表明,种内竞争对马尾松种群的影响具有阶段化的差异,种群内不同大小的个体对于种内竞争的响应不同。
关键词:生态恢复;种内竞争;马尾松;种群死亡率
中图分类号:S 791.248 文献标志码:A 文章编号:0253-2301(2021)01-0026-06
DOI:10.13651/j.cnki.fjnykj.2021.01.005
Abstracts:Intraspecific competition, as a long-standing and constantly adjusted ecological relationship, is an important factor affecting the variation of population mortality. In order to clarify the effect of intraspecific competition on the population mortality of Pinus massoniana during ecological restoration, three sample plots were set up in Changting County of Fujian Province. The survival status of pioneer species of Pinus massoniana was investigated, and the intensity of intraspecific competition was quantified by using Hegyi competition index. The results showed that in the later period of pioneer stage, the individual size distribution of Pinus massoniana was scattered, and the intensity of intraspecific competition was the most intense. In the middle and early periods of pioneer stage, the variability of individual size of Pinus massoniana was low, and the intensity of intraspecific competition in the early stage was higher than that in the middle stage. With the increase of intraspecific competition intensity, the mortality risk of seedling increased. Intraspecific competition significantly affected the population mortality of Pinus massoniana in the early and middle periods of pioneer stage during ecological restoration, but in the later period of pioneer stage, intraspecific competition was not the main factor affecting the population mortality of Pinus massoniana. The research results showed that the effects of intraspecific competition on the population of Pinus massoniana were different in stages, and the individuals of different sizes in the population had different responses to intraspecific competition.
Key words:Ecological restoration; Intraspecific competition; Pinus massoniana; Mortality of the population
種群动态作为种群生态学的核心问题[1-2],现有研究多从种群的生死过程对其进行讨论。研究证实个体死亡对种群动态产生的影响不可忽视[3],但在目前的种群动态分析中多从内源性的影响因素对个体的死亡机制进行讨论[4-5],忽略了外源性因素。作为种群发展过程中长期存在且不断调整的生态关系,竞争来源不仅包括内源性的种内竞争,还包括外源性的种间竞争[6-7],是影响种群动态变化的不可忽视的因素,但其对个体死亡率的影响机制尚不明确。
竞争普遍存在于林木之间,影响着个体的存活[8-9]。一般来说,竞争强度的增加会提升个体死亡的风险[10-11],而Pillet等[12]研究指出竞争强度的增强会造成死亡率的降低,同时也有研究认为竞争对于种群死亡率的影响效果很低甚至没有影响[13]。由此可见,竞争对于种群死亡率的影响复杂且多变。同时还有研究发现,伴随竞争强度的增加,老龄个体则相对于幼龄个体表现出更高的死亡率[14],这表明种群内不同大小个体对竞争的响应具有差异[13,15]。为明确竞争对马尾松个体死亡率的影响机制,本研究于福建省长汀县河田镇设立3处不同恢复水平的马尾松样地。该地区从20世纪80年代起以马尾松为先锋树种建立了大量的人工林[16],历经40年的生态恢复,当地马尾松林呈现出不同的恢复水平。本研究量化3个样地内马尾松个体所受的竞争强度,并构建马尾松种群死亡率函数,进而分析个体所受种内竞争的差异对种群个体死亡率产生的影响,以期为该地区的生态恢复提供理论指导依据。
1 材料与方法
1.1 研究区域
研究区位于中国福建省长汀县(116°00′45″~116°39′20″E,25°18′40″~26°02′05″N),气候类型为亚热带海洋性季风气候,年平均气温18.3℃,年均降水量1685.6 mm,降水主要集中于4月至6月份。由于人类干扰,20世纪时当地植被被大规模破坏,该地区形成了中国南方最典型的红壤水土流失区。经过40多年的生态恢复,目前植被主要为以马尾松为主的纯林和针阔混交林[17]。
1.2 样地设置与野外调查
试验在研究区设置了3处10 hm2永久性样地,分别为伯湖(BH)、马坑(MK)、水东坊(SDF),样地生态恢复水平依次提高。在每个样地内设立30 m×30 m的样方,同时对样方内的马尾松个体进行每木调查,测量树木的基径大小。调查过程中对所有个体挂牌标记,记录其坐标位置。随后每年对挂牌个体重复测量并记录存活状况,调查当年发现个体死亡证据或连续两年未找到则判定树木死亡。样地初次调查时间为2017年7月,并于2018年7月、2019年7月及2020年8月对样地再次进行调查。
1.3 数据分析
1.3.1 Hegyi竞争指数 本研究中使用Hegyi竞争指数[18]对样地内马尾松个体所受的竞争强度进行量化。
式中I表示每个马尾松个体的Hegyi竞争指数,n为竞争木株数,di为竞争木基径,dj为对象木基径,distij为竞争木到对象木的距离。I值越大表示对象木种群受到的竞争越激烈。
本研究采用固定样圆[19]的方法筛选对象木所对应的竞争木,计算样圆范围内所有竞争木对对象木的竞争强度,样圆的半径根据经验设置为5 m。为去除边缘效应,在每个样方内设立宽度为5 m的缓冲区域,后续数据处理及模型拟合基于去除边缘效应后的样地内马尾松个体数据。
1.3.2 死亡率函数 为了验证竞争影响下马尾松种群死亡率的差异,本研究将个体大小(x,att)与未来个体的大小(y,att+1)及个体存活状态建立回归方程。由于马尾松个体的存活状态的是一个离散时间,判别的基准仅存在0(存活)或1(死亡),故死亡率函数的构建是基于二项式分布GLMM构建。
对3个不同恢复水平的样地,本研究将个体大小与竞争强度作为固定效应,年份作为随机效应,构建死亡函数。同时也构建了忽略竞争影响,仅将个体大小作为固定效应、年份作为随机效应的死亡率函数。不同样地的死亡率函数模型选择基于赤池信息准则(AIC),较低的AIC值表示模型更为可靠。
1.4 数据统计
试验数据处理与模型构建采用r语言开源统计软件R 4.0.3进行。
2 结果与分析
2.1 不同恢复水平样地马尾松个体大小及其競争指数
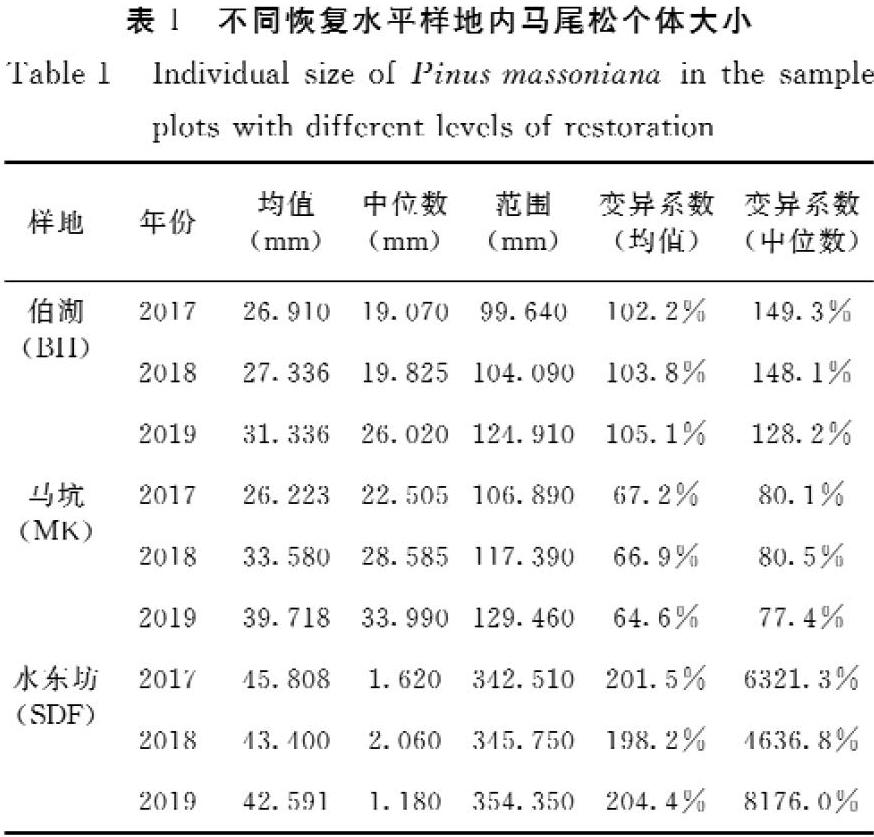
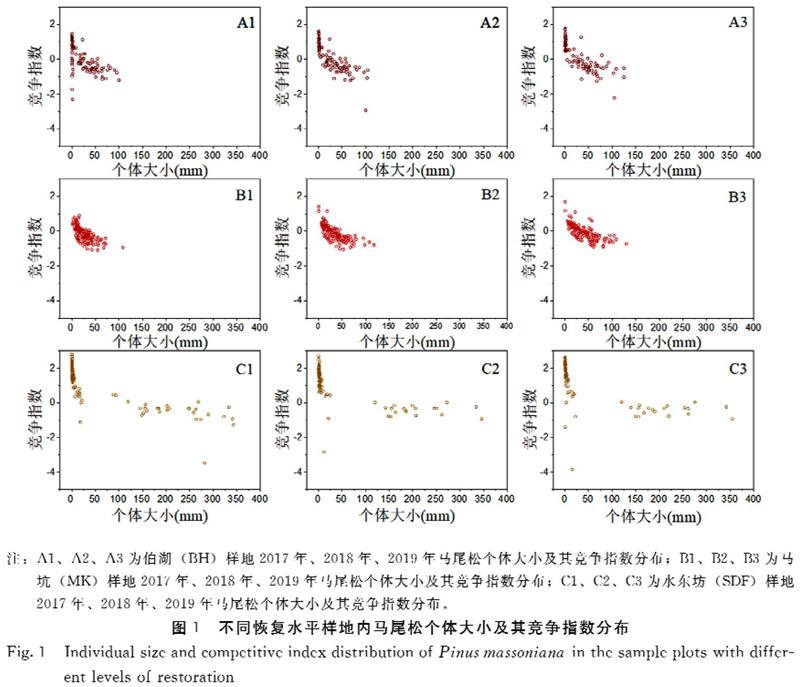
对3个不同恢复水平样地内的个体大小(龄级)的分布情况进行了分析,通过变异系数体现样地间大小结构的差异。结果(表1,图1)表明,3个样地的马尾松个体大小结构差异显著,但同一样地在年份之间差异并不显著。相较于伯湖(BH)与水东坊(SDF)样地,马坑(MK)样地内马尾松个体大小分布最为集中,平均大小为33.17 mm,幼苗数量少。伯湖(BH)样地内个体大小分布较为集中,马尾松个体大小平均为28.53 mm,幼苗数量多。而水东坊(SDF)样地内马尾松个体大小分布最为分散,幼苗较多,且相较于其他样地存在较多大个体的马尾松(基径>150 mm)。
本研究计算了各样地内个体竞争指数,并对其变异系数进行了计算。结果表明(图1,表2),在3个样地中,水东坊(SDF)样地的马尾松种内竞争最为激烈,其次为伯湖(BH)样地,马坑(MK)样地的马尾松种群种内竞争最低,其中伯湖(BH)样地内个体的竞争指数变异系数最高。3个样地的马尾松种群均体现出随着大小的增加,个体的竞争指数逐渐降低的趋势。同时研究还发现伯湖(BH)与马坑(MK)样地内的马尾松个体平均竞争指数在3年内呈增加趋势。
2.2 不同恢复水平样地马尾松种群死亡率函数
通过对3个样地的马尾松种群死亡率函数拟合结果发现,纳入竞争的死亡率函数在伯湖(BH)和马坑(MK)样地拟合程度高,而在水东坊(SDF)样地拟合度较差(表3)。筛选出最优的死亡率函数后,对3个样地的死亡率函数作图(图2)。由图2可知,水东坊(SDF)的幼苗死亡的风险最高,其次为伯湖(BH)样地,马坑(MK)样地幼苗的死亡率较低。同时研究还发现,3个样地中非幼苗的死亡风险几乎为0。
3 讨论与结论
种群死亡率受到外界环境变化及个体间的相互作用共同影响[20]。研究表明种内竞争可以通过影响种群内的资源分配直接影响树木的存活状态[13,21]。特别在生态恢复的先锋阶段,由于生境的限制,先锋种作为优势种在群落占据绝对优势地位,其个体所受竞争作用的主要来源为种内竞争[7]。但由于非对称竞争影响,不同大小的个体对种内竞争的响应不同[22]。有研究认为处于同一生境的种群,幼苗的存活相较于大个体对环境的变化具有更高的敏感性[11],光照、水热等条件对于幼苗的存活具有关键作用[23-25],幼苗更易受种内竞争的影响。本研究研究结果支持幼苗在竞争影响下死亡率更高的观点,研究结果发现在生态恢复的先锋阶段,马尾松种内竞争强度的增加主要对马尾松幼苗的存活影响更为显著。研究表明伴随种内竞争强度的增加,马尾松幼苗的死亡风险提升。
有研究表明由于生境资源的限制,植株幼苗在竞争影响下无法获得维持生存的资源,易使幼苗因种内竞争而死亡[26]。通常来说,伴随个体间的竞争强度增加,处于竞争劣势的个体的死亡风险可能会因资源的限制而提高[27]。但本研究发现,种内竞争对于先锋种马尾松个体存活的影响在不同的恢复阶段具有差异,种内竞争对个体死亡率的影响在生态恢复的先锋阶段早期与中期更为显著,而其对个体死亡率的影响在先锋阶段的后期阶段较小甚至可以忽略,这是受不同生态恢复阶段马尾松种群内竞争指数变异性及个体大小的分布差异所影响的。竞争强度的量化基于个体大小与个体间的距离[18],因此个体所受竞争强度的与其竞争单元内的竞争木个体大小及距离有关。调查发现先锋阶段后期马尾松幼苗数量较多,个体大小分布的变异系数高,幼苗的种内竞争来源主要为大个体马尾松,但其竞争指数的变异性较低,可能是因为样地内个体大小差异较大,且样地内的幼苗聚集且与大个体间的距离较远所导致的,该阶段种内竞争并不是导致该阶段幼苗死亡的主要原因,林下植被的生长或其他物种带来的种间竞争可能对幼苗的死亡贡献更高[10-11,28]。而在先锋阶段的早期与中期,马尾松个体普遍偏小,且与大个体聚集,故幼苗更容易因种内竞争所带来的资源限制而死亡,因此作为马尾松种群内个体死亡的主要来源,幼苗的死亡是导致种群死亡率提高的主要原因,因此在此阶段种内竞争对马尾松种群的死亡函数产生了显著的影响。
种内竞争作为影响种群动态的重要调节因素,影响了生态恢复先锋阶段不同时期的种群死亡率。本研究发现生态恢复先锋阶段的早期与中期,马尾松幼苗由于大个体马尾松带来的资源竞争死亡风险增加,并显著影响了种群的死亡率。而在先锋阶段后期,种内竞争并不是影响马尾松死亡率变化的主要因素。本研究结果可以帮助分析不同生态恢复水平下马尾松个体的死亡风险的来源,有助于为生态恢复过程中的森林管理提供理论依据。
参考文献:
[1]董鸣.生态学透视:种群生态学[M].北京:科学出版社,2016.
[2]江洪.云杉种群生态学[M].北京:中国林业出版社,1992.
[3]BUGMANN H,SEIDL R,HARTIG F,et al.Tree mortality submodels drive simulated long-term forest dynamics:assessing 15 models from the stand to global scale[J].Ecosphere,2019,10(2):e02616.
[4]杨贤宇,李守中,宋铁燕,等.长汀侵蚀退化区砍伐迹地马尾松种群动态[J].生态学报,2018,38(9):3175-3182.
[5]YANG X Y,LI S Z,LI S L,et al.Demographic strategies of a dominant tree species in response to logging in a degraded subtropical forest in Southeast China[J].Annals of Forest Science,2018,75(3):84.
[6]林育真,付荣恕.生态学[M].北京:科学出版社,2011.
[7]王从容,李守中,杨贤宇,等.亚热带红壤侵蚀退化区不同恢复水平生境内先锋树种竞争特征[J].应用与环境生物学报,2019,25(2):239-245.
[8]CANHAM C D,PAPAIK M J,MARIA U,et al.Neighborhood analyses of canopy tree competition along environmental gradients in New England forests[J].Ecological Applications,2006(16):540-554.
[9]ANNING A K,MCCARTHY B C.Competition,size and age affect tree growth response to fuel reduction treatments in mixed-oak forests of Ohio[J].Forest Ecology and Management,2013,307:74-83.
[10]URIARTE M,CANHAM C D,THOMPSON J,et al.A neighborhood analysis of tree growth and survival in a hurricane-driven tropical forest[J].Ecological Monographs,2004,74:591-614.
[11]LASKY J R,BNDICTE B,MUSCARELLA R,et al.Ontogenetic shifts in trait-mediated mechanisms of plant community assembly[J].Ecology,2015,96:2157-2169.
[12]PILLET M,JOETZJER E,BELMIN C,et al.Disentangling competitive vs.climatic drivers of tropical forest mortality[J].Journal of Ecology,2018,106(3):1165-1179.
[13]ROZENDAAL D M A,PHILLIPS O L,LWEIS S L,et al.Competition influences tree growth,but not mortality,across environmental gradients in Amazonia and tropical Africa[J].Ecology,2020,101(7):e03052.
[14]QUARLES B M,ROACH D A.Ageing in an Herbaceous Plant:Increases in Mortality and Decreases in Physiology and Seed Mass[J].Journal of Ecology,2019,107(3):1409-1418.
[15]TREDENNICK A T,TELLER B,ADLER P B,et al.Size-by-environment interactions:a neglected dimension of species′ responses to environmental variation[J].Ecology Letters,2018,21(12):1757-1770.
[16]謝锦升,杨玉盛,陈光水,等.封禁管理对严重退化群落养分循环与能量的影响[J].山地学报,2002,20(3):325-330.
[17]姚雄,余坤勇,刘健,等.南方水土流失严重区的生态脆弱性时空演变[J].应用生态学报,2016,27(3):735-745.
[18]李际平,房晓娜,封尧,等.基于加权Voronoi图的林木竞争指数[J].北京林业大学学报,2015,37(3):61-68.
[19]李清雪,李钢铁,王永胜,等.浑善达克沙地不同立地类型榆树种内竞争分析[J].内蒙古农业大学学报(自然科学版),2010,31(4):304-308.
[20]程越,林旻,杨刚,等.基于个体植物生长模型的森林动态演替模拟[J].北京林业大学学报,2017,39(6):96-106.
[21]TSKSHASHI K.Effects of altitude and competition on growth and mortality of the conifer Abies sachalinensis[J].Ecological Research,2010,25(4):801-812.
[22]王平,王天慧,周道玮,等.植物地上竞争与地下竞争研究进展[J].生态学报,2007(8):3489-3499.
[23]喻方圆,徐锡增,ROBERT D G.水分和热胁迫对5种苗木生长及生物量的影响[J].南京林业大学学报(自然科学版),2013,27(4):10-14.
[24]HAN W P,DING G J,BAO B.Physiological and ecological responses of Pinus massoniana seedling from different provenances to drought stress[J].J Cent S Univ For Technol,2012,32:25-29.
[25]MA L,LIAN J Y,LIN G J,et al.Forest dynamics and its driving forces of sub-tropical forest in South China[J].Scientific Reports,2016(6):22561.
[26]彭闪江,黄忠良,彭少麟,等.植物天然更新过程中种子和幼苗死亡的影响因素[J].广西植物,2004(2):113-121,124.
[27]潘磊磊,KWON S,刘艳书,等.沙地樟子松天然林南缘分布区林木竞争、空间格局及其更新特征[J].生态学报,2019,39(10):3687-3699.
[28]TINGSTAD L,OLSEN S L,KLANDERUD K,et al.Temperature,precipitation and biotic interactions as determinants of tree seedling recruitment across the tree line ecotone[J].Oecologia,2016,180(3):917-918.
(責任编辑:林玲娜)
猜你喜欢
南方农业·中旬(2021年5期)2021-10-25
中学生百科·小文艺(2020年8期)2020-01-13
山地农业生物学报(2019年1期)2019-09-10
建筑建材装饰(2016年13期)2017-01-04
环球人文地理·评论版(2016年5期)2017-01-03
今传媒(2016年9期)2016-10-15
