
红景天苷对抑郁模型大鼠炎症反应和神经细胞凋亡的抑制作用及其机制*
2021-05-26 05:19韩亚琼李涛谷争陈永新
医药导报 2021年5期
韩亚琼,李涛,谷争,陈永新
(新乡医学院第二附属医院药学部,新乡 453002)
抑郁症具有患病率高、致残率高、复发率高、检出率低、就诊率低、治愈率低等特点,可能将成为全球第二大杀手[1]。目前主要用于抑郁症治疗的药物为选择性5-羟色胺再摄取抑制剂和三环类抗抑郁药等化学药物,但起效慢、易产生药物依赖性、不良反应大[2]。红景天苷是药用植物红景天的主要活性成分,是一种苯乙醇类化合物,具有抗炎、抗氧化、抗癌、抗衰老、保肝等生理活性[3]。研究表明,红景天苷对阿尔茨海默病、帕金森病、亨廷顿病、卒中、抑郁、焦虑等具有潜在的治疗作用,是一种高效、低毒的治疗药物[4]。然而,红景天苷对抑郁症影响的分子机制尚不清楚,进一步研究红景天苷对抑郁症影响的分子机制具有重要意义。炎症反应和神经细胞凋亡在抑郁症的发生中具有重要作用[5],本研究探讨红景天苷对慢性不可预测的轻度压力(chronic unpredictable mild stress,CUMS)抑郁模型大鼠炎症反应和神经细胞凋亡的影响及其机制,以期为抑郁症的预防和治疗提供参考。
1 材料与方法
1.1药品与试剂红景天苷(甄准生物有限公司,含量≥98%,批号:ES-4943S),氟西汀(常州四药制药有限公司,批号:100513-201602),H-89(美国Abcam公司,批号:ab143787),白细胞介素(IL)-1β、IL-6、肿瘤坏死因子(TNF-α)酶联免疫吸附测定(ELISA)试剂盒(上海联硕生物刻科技有限公司,批号:ml037361、ml059476、ml002859),环磷酸腺苷(cyclic adenosine monophosphate,cAMP)ELISA试剂盒(上海谷研实业有限公司,批号:GOY-2242),RIPA裂解缓冲液(南京海克尔生物科技有限公司,批号:SBJ-0999),二喹啉甲酸(bicinchoninic acid,BCA)试剂盒(上海易色医疗科技有限公司,批号:BC201),cAMP依赖的蛋白激酶催化亚基-Ⅱα(cAMP-dependent protein kinase catalytic subunit-Ⅱα,PKAc-Ⅱα)抗体(上海基因生物技术有限公司,批号:5842S),磷酸化环磷腺苷效应元件结合蛋白(phosphorylated cAMP-response element binding protein,p-CREB)抗体,CREB抗体(美国Abcam公司,批号:ab32096、ab31387),山羊抗兔二抗(上海康培科技有限公司,批号:C86-SSA004)。
1.2实验动物清洁级雄性SD 大鼠 150 只,体质量(200±10)g,购自四川夏派森医药科技有限公司,动物生产许可证号:SYXK(川)2017-203。饲养环境温度(24±2)℃,相对湿度约50%,12/12 h光暗交替,大鼠在实验前适应性喂养1周。
1.3分组、CUMS模型构建及给药实验1:将动物采用随机数字表法分为正常对照组、模型对照组、氟西汀组和红景天苷小、中、大剂量组,每组15只;实验2:将动物采用随机数字表法分为模型对照组、H-89组、红景天大剂量组、H-89+红景天大剂量组,每组15只。CUMS抑郁模型构建[6]:将大鼠置单独笼中,每天采用随机数字表法接受一次以下模式的刺激:24 h笼子倾斜(45 °),2 h噪音(92 dB,92 Hz),夜间照明,4 ℃冷游5 min,湿垫料24 h,尾捏1 min(距离尾部1 cm),24 h水和食物剥夺,5 min热环境(45 ℃)。同一刺激不连续进行。正常对照组大鼠在单独的笼中,不受任何刺激。给药:CUMS抑郁模型的第6~12周,红景天苷小、中、大剂量组大鼠分别每天一次腹腔注射红景天苷(溶剂为0.9%氯化钠溶液)12.5,25,50 mg·kg-1[7],氟西汀组大鼠每天一次腹腔注射氟西汀(溶剂为0.9%氯化钠溶液)5 mg·kg-1[8],正常对照组和模型对照组大鼠注射等量0.9%氯化钠溶液,H-89组大鼠每天一次鞘内注射H-89(溶剂为0.9%氯化钠溶液)2 mg·kg-1,H-89+红景天大剂量组大鼠每天一次鞘内注射2 mg·kg-1H-89[9],并腹腔注射红景天苷50 mg·kg-1。
1.4体质量观察和蔗糖偏好实验实验过程中,记录大鼠体质量和蔗糖偏好。蔗糖偏好实验:第1天,给每只大鼠2瓶1%蔗糖水。第2天,给每只大鼠1瓶水和1瓶1%蔗糖水。每2 h更换一次瓶子。训练后,禁食水处理24 h。然后给每只大鼠1瓶水和1瓶1%蔗糖水,预先称体质量,1 h后测量每瓶的消耗量。大鼠的蔗糖偏好率(%)=蔗糖消耗量/(蔗糖消耗量+水消耗量)×100%。
1.5旷场实验在强迫游泳实验之前,将大鼠单独放置在长100 cm、宽100 cm、深50 cm的箱子中5 min。Med旷场视频追踪系统分析5 min内大鼠行进距离、行动路线交叉次数及站立的次数。在测试之间用75%乙醇彻底清洁箱子的底面。
1.6强迫游泳测试在圆柱形树脂玻璃游泳装置(高60 cm、直径20 cm)中加入(24±2)℃水至30 cm处。将每只大鼠单独放入强制游泳装置中6 min,用安装在游泳装置上方的摄像机记录大鼠行为。然后将大鼠从游泳装置中取出并用毛巾擦干,然后返回笼中。记录最后4 min内的大鼠不动的时间。
1.7莫里斯(Morris)水迷宫Morris水迷宫装置由高60 cm、直径150 cm的黑色圆形池组成,分为4个视觉象限,加(24±2)℃满水。将直径10 cm的圆形平台放置在第三象限的中心,水面下方2 cm处。连续4 d,将大鼠放入面向墙壁的池中,每只大鼠依次从4个象限进入,并以相同的顺序进入水里。在60 s内未能找到平台的大鼠被引导并允许在平台上停留10 s。在第5天,将大鼠用于寻找平台的时间记录为潜伏期。在第6天,移除平台并给予大鼠60 s以确定它们学习平台位置的程度,将大鼠置于进行最后一次训练的入口处的水中并允许游泳60 s。记录平台所在象限中位置的次数。
1.8苏木精-伊红(HE)染色麻醉后处死大鼠,分离出脑海马区组织,将脑海马区组织用4%多聚甲醛溶液固定后,常规脱水制成石蜡切片,然后脱蜡、苏木精染色、1%盐酸乙醇分化、0.6%氨溶液返蓝、0.5% 伊红液染色,接着常规脱水,二甲苯透明,中性树胶封片,在400倍显微镜观察神经细胞损伤情况。
1.9流式细胞术将海马区组织裂解并用4 ℃、0.9%氯化钠溶液迅速制成 1:9匀浆液,加入 Annexin-V-FITC和碘化丙啶各5 μL,在黑暗的房间里孵化15 min,然后用流式细胞仪分析细胞凋亡情况。
1.10ELISA实验将海马区组织用4 ℃、0.9%氯化钠溶液迅速制成 1:9匀浆液,3000 r·min-1离心15 min,取上清液按照ELISA试剂盒说明书操作检测cAMP、IL-6、IL-1β、TNF-α的含量。
1.11免疫印迹法检测提取海马组织中总蛋白,采用BCA蛋白检测试剂盒进行定量。将蛋白质样品25 μg用12%十二烷基硫酸钠(sodium dodecyl sulfate,SDS)凝胶电泳分离,并转移到聚偏二氟乙烯膜,用5% 牛血清白蛋白封闭过夜后,用兔抗大鼠单克隆抗体(PKAc-II 1:1000、P-CREB 1:1000、CREB 1:1000),4 ℃摇床上2 h,然后与山羊抗兔二抗(1:1000)在4 ℃的摇床上摇1 h,电化学曝光,以β-actin为内参,使用Quantity One软件进行分析目标蛋白质的相对表达水平。
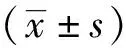
2 结果
2.1红景天苷改善CUMS抑郁模型大鼠体质量见图1。随着给药时间增加,各组大鼠体质量差异逐渐增大。在12周时,正常对照组,模型对照组,红景天小、中、大剂量组,氟西汀组大鼠的体质量分别是(376.17±23.87),(286.34±24.31),(295.64±26.17),(332.56±25.52),(354.32±22.64),(361.77±29.12)g,差异有统计学意义(F=82.43,P<0.01),其中,与正常对照组比较,模型对照组大鼠体质量明显减轻(P<0.01);与模型对照组比较,红景天苷小剂量组大鼠体质量无明显变化(P>0.05),红景天苷中剂量组体质量增加(P<0.05),大剂量组和氟西汀组体质量均明显增加(P<0.01)。

A.正常对照组;B.模型对照组;C.红景天苷小剂量组;D.红景天苷中剂量组;E.红景天苷大剂量组;F.氟西汀组。①与正常对照组比较,P<0.01;②与模型对照组比较,P<0.05;③与模型对照组比较,P<0.01。
2.2红景天苷改善CUMS抑郁模型大鼠快感缺乏见图2。在第12周时,正常对照组,模型对照组,红景天小、中、大剂量组,氟西汀组大鼠的蔗糖偏好率分别是(91.67±2.22)%,(52.56±2.17)%,(58.92±2.77)%,(72.38±2.73)%,(86.35±2.84)%,(88.24±2.59)%,差异有统计学意义(F=76.54,P<0.01),其中,与正常对照组比较,模型对照组大鼠蔗糖偏好率明显降低(P<0.01),说明CUMS抑郁模型大鼠快感明显缺乏;与模型对照组比较,红景天苷小剂量组大鼠蔗糖偏好率无明显变化(P>0.05),红景天苷中剂量组蔗糖偏好率均明显升高(P<0.05),红景天苷大剂量组和氟西汀组蔗糖偏好率均明显升高(P<0.01),说明红景天苷能明显恢复CUMS抑郁模型大鼠快感。

A.正常对照组;B.模型对照组;C.红景天苷小剂量组;D.红景天苷中剂量组;E.红景天苷大剂量组;F.氟西汀组。①与正常对照组比较,P<0.01;②与模型对照组比较,P<0.05;③与模型对照组比较,P<0.01。
2.3红景天苷改善CUMS抑郁模型大鼠活动和探索行为见图3。正常对照组、模型对照组,红景天小、中、大剂量组,氟西汀组大鼠的行进距离、运动轨迹交叉次数、站立次数、游泳不动时间均差异有统计学意义(F=97.53,32.15,43.26,66.37,均P<0.01),其中,与正常对照组比较,模型对照组大鼠旷场实验中行进距离、运动轨迹交叉次数、站立次数均明显减少(P<0.01),强迫游泳实验中不动时间明显增多(P<0.01),说明CUMS抑郁模型大鼠活动和探索行为减弱;与模型对照组比较,红景天苷小剂量组大鼠旷场实验中行进距离、运动轨迹交叉次数、站立次数及强迫游泳实验中不动时间均无明显变化,红景天苷中、大剂量组和氟西汀组旷场实验中行进距离、运动轨迹交叉次数、站立次数均明显增加(P<0.05,P<0.01),强迫游泳实验中不动时间明显减少(P<0.05,P<0.01),说明红景天苷能明显增强CUMS抑郁模型大鼠活动和探索行为。

A.正常对照组;B.模型对照组;C.红景天苷小剂量组;D.红景天苷中剂量组;E.红景天苷大剂量组;F.氟西汀组。①与正常对照组比较,P<0.01;②与模型对照组比较,P<0.05;③与模型对照组比较,P<0.01。
2.4红景天苷改善CUMS抑郁模型大鼠学习和认知能力见图4。正常对照组、模型对照组,红景天小、中、大剂量组,氟西汀组大鼠第5 天找到平台所需时间及60 s 内找到平台次数均差异有统计学意义(F=51.24,67.48,P<0.01),其中,与正常对照组比较,模型对照组大鼠找到平台所需时间明显增加(P<0.01),60 s内找到平台次数明显减少(P<0.01),说明CUMS抑郁模型大鼠学习和认知能力减弱;与模型对照组比较,红景天苷小剂量组大鼠找到平台所需时间及60 s内找到平台次数均无明显变化,红景天苷中、大剂量组和氟西汀组找到平台所需时间明显减少(P<0.05,P<0.01),60 s内找到平台次数明显增加(P<0.05,P<0.01),说明红景天苷能明显增强CUMS抑郁模型大鼠学习和认知能力。

A.正常对照组;B.模型对照组;C.红景天苷小剂量组;D.红景天苷中剂量组;E.红景天苷大剂量组;F.氟西汀组。①与正常对照组比较,P<0.05;②与模型对照组比较,P<0.05;③与正常对照组比较,P<0.01;④与模型对照组比较,P<0.01。
2.5红景天苷改善CUMS抑郁模型大鼠神经损伤见图5。正常对照组大鼠海马区组织显微结构正常,神经细胞形态结构正常,排列整齐,细胞质饱满,细胞核清晰;模型对照组神经细胞体积变小,细胞核出现皱缩,存在严重的炎症细胞浸润;与模型对照组比较,红景天苷中、大剂量组和氟西汀组大鼠神经细胞体积变小、细胞核皱缩和炎症细胞浸润情况均有不同程度减轻。

图5 HE染色观察6组大鼠海马区神经细胞损伤(×400)
2.6红景天苷抑制CUMS抑郁模型大鼠神经细胞凋亡正常对照组,模型对照组,红景天小、中、大剂量组,氟西汀组大鼠神经细胞凋亡率分别为(5.85±1.03)%,(16.06±1.98)%,(14.57±1.12)%,(10.46±1.09)%,(7.01±1.07)%,(6.46±1.18)%,差异有统计学意义(F=58.76,P<0.01),其中,模型对照组大鼠神经细胞凋亡率升高至正常对照组约3倍(P<0.01),红景天苷小剂量组与模型对照组比较大鼠神经细胞凋亡率无明显变化(P>0.05),红景天苷中剂量组神经细胞凋亡率下降至模型对照组约2/3(P<0.05),大剂量组和氟西汀组神经细胞凋亡率下降至模型对照组约1/2(P<0.01),说明红景天苷能明显抑制CUMS抑郁模型大鼠神经细胞凋亡。
2.7红景天苷抑制CUMS抑郁模型大鼠炎症反应见图6。正常对照组、模型对照组,红景天小、中、大剂量组,氟西汀组大鼠IL-1β、IL-6、TNF-α含量均差异有统计学意义(F=43.15,37.28,46.52,P<0.01),其中,与正常对照组比较,模型对照组大鼠海马组织中炎症因子IL-1β、IL-6、TNF-α含量明显增加(P<0.01);与模型对照组比较,红景天苷小剂量组大鼠海马组织中炎症因子IL-1β、IL-6、TNF-α含量无明显变化,红景天苷中、大剂量组和氟西汀组海马组织中炎症因子IL-1β、IL-6、TNF-α含量明显减少(P<0.05,P<0.01),说明红景天苷能明显抑制CUMS抑郁模型大鼠炎症反应。

A.正常对照组;B.模型对照组;C.红景天苷小剂量组;D.红景天苷中剂量组;E.红景天苷大剂量组;F.氟西汀组。①与正常对照组比较,P<0.01;②与模型对照组比较,P<0.05;③与模型对照组比较,P<0.01。
2.8红景天苷激活CUMS抑郁模型大鼠cAMP/PKA/CREB信号通路正常对照组、模型对照组,红景天小、中、大剂量组,氟西汀组大鼠海马组织cAMP含量分别是(1.98±0.23),(1.42±0.21),(1.49±0.19),(1.71±0.22),(1.92±0.24),(1.54±0.18)pmol·mg-1,PKAc-Ⅱα/β-actin和p-CREB/CREB蛋白相对表达水平见图7。各组大鼠cAMP含量及PKAc-Ⅱα/β-actin和p-CREB/CREB蛋白均差异有统计学意义(F=37.24,26.49,24.87,均P<0.01),其中,与正常对照组比较,模型对照组大鼠海马组织中cAMP含量明显减少(P<0.01),PKAc-Ⅱα/β-actin和p-CREB/CREB蛋白表达明显下调;与模型对照组比较,红景天苷小剂量组和氟西汀组大鼠海马组织中cAMP含量及PKAc-Ⅱα/β-actin和p-CREB/CREB蛋白表达无明显变化,红景天苷中、大剂量组海马组织中cAMP含量明显增加(P<0.05,P<0.01),PKAc-Ⅱα/β-actin和p-CREB/CREB蛋白表达明显上调(P<0.05,P<0.01),说明红景天苷能明显激活CUMS抑郁模型大鼠cAMP/PKA/CREB信号通路。

A.正常对照组;B.模型对照组;C.红景天苷小剂量组;D.红景天苷中剂量组;E.红景天苷大剂量组;F.氟西汀组。①与正常对照组比较,P<0.01;②与模型对照组比较,P<0.05;③与模型对照组比较,P<0.01。
2.9红景天苷通过cAMP/PKA/CREB信号通路抑制CUMS抑郁模型大鼠炎症反应和神经细胞凋亡见图8。模型对照组、H-89组、红景天苷大剂量组、H-89+红景天苷大剂量组大鼠体质量、蔗糖偏好率、行进距离、交叉次数、站立次数、不动时间、平台所需时间、到达平台次数、海马区PKAc-Ⅱα/β-actin和p-CREB/CREB蛋白表达、神经细胞凋亡率、海马区IL-1β、IL-6、TNF-α含量均差异有统计学意义(F=80.24,73.14,95.47,36.22,45.13,68.42,49.67,63.44,28.57,25.76,56.54,44.75,38.67,47.22,P<0.01);在给予大剂量红景天苷的基础上,给予H-89能够逆转大剂量红景天苷引起的CUMS抑郁模型大鼠的改变(P<0.01);表明红景天苷可能通过cAMP/PKA/CREB信号通路抑制CUMS抑郁模型大鼠炎症反应和神经细胞凋亡。

A.模型对照组;B.H-89组;C.红景天苷大剂量组;D.H-89+红景天苷大剂量组。①与模型对照组比较,P<0.05;②与红景天苷大剂量组比较,P<0.05;③与模型对照组比较,P<0.01;④与红景天苷大剂量组比较,P<0.01。
3 讨论
抑郁症是一种常见的精神疾病,易导致心理和身体受损,自杀率高,是一个广受关注的公共卫生问题。虽然如选择性5-羟色胺再摄取抑制剂、单胺氧化酶抑制剂等抗抑郁药对抑郁症具有一定效果,但存在起效慢、反应率低、对器官的毒性作用和药物耐药性等缺点[10]。传统中草药中提取的天然化合物与传统的抗抑郁药物比较,具有较少的不良反应(如失眠、便秘、心血管功能和代谢紊乱),是抑郁症的有效补充和替代药物。
慢性不可预见性温和刺激动物抑郁症与人类抑郁症病因相似,是一种经过广泛使用和充分验证、并得到国际认可的抑郁症动物造模方法[11]。CUMS动物暴露于不同类型的压力,模仿不可预测的压力性生活事件,并引起类似认知、生理、情绪变化的抑郁样行为[12]。本研究显示,CUMS大鼠6周后蔗糖偏好率明显减少,说明快感缺失,这是人类抑郁症的核心症状,也表明大鼠抑郁模型已成功建立。给予红景天苷,蔗糖偏好率明显上升,说明红景天苷可改善CUMS抑郁模型大鼠快感缺乏症。
旷场实验和强迫游泳实验可以反映大鼠活动和探索能力。旷场实验中行走的总距离反映了新环境中的活动及情绪的变化;活动路线交叉和站立的次数是动物探索周围环境的行为的指数,反映安全感的变化;强迫游泳中不动时间是“行为绝望”的表现;Morris水迷宫实验可以反映大鼠学习和认知能力[13]。本研究显示,给予红景天苷后,CUMS大鼠在旷场实验中行进距离、运动轨迹交叉次数、站立次数均明显增加,而强迫游泳实验中不动时间明显减少,找到平台所需时间明显减少,60 s内找到平台次数明显增加,说明红景天苷能明显增强CUMS抑郁模型大鼠活动探索行为及学习认知能力。
海马在学习、记忆、情绪、内分泌及内脏活动中起重要作用,也是介导应激反应的重要脑区[14-15]。抑郁症患者存在海马损伤,会引起海马区神经元的凋亡增加。本研究发现,给予红景天苷后,CUMS大鼠海马神经损伤减轻,神经细胞凋亡率下降,说明红景天苷能够抑制CUMS大鼠海马神经损伤及神经细胞凋亡。大脑中炎症细胞因子增加在抑郁症的发病机制中起着至关重要的作用,因此,促炎细胞因子可能是治疗抑郁症的明显目标[16]。临床研究发现包括IL-1β,IL-6和TNF-α等促炎症细胞因子在抑郁症期间显著升高[17]。本研究发现,给予红景天苷后,CUMS大鼠IL-1β、IL-6、TNF-α含量显著减少,说明红景天苷能够抑制CUMS大鼠海马区炎症反应。
cAMP信号传导途径是参与神经干细胞的神经发生、学习和记忆能力的细胞内信号传导途径之一。抑郁破坏cAMP/PKA/CREB信号传导途径。cAMP的水平受cAMP依赖性PKA的活性影响,PKA是CREB的上游激活剂,是与抑郁症和抗抑郁过程有关的研究最广泛的转录因子之一[18]。CREB信号传导是通过调节增加突触和神经可塑性的基因来促进突触和神经可塑性的关键因素。p-CREB可能是一种指示抗抑郁药治疗的有效性的分子标志物[19]。cAMP/PKA/CREB信号通路的系统性扰动可诱导抑郁症一系列神经病理反应,包括区域大脑活动异常、突触功能改变和神经发生受损等[20]。本研究发现,给予红景天苷后,CUMS大鼠海马组织中cAMP含量明显增加,PKAc-Ⅱα/β-actin和p-CREB/CREB蛋白表达明显上调,说明红景天苷能明显激活CUMS抑郁模型大鼠cAMP/PKA/CREB信号通路。添加PKA抑制剂H-89后,能明显逆转红景天苷引起的CUMS大鼠变化,说明红景天苷可能通过cAMP/PKA/CREB信号通路抑制CUMS抑郁模型大鼠炎症反应和神经细胞凋亡。
综上所述,红景天苷可能通过cAMP/PKA/CREB信号通路抑制CUMS抑郁模型大鼠炎症反应和神经细胞凋亡,红景天苷是一种有前景的抑郁症治疗药物,并为以天然产品为重点的抑郁症药物的治疗开发奠定了基础。未来的研究需要在分子和基因水平证实这些机制,并充分阐明红景天苷的抗抑郁作用。
猜你喜欢
今日农业(2022年14期)2022-09-15
中国现代医生(2022年6期)2022-04-23
科学24小时(2022年2期)2022-01-26
保健与生活(2021年22期)2021-11-11
家庭医药(2021年7期)2021-07-23
好孩子画报(2021年5期)2021-06-21
昆明医科大学学报(2021年2期)2021-03-29
——题《圣山系-金巅》
草堂(2020年9期)2020-11-18
中国药房(2018年18期)2018-09-10
心脑血管病防治(2016年6期)2017-01-16
