
舟山群岛东侧海域春秋季主要甲壳类物种的空间生态位分析
2021-07-21 06:10刘坤俞存根许永久江新琴郑基于南京张佩怡蒋巧丽
浙江大学学报(理学版) 2021年4期
刘坤,俞存根,许永久,江新琴,郑基,于南京,张佩怡,蒋巧丽
(浙江海洋大学水产学院,浙江舟山316022)
生态位指生物种群在多维空间中的分布及其与相关种群之间的功能关系,反映了种群资源利用的能力[1]。自GRINNELL[2]首次 对生态位做 出定义以来,生态位理论不断得到深化和完善,并被广泛应用于物种多样性、群落结构演替与种群进化、群落物种积聚等研究[3-4],逐渐成为评价种间和种内关系的重要手段[5]。空间生态位反映了特定区域内生物群落中物种对环境资源的综合利用状况和种间的竞争关系[6],常用于鸟类[7]、昆虫[8]及植物保护[9]等领域的研究。近年来,生态位理论开始用于海洋生态系统生物群落研究,以评估物种对资源的利用状况和物种间的生态关系,如文献[10-12]通过对主要鱼类的生态位分析,揭示了物种在不同尺度上的均匀度和季节变化情况;文献[13]分析了舟山近岸海域主要甲壳类物种的生态位及种间联结,指出物种资源利用的相似性导致种间竞争;文献[14-15]借助生态位理论分析了虾类的种群特征和生存状况,揭示了不同海域虾类的生态位分化、种间竞争及对环境的适应性情况。以上研究从生态学角度更为全面地分析了海洋生物群落的结构和功能。
近年来,舟山群岛东侧海域受过度捕捞和环境污染等因素的影响,鱼类资源衰退,而作为鱼类饵料的甲壳类物种生存空间扩大,大量繁育,在捕捞渔获物中所占比例增加,成为重要的渔业生物资源及捕捞对象[16]。甲壳类作为海洋生态系统能量流动和物质循环过程中的关键物种,其资源状况的变动将影响许多重要的海洋生态过程[13],因此,关注甲壳类物种的群落生态特征,分析其生态位宽度、生态位重叠与分化具有重要意义。本文以2018年在舟山群岛东侧海域进行的2次渔业资源底拖网调查数据为基础,开展甲壳类物种生态位研究,重点分析海域内主要甲壳类物种的空间资源利用状况以及物种间的竞争或共存机制,以期为海域内甲壳类资源的养护和合理利用提供基础资料。
1 材料与方法
1.1 调查及采样方法
本文数据来源于2018年4月(春季)和10月(秋季)在舟山群岛东侧海域开展的渔业资源底拖网调查,调查方法参照《海洋渔业资源调查规范》(SC/T 9403-2012)[17]。调 查 范 围(122°46'54.53″E~124°48'28.31″E,29°10'25.54″N~30°45'0.00″N)及 站位分布见图1,调查船主机功率为1 007 kW,网具规格为1 020目×80 mm,各站位拖曳时间约为1 h,拖速约为3 kn(1 kn=1.852 km·h-1),每个站位拖网所得渔获物全部装入样品袋,编号、记录后,冰鲜保存,带回实验室进行生物学测定和分析,统计各甲壳类物种的尾数和重量(精确到0.1 g),详细记录原始数据并进行标准化处理。2次调查均采用CTD(多功能温盐深仪)同步采集每个站位的水温、盐度、水深等环境因子数据。
1.2 数据处理
用相对重要性指数(index of relative importance,IRI)[18]对甲壳类物种进行分类,IRI计算式为
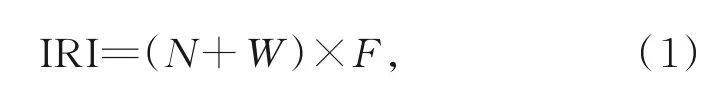
其中,N为某甲壳类个体数占甲壳类物种总数的百分比;W为某甲壳类重量占甲壳类总重量的百分比;F为某甲壳类出现的站位数占调查站位总数的百分比。优势种、常见种参照俞存根等[19]的划分标准,将IRI>1 000的物种定义为优势种,将IRI=100~1 000的物种定义为常见种;参照文献[10,12,14]的相关研究,将IRI>100的物种定义为主要甲壳类。
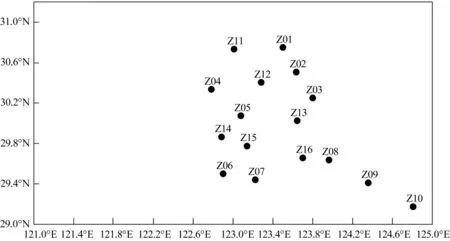
图1 渔业资源调查站位Fig.1 Survey stations of fishery resources
用Shannon指数[20]测定生态位宽度:
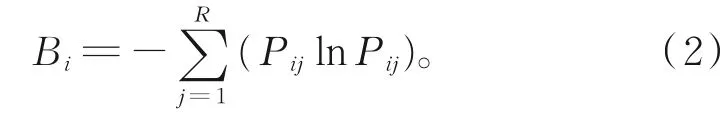
用Pianka公式[21]测定生态位重叠值:
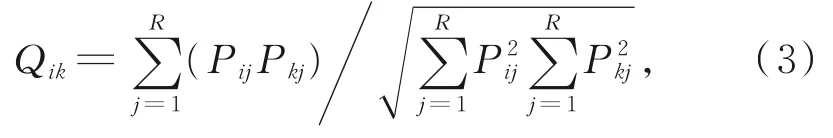
其中,R为资源状态(调查站位)总数;Pij和Pkj分别表示在j资源状态(调查站位)下物种i和物种k的个体数占该物种总数的百分比;Bi表示生态位宽度,取值范围为[0,R];Qik表示生态位重叠值,取值范围为[0,1][22],当Qik>0.3时,重叠有意义,当Qik>0.6时,显著重叠[23]。
用种间竞争系数[24]测定主要甲壳类物种空间维度的种间竞争:
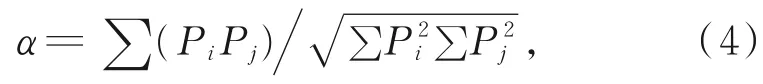
其中,α表示种间竞争系数,Pi和Pj分别表示在同一资源状态下物种i和物种j占物种总数的百分比。
冗余分析(redundancy analysis,RDA)的原理是:将样方和主要研究对象排列在可视化的低维空间,分析物种分布与环境因子之间的关系[25],分析结果可揭示主要甲壳类物种的生态位分化情况。用Canoco 5.0软件分析相关环境因子与主要甲壳类的关系,并绘制排序图,同时采用蒙特卡洛显著性检验(Monte Carlo permutation test)验证环境因子与物种间的显著性。
用Microsoft Excel 2016软件计算IRI和α;用R 3.5.2软件的“spaa”程序包[26]计算Bi和Qik;用Primer 5软件依据空间生态位宽度对主要甲壳类进行聚类分析,用Surfer 11.0软件制作调查站位图。
2 结果与分析
2.1 物种组成
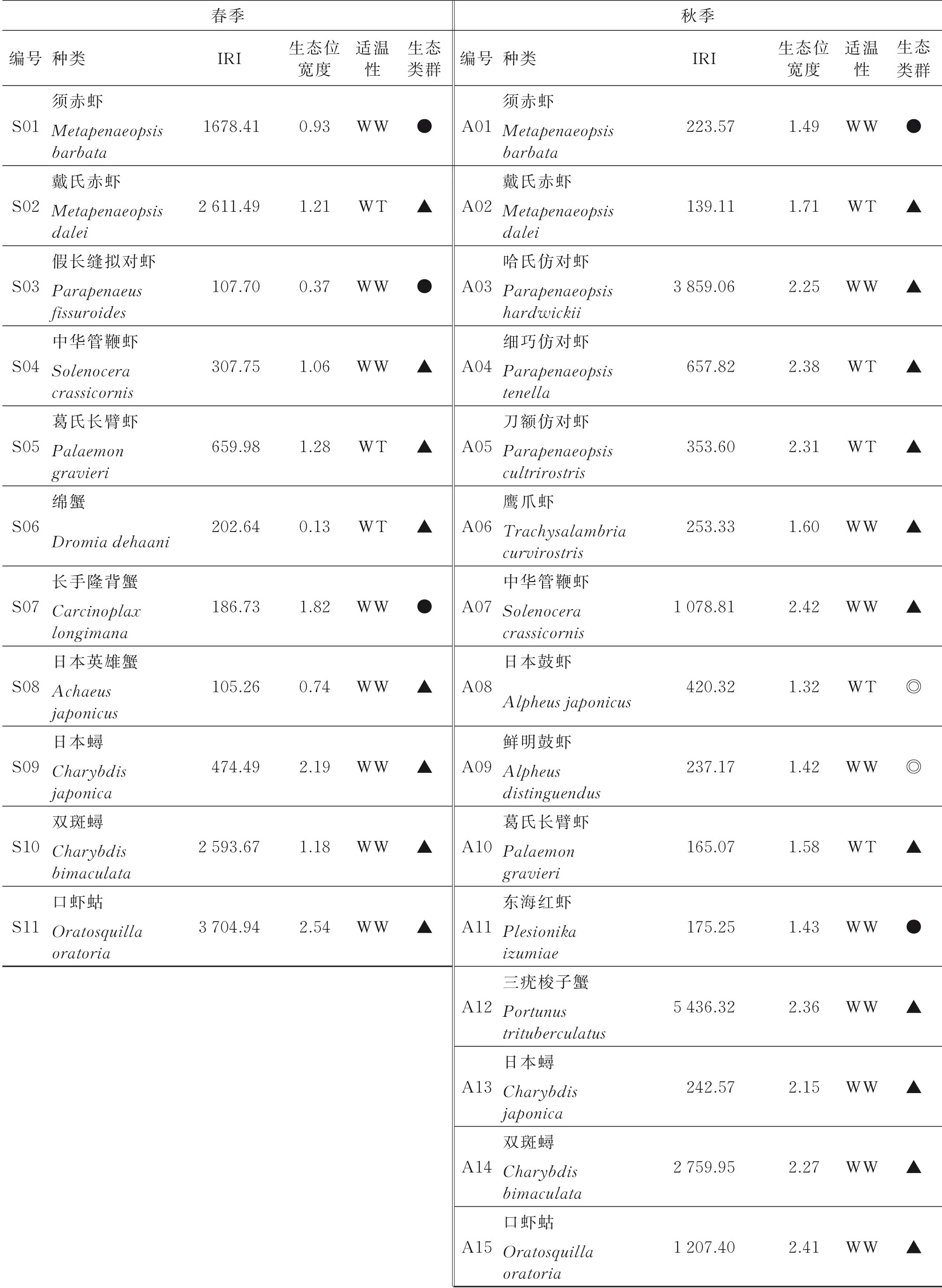
对2个航次的拖网调查渔获物进行了鉴定,分别得到:春季,虾类22种,隶属于9科16属;蟹类29种,隶属于9科18属;虾蛄类1种。秋季,虾类29种,隶属于9科20属;蟹类35种,隶属于11科21属;虾蛄类1种。根据IRI值(见表1)进行分类,本研究中主要甲壳类共19种,其中春季优势种为须赤虾、戴氏赤虾、双斑蟳和口虾蛄4种,秋季优势种为哈氏仿对虾、中华管鞭虾、三疣梭子蟹、双斑蟳和口虾蛄5种,双斑蟳和口虾蛄同为春、秋二季的优势种;春季常见种有7种,秋季常见种有10种。
2.2 生态位宽度
由表1可知,春季主要甲壳类生态位宽度为0.13~2.54,生态位宽度最大的为口虾蛄(2.54),其次为日本蟳(2.19)和长手隆背蟹(1.82),生态位宽度最小的为绵蟹(0.13)。秋季主要甲壳类生态位宽度为1.32~2.42,生态位宽度最大的为中华管鞭虾(2.42),其次为口虾蛄(2.41)和细巧仿对虾(2.38),生态位宽度最小的为日本鼓虾(1.32)。
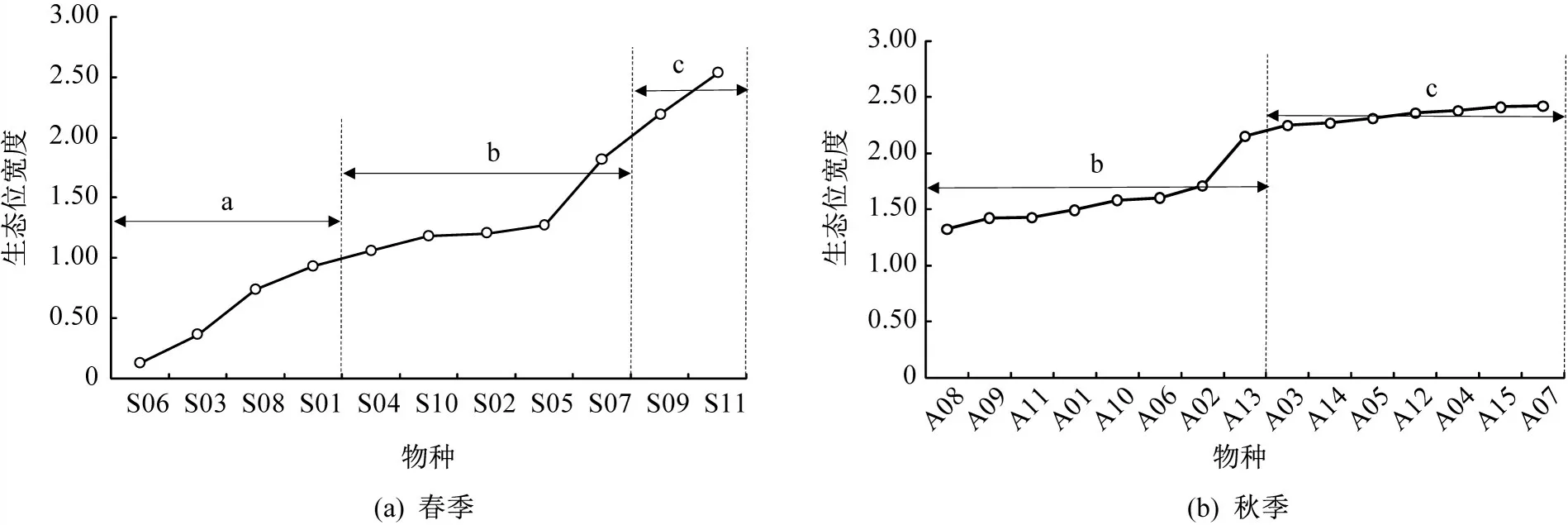
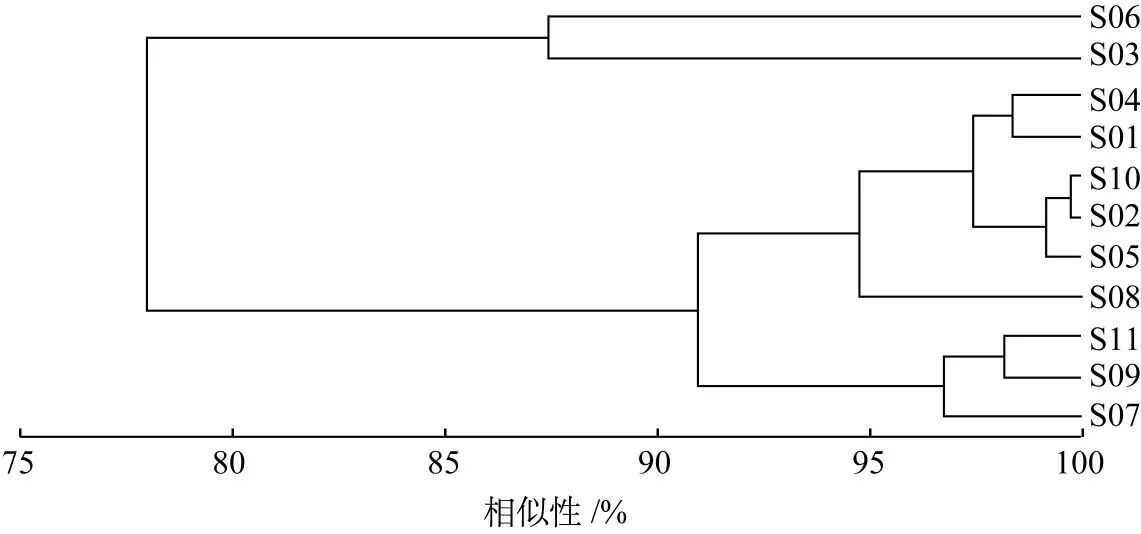
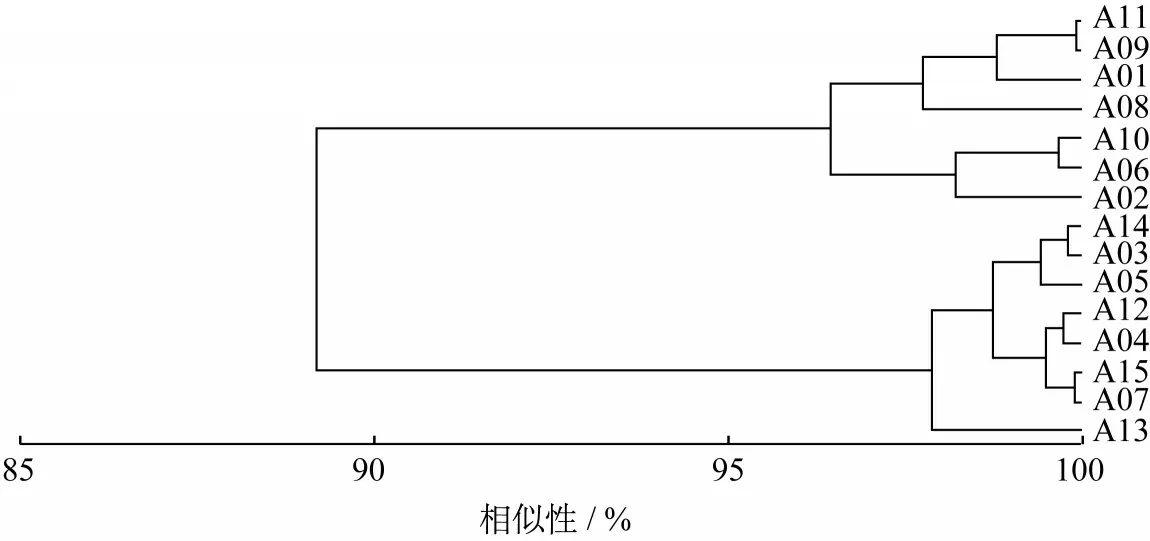
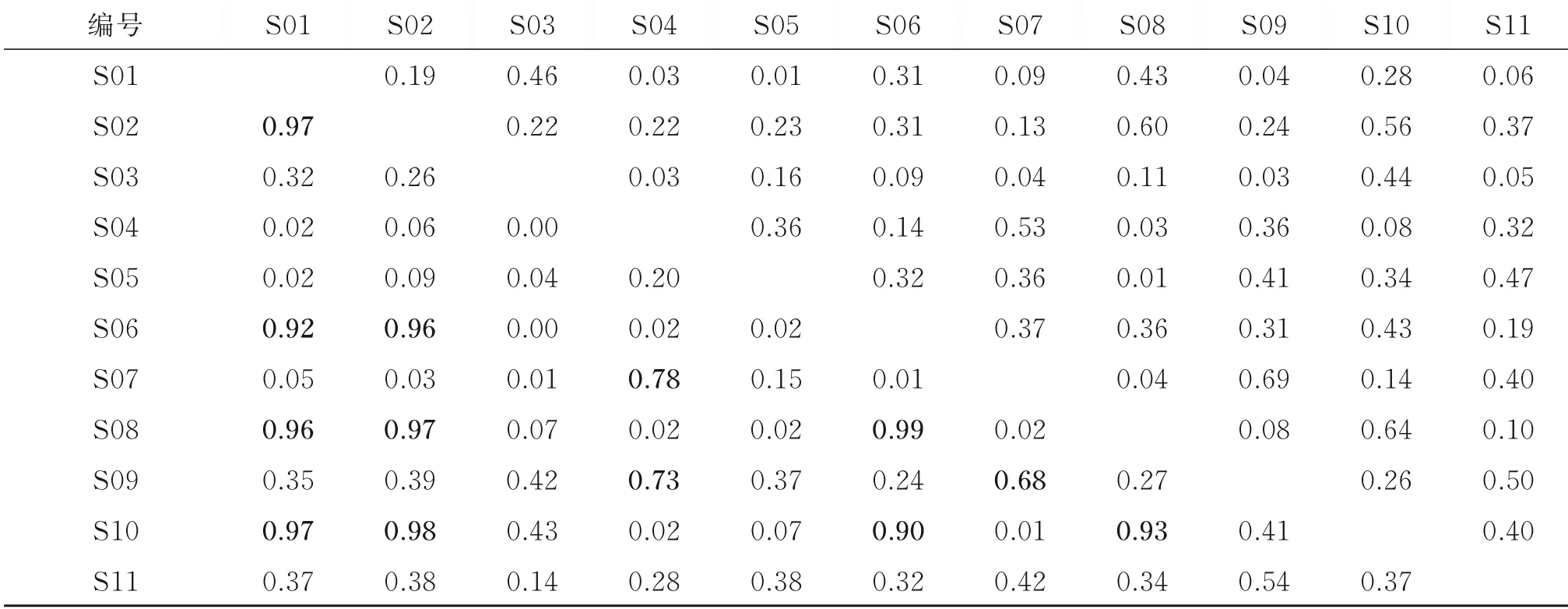
根据生态位宽度将主要甲壳类划分为3组:窄生态位种(0 表1 主要甲壳类IRI及生态位宽度Table 1 The IRIand niche breadth of major crustacean species 注 WW:暖水性;WT:暖温性;●高温高盐;▲广温广盐;◎广温低盐。 图2 主要甲壳类生态位宽度变化Fig.2 Variation of niche breath of major crustacean species a—窄生态位种;b—中生态位种;c—广生态位种;物种种类序号同表1,下同。a-narrow niche species;b-medium niche species;c-wide niche species;The sequence code of species was the same as table 1. 空间生态位宽度聚类分析(见图3)显示,春季主要甲壳类可分为3组,第1组由长手隆背蟹、日本蟳、口虾蛄3种组成;第2组由日本英雄蟹、葛氏长臂虾、戴氏赤虾、双斑蟳、须赤虾和中华管鞭虾6种组成;第3组由假长缝拟对虾和绵蟹组成;秋季主要甲壳类可分为2组(见图4),第1组由日本蟳、中华管鞭虾、口虾蛄、细巧仿对虾、三疣梭子蟹、刀额仿对虾、哈氏仿对虾、双斑蟳8种组成;第2组由戴氏赤虾、鹰爪虾、葛氏长臂虾、日本鼓虾、须赤虾、鲜明鼓虾、东海红虾7种组成。 图3 春季主要甲壳类物种的生态位宽度聚类分析Fig.3 Cluster analysis of niche breadth of major crustacean species in spring 图4 秋季主要甲壳类物种的生态位宽度聚类分析Fig.4 Cluster analysis of niche breadth of major crustacean species in autumn 春季主要甲壳类生态位重叠值和种间竞争系数如表2所示。由表2可知,春季主要甲壳类种对有55对,生态位重叠值为0~0.99。其中,生态位显著重叠(Qik>0.6)的种对有13对,占总种对数的23.64%,绵蟹和日本英雄蟹的生态位重叠值最大(0.99);生态位重叠有意义(Qik>0.3)的种对有15对,占总种对数的27.27%;其余27对的Qik<0.3,占总种对数的49.09%。而假长缝拟对虾与中华管鞭虾和绵蟹的生态位重叠值为0。 春季主要甲壳类种间竞争系数为0~0.69,平均值为0.26,种间竞争程度较低。其中,长手隆背蟹与日本蟳种间竞争系数最大(0.69),日本英雄蟹与双斑蟳、戴氏赤虾的种间竞争系数均较大(>0.60),而春季优势种,如须赤虾与戴氏赤虾、双斑蟳与口虾蛄之间的种间竞争系数较小。 表2 春季主要甲壳类生态位重叠值(主对角线以下)和种间竞争系数(主对角线以上)Table 2 The niche overlap(under the main diagonal)and interspecific competition coefficient(above the main diagonal)of major crustacean species in spring 秋季主要甲壳类生态位重叠值和种间竞争系数如表3所示,由表3可知,秋季主要甲壳类种对有105对,生态位重叠值为0~0.83。其中,生态位显著重叠(Qik>0.6)的种对有21对,占总种对数的20.00%,须赤虾和东海红虾的生态位重叠值最大(0.83);生态位重叠有意义(Qik>0.3)的种对有43对,占总种对数的40.95%;其余41个种对的Qik<0.3,占总种对数的39.05%。秋季共有60.95%的种对生态位重叠值超过0.3,表明秋季主要甲壳类物种空间生态位重叠达到有意义的种对比例较高。而须赤虾与鲜明鼓虾和葛氏长臂虾、东海红虾与鲜明鼓虾和葛氏长臂虾的生态位重叠值均为0。 秋季主要甲壳类种间竞争系数为0~0.97,平均值为0.44,种间竞争程度较春季高。其中,须赤虾与东海红虾的种间竞争系数最大(0.97),须赤虾与哈氏仿对虾的种间竞争系数也较大(0.91),其余种对如细巧仿对虾与口虾蛄的种间竞争系数较大(>0.85),而须赤虾与日本鼓虾、葛氏长臂虾与东海红虾的种间竞争系数为0。 主要甲壳类与环境因子的RDA结果如图5所示,由图5可知,春季的第1排序轴(RDA 1)特征值为0.388 4,第2排序轴(RDA 2)特征值为0.088 4,RDA 1和RDA 2与环境因子的相关系数分别为0.886和0.908。蒙特卡洛显著性检验结果显示,2个排序轴呈极显著差异(F=2.7,P=0.006)。2个排序轴共解释了主要甲壳类组成47.68%的变异。水深及表层盐度聚集在RDA 1的正轴,与RDA 1呈显著正相关;表层温度、底层温度和底层盐度聚集在RDA 2的负轴,与RDA 2呈显著负相关。其中,长手隆背蟹(S07)和中华管鞭虾(S04)聚集在RDA 2的负轴,与表层温度和底层温度呈显著正相关,说明温度是影响其分布的主要因素;须赤虾(S01)和日本英雄蟹(S08)聚集在RDA 1的正轴,与水深呈明显正相关,说明水深是影响其分布的最主要因素;日本蟳(S09)分布于2个排序轴的原点附近,表明该物种对环境因子无特别偏好,属广布性种类。 秋季的RDA 1特征值为0.339 8,RDA 2特征值为0.089 8,RDA1和RDA2与环境因子的相关系数分别为0.868和0.790。蒙特卡洛显著性检验结果显示,2个排序轴呈显著差异(F=2.0,P=0.024)。2个排序轴共解释了主要甲壳类组成42.96%的变异。底层温度聚集在RDA 1的正轴,与RDA 1呈显著正相关,表层盐度、底层盐度及表层温度聚集在RDA1的负轴,与RDA1呈显著负相关,其中,刀额仿对虾(A05)和口虾蛄(A15)聚集在RDA 1的正轴,与底层温度呈显著正相关,与表层温度呈显著负相关,说明温度对其分布影响较大;须赤虾(A01)和戴氏赤虾(A02)聚集在RDA2的负轴,与表层盐度呈显著正相关,说明表层盐度对其分布影响较大;东海红虾(A11)与底层盐度呈显著正相关,说明底层盐度对其分布影响较大。 表3 秋季主要甲壳类生态位重叠值(主对角线以下)和种间竞争系数(主对角线以上)Table 3 The niche overlap(under the main diagonal)and interspecific competition coefficient(above the main diagonal)of major crustacean species in autumn 图5 主要甲壳类与环境因子的RDA结果Fig.5 Redundancy analysis of major crustacean species and environmental factorsST—表层温度;SS—表层盐度;BT—底层温度;BS—底层盐度;D—水深;图中编码对应的物种参照表1。ST-surface temperature;SS-surface salinity;BT-bottom temperature;BS-bottom salinity;D-water depth;Codes in the figure are in accord with table 1. 空间生态位宽度反映的是物种利用资源的能力,表征种群在生境中的分布范围和生物量[27],空间生态位宽度越大,物种在空间维度上的分布范围越广、均匀度越高[10]。本研究结果显示,广温广盐种群的生态位宽度明显大于高温高盐和广温低盐种群,揭示了种群间环境适应及资源利用能力的差异性,如口虾蛄和日本蟳春、秋两季的空间生态位宽度均较大,为广生态位种,说明这2个物种在研究海域春、秋两季分布范围较广,且生物量在各调查站位较均匀,有较强的资源获取能力。广生态位种如中华管鞭虾、细巧仿对虾、三疣梭子蟹等,在温盐空间梯度变化较大的秋季,广生态位种的分布更为均匀,生态位宽度大,其物种群属典型广温广盐生态类种群[28],物种分布与该海域的温度、盐度等生态环境相适应,生境适应力强;而春季须赤虾、假长缝拟对虾等物种,以及秋季日本鼓虾、鲜明鼓虾等物种的生态位宽度均较低,对环境变化敏感,因此导致其物种分布不均,如须赤虾在春季集中分布于Z08、Z09、Z10站位,在秋季集中分布于Z01、Z08、Z09、Z10站位,物种集中度高,斑块状特征明显,该结果与文献[29]中的物种生态位宽度与其生境适应性密切相关这一结论相符。 主要甲壳类生态位宽度在春、秋季差异较大,总体上秋季物种的生态位宽度大于春季。秋季广生态位种占物种总数的53.33%,中生态位种占46.67%,因此,甲壳类群落中生态位宽度较高的种群占主导地位,这一方面由于甲壳类物种的消长与研究海域秋季水温、盐度等主要环境因子较高有关[30],另一方面由于4月水温升高,利于多种甲壳类物种繁殖,物种集群明显,分布呈斑块性增强,导致甲壳类物种受繁殖习性影响,分布不均,生态位宽度较小。伏季休渔结束后,研究海域甲壳类资源得到一定的养护,数量增多,分布范围扩大,导致物种Qik普遍较高,该结论也与徐开达等[31]的研究结果相符。 比较分析主要甲壳类的生态位宽度与IRI可知,受物种数量及分布的影响,二者并无直接关系,如春季日本蟳的生态位宽度(2.19)高于双斑蟳(1.18),而其IRI(474.49)却低于后者(2 593.67);秋季哈氏仿对虾的IRI(3 859.06)远高于刀额仿对虾(353.60),而后者的生态位宽度(2.31)却高于前者(2.25),这说明生态位宽度与IRI的计算结果因生态特征角度不同而存在差异。在用生态位宽度对物种进行测度时,因综合考虑了物种尾数、调查站位及其分布差异,较IRI更全面。综上可知,IRI较大的物种,其生态位宽度并不大,这与卢占晖等[32]的研究结果相符。 生态位重叠反映的是同一生境内物种资源需求的相似性[33],生态位重叠显著,则种对间的资源需求趋同[34],如春季日本英雄蟹与绵蟹的生态位重叠值最高,可能与二者在调查海域空间资源序列上的同域性最强,且二者均属广温广盐类群,在水深20~100 m的泥底或泥沙底海域均有分布[35],即生态习性相似有关。秋季生态位重叠值最高的种对为须赤虾-东海红虾,这可能与二者生态习性相似有关,如同属高温高盐生态类群,均栖息于高盐海域,均分布于混合水区近高盐水一侧[36],且在舟山渔场内群体数量较大[37],当资源不足时种间易产生较激烈的竞争。哈氏仿对虾与中华管鞭虾因在洄游及繁殖季节方面具有相似性[36],二者间的生态位重叠值显著。日本鼓虾与鲜明鼓虾虽同为鼓虾属,生态习性相似,春、夏季均生活在泥沙底质的浅海,多穴居于泥沙,在分布范围上重叠程度较高[38],但本研究中二者的生态位重叠值在所有种对中并非最高,与刘明华等[15]的研究结果存在差异,推测可能是由于研究海域范围较广,虾类饵料生物丰富,这2个物种占据了相近的生态空间,并未导致竞争或仅存在微弱竞争[39],从而实现了共存。 研究结果表明,甲壳类物种生态位重叠与物种间在调查站位中的重叠率有关。春季戴氏赤虾-双斑蟳种对的生态位重叠值为0.98,重叠程度高,且从调查站位分布情况看,二者的站位重叠率高达81.25%。秋季须赤虾-鲜明鼓虾种对的生态位重叠值为0,须赤虾分布的海区具有高温高盐的特点,生长分布更容易受温度和盐度的影响,而鲜明鼓虾适温范围较广,且生活于泥沙底质的浅海,除种对间生态类群不同导致的物种本身生态学特性不同外,二者的站位重叠率仅为18.75%,该结论与WATHNE等[34]提出的生态位重叠显著的种对间与站位重叠程度较高的研究结论相符。 本研究中甲壳类物种生态位重叠与生态位宽度并非直接相关,如春季戴氏赤虾仅为中生态位种,但其与须赤虾和双斑蟳的生态位重叠值均较高(≥0.97),而秋季生态位宽度最大的2个物种中华管鞭虾与口虾蛄之间的生态位重叠值为0.56,生态位重叠并不显著,说明环境资源的高异质性、物种的斑块状分布、种群生态学特性差异等因素,均会导致不同生态位宽度的物种间产生不同程度的生态位重叠[5],因此,不能仅依靠生态位宽度判断某物种与生境内其他物种的竞争性。 RDA能很好地解释物种与环境之间的内在联系,反映甲壳类对不同生态栖息环境的适应性特征。结果发现,温度和栖息海域盐度的变化影响甲壳类物种的生物量及空间分布,陆架和岛礁生态区的差异影响甲壳类物种分布,如春季属高温高盐生态类群的长手隆背蟹,其生长分布更易受温度和盐度的影响,由于其属于底栖动物种类,因此RDA结果中表层温度和底层温度对物种分布的影响是合理的,该结论与卢衎尔等[40]的研究结果相符;而葛氏长臂虾在盐度为25~34、水温为8~25℃的沿岸水域和高低盐水交汇的混合水域均有分布,属广温广盐类群,生境适应范围广,温度和盐度变化对物种分布影响较小,因此RDA结果与甲壳类种群生态习性相符。RDA结果也能间接反映不同采样站点间的环境差异,如春季须赤虾和日本英雄蟹受水深影响较大,二者均集中分布于Z08、Z09、Z10站位,均处于123.8°E以东,水深明显增加,对物种分布产生影响。 RDA可对物种生态位宽度与重叠进行补充说明,借助生态位分化解释主要甲壳类在生态位重叠值中存在差异的原因,如春季须赤虾(S01)和日本英雄蟹(S08)资源需求相似性较低,但二者均受水深影响,具有相似的底栖生境,且分布于相近区域,因而导致二者虽均为窄生态位种但重叠值高。秋季刀额仿对虾(A 05)与口虾蛄(A 15)的生态位宽度较大,但生态位重叠不显著,RDA结果显示口虾蛄(A 15)受底层温度影响较大,刀额仿对虾(A 05)受相关环境因子影响较小,因此种对间的生态位分化呈空间分布差异。 本研究将生态位重叠与种间竞争系数的计算结果相结合进行综合分析,增加了对主要甲壳类种间竞争、共存及生态位分化情况的判断依据,将分析结果量化。如秋季须赤虾-东海红虾种对的空间生态位重叠最大,其种间竞争系数也最大,说明二者之间可能存在明显的种间竞争;春季戴氏赤虾-绵蟹种对的空间生态位重叠值较高,但其种间竞争系数较低,空间生态位分化可能由季节变化、物种摄食习性变化导致,此时二者的生态位重叠可能由生态空间相近导致。 本研究分析了舟山群岛东侧海域主要甲壳类物种的空间生态位宽度和生态位重叠值,结合RDA与种间竞争系数探讨了主要甲壳类物种的生态位分化情况。为进一步更准确地揭示物种的生存竞争状况,将来需增加时间和空间维度上的调查航次及调查站位,增加生态位计测维度,特别是营养和时间生态位等方面的研究,以期更加全面地了解该海域甲壳类物种的资源状况。
2.3 生态位重叠和种间竞争系数
2.4 生态位分化及物种与环境因子的关系


3 讨 论
3.1 生态位宽度分析
3.2 生态位重叠分析
3.3 生态位分化及物种与环境因子的关系分析
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
中国农业科学(2022年10期)2022-06-28
广东蚕业(2021年12期)2022-01-18
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
人大建设(2019年5期)2019-10-08
热带农业科学(2019年8期)2019-09-19
人大建设(2019年3期)2019-07-13
人生十六七(2015年5期)2015-02-28
销售与市场·管理版(2009年21期)2009-09-03
