
中国沿海三井卷曲纽虫的分布与COI序列分析*
2021-07-27 14:29高凯梦孙世春
中国海洋大学学报(自然科学版) 2021年9期
高凯梦, 孙世春
(中国海洋大学水产学院, 海洋生物多样性与进化研究所, 山东 青岛 266003)
卷曲属(EmplectonemaStimpson, 1857)属于纽形动物门(Nemertea) 针纽纲(Hoplonemertea)单针目(Monostilifera)卷曲科(Emplectonemertidae Bürger, 1904),是一类身体纤细的单针纽虫,目前包含二十多个物种。在中国沿海曾报道细卷曲纽虫(Emplectonemagracile(Johnston, 1837))和三井卷曲纽虫(EmplectonemamitsuiiYamaoka, 1947) 两种[1-2]。其中,三井卷曲纽虫在中国(大连)的记录[2]存有较大疑问(见下文)。
三井卷曲纽虫最早由山冈贞一(Yamaoka)发现于日本下田,原始描述是由奥田四郎对山冈贞一的遗作简化后发表于《日本动物图鉴》(改订增补)[3],山冈氏对该种的完整描述则于2005年发表[4]。该种的已知分布区包括日本东海岸、日本海沿岸[4-7]和韩国[8]。
近年来,本课题组在中国沿海纽形动物采集调查时,在中国的东海、南海8地采集到三井卷曲纽虫标本27个,并对其开展了形态学和分子生物学研究。
1 材料与方法
1.1 标本的采集、观察与处理
调查采集主要在岩相潮间带进行。采集方法一是肉眼搜索,二是采集海藻、固着贝类、藤壶等污损生物,加海水浸泡通过溶氧胁迫获得纽虫。所获纽虫饥饿24 h后,直接以95%乙醇固定用于后续分子生物学研究,或活体运回实验室进行形态观察。
活体纽虫用7.5% MgCl2麻醉,先用体视显微镜(尼康SMZ800)观察、测量、拍照。然后将标本转移到载玻片上,用盖玻片轻压,于显微镜(尼康E600)下进一步观察吻针等内部构造。标本以波恩氏液或10%海水福尔马林固定,或以10%海水福尔马林固定头部,以95%乙醇固定尾部(用于分子生物学研究)。形态学标本固定至少24 h后转移到70%乙醇保存,乙醇固定的分子生物学样品于-25 ℃保存。
本研究在中国沿海8个地点获得的27个标本,其中用于分子生物学研究的标本21个,采集信息见表1、图1。标本保存于中国海洋大学海洋生物多样性与进化研究所。除表1所列地点外,近20年来,我们还在辽宁大连、旅顺、大长山岛、獐子岛、广鹿岛、葫芦岛,河北秦皇岛,山东北隍城岛、大钦岛、砣矶岛、北长山岛、南长山岛、大黑山岛、崆峒岛、威海、荣成马山、乳山、鳌山卫、青岛、灵山岛、日照,福建平潭、南日岛、惠安、厦门、东山,广东南澳岛、深圳东山、硇洲岛、海陵岛,广西北海、防城港、涠洲岛,海南海口、临高、莺歌海、儋州、三亚、陵水、永兴岛、七连屿等40余个地点进行了样品采集(调查地点覆盖了自渤海至南海中国各海区),但未采集到本种纽虫。
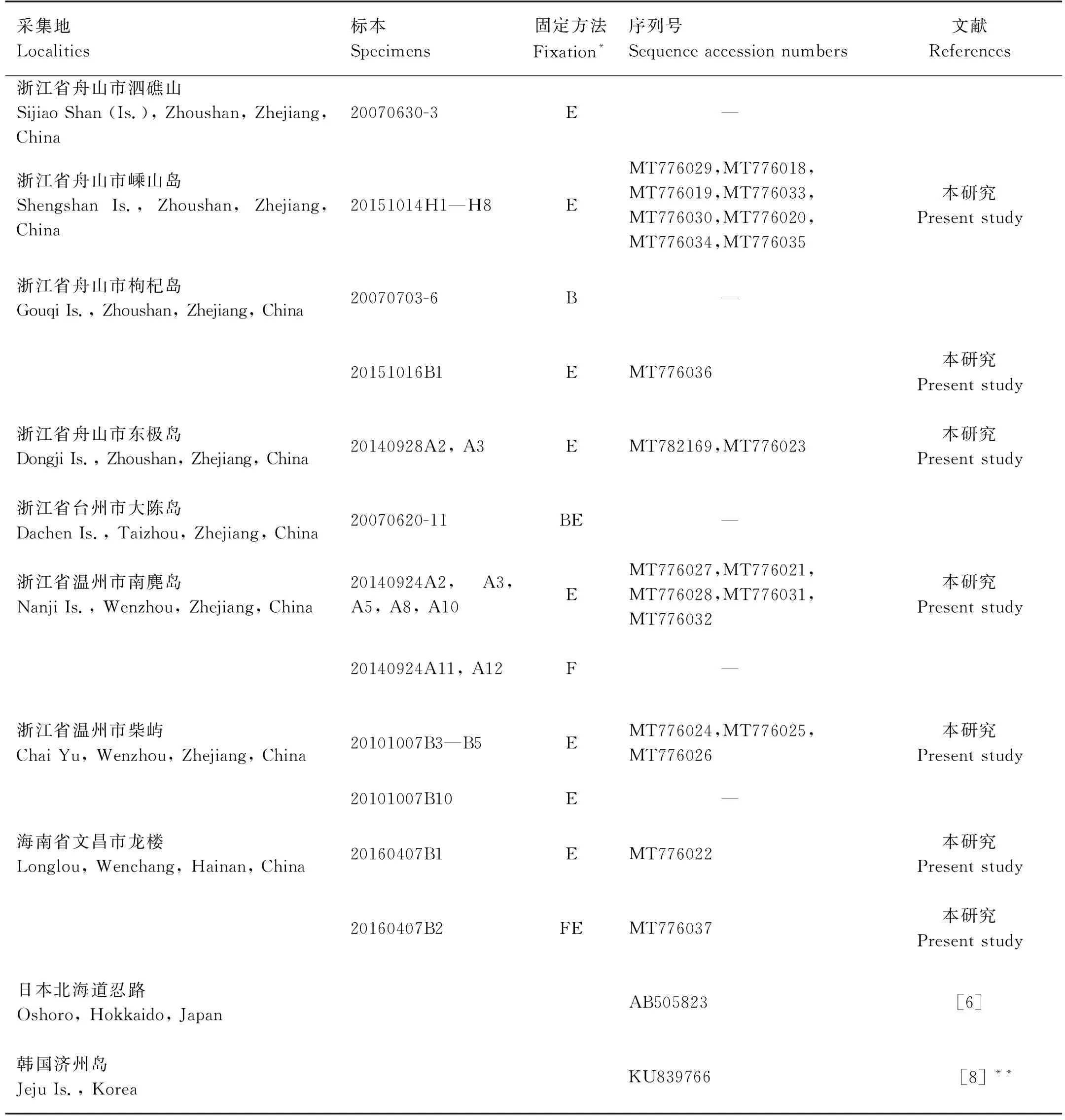
表1 研究标本的采集信息及COI的GenBank序列号Table 1 Sampling information of specimens used in the present study, and the GenBank accession numbers of COI sequences
1.2 分子生物学研究
所有样本均使用试剂盒MicroElute Genomic DNA Kit200(OMEGA, USA)进行DNA的提取。利用通用引物正向引物LCO-1490 GGTCAACAAATCATAAAGATATTGG和反向引物HCO-2198 TTAACTTCAGGGTGACCAAAAAATCA进行COI基因片段扩增[9]。PCR反应体系为25 μL,其热循环条件为:94 ℃下预变性2 min,循环(94 ℃下变性1 min,49 ℃退火40 s,72 ℃延伸1 min)35次,最后在72 ℃延伸7 min。扩增产物用1%琼脂糖凝胶进行电泳检测,经纯化后用相同引物进行双向测序(北京六合华大基因科技有限公司)。DNA序列的GenBank号(包括从GenBank下载的2条序列)见表1。
用MEGA ver.6.0 软件[10]对序列进行比对、计算遗传距离(未校正p-距离,uncorrected p-distance;以下简称p-距离)。用软件TCS v.1.21[11]在95%的链接概率下生成单倍型网络图。单倍型及核苷酸多样性参数由DnaSP ver.5.10.01软件计算[12]。
2 结果
三井卷曲纽虫Emplectonema mitsuii Yamaoka, 1947
EmplectonemamitsuiiYamaoka: 山冈贞一 1947: 1466, fig. 4133[3]; 佐藤隼夫, 伊藤猛夫 1961: 187, fig. 7.1.7[13]; 奥田四郎, 岩田文男 1965: 397, figs a-d[14]; 宋鹏东等 1989: 53-54[2]; Crandall et al. 2001: 177-178, pl. 1 figs 1-4, pl. 2 figs 16, 16a, 16b[5]; Yamaoka 2005: 142[4]; Kajihara et al. 2011: tab. 1[6]; Sundberg et al. 2016: tab. S1[8]。
2.1 标本及采集信息
研究标本共27个。采自浙江省舟山市泗礁山、嵊山岛、枸杞岛、东极岛,台州市大陈岛,温州市南麂岛和柴屿,海南省文昌市龙楼(铜鼓岭)共8个地点,详细信息见表1、图1。所有标本均采自潮间带,除在柴屿发现个别个体生活于岩石缝隙外,均生活于海藻丛中。

(圆点表示本研究的采集地点;三角为其它文献记录[2]。Circles show collecting sites of present study; the triangle shows the site of a previous record [2].)
2.2 形态
体扁平,前部背面稍隆起,腹面平,麻醉后常向腹面弯曲。麻醉后体长约15~29 mm,体宽0.5~0.7 mm。体黄,显绿,或黄绿色(见图2A~E),有的个体背面具青色小斑点(见图2C)。头端尖,运动时尤为明显。眼点较大,分4组,前两组各2~6个,后两组各1~4个,一般橙红色,偶见暗褐色(见图2C, D)。前头沟在腹面,向上不延伸至背面;后头沟位于后眼之后,呈环绕身体的闭环(见图2C~E)。头部中央具一橙红色色斑,位于左右前眼点之间(见图2B, C)。吻孔腹位,纵裂缝状(见图2D)。吻腔不及体长的一半。吻主针光滑,长62~85 μm,针座近锥形,后端圆,长59~69 μm,针座宽20~30 μm(见图2I)。主针略长于针座,针长/座长为1.02~1.14。副针囊2个,各具副针2~3枚。肠具侧盲囊。没有对所有个体进行镜检,但龙楼4月标本、枸杞7月标本、柴屿10月标本见成熟个体。生殖腺位于体后部约60%,数目很多(见图2F, G)。精子头部棒状,约3~4 μm,具很长的鞭毛(见图2H)。

(A:活体外形(柴屿标本);B, C:头部背面观,示眼的数目和颜色变化(B大陈岛标本,C柴屿标本);D:头部腹面观(枸杞岛标本);E:头部侧面观(枸杞标本);F:柴屿10月份雌性个体的肠区压片,示成熟卵;G:柴屿10月份雄性个体的肠区压片,示成熟精巢;H:精子;I:主针及针座。gr1: 前头沟;gr2: 后头沟; rs: 吻口。比例尺:B~E = 200 μm; F,G = 100 μm; H = 10 μm; I = 50 μm。A: A complete worm from Chai Yu (Island); B, C: Dorsal view of head (A from Dachen Island, B from Chai Yu), showing number and color variation of eyes; D: Ventral view of head (from Gouqi Island); E: Lateral view of head (from Gouqi Island); F: Squeezed intestinal region of a female collected from Chai Yu in October; G: Squeezed intestinal region of a male collected from Chai Yu in October; H: Sperms; I: Central stylet and basis. gr1: Anterior cephalic groove; gr2: Posterior cephalic groove; rs: Rhynchostome. Scale bars: B~E = 200 μm; F,G = 100 μm; H = 10 μm; I = 50 μm.)
2.3 DNA序列
将中国21个标本的COI序列(658 bp)与GenBank下载的2条序列(见表1)比对后,得到长度为513 bp的序列数据集。对23条COI序列的分析共检测到4个不同单倍型(Hap1,Hap2,Hap3,Hap4),构成一个单倍型网络。其中,主单倍型Hap1包含19个个体,来自中国沿海的6个地点(嵊山岛、枸杞岛、东极岛、南麂岛、柴屿、龙楼);Hap2包含2个个体,分别来自日本忍路和韩国济州岛;Hap3和Hap4各包含1个个体,分别采自嵊山岛和柴屿(见图3)。所有23条序列的p-距离为0.000 0~0.017 5(平均0.003 3),平均单倍型多样性(Hd)为0.320±0.121, 平均核苷酸多样性(π)为0.003±0.002,呈现单倍型多样性与核苷酸多样性均低的模式。中国沿海21个样本间的p-距离为0.000 0~0.003 9(平均0.000 4),平均单倍型多样性Hd为0.186±0.110, 平均核苷酸多样性π为0.000 4±0.000 2。东海(浙江5地19个标本)和南海(龙楼2个标本)样本间平均遗传距离仅为0.000 2。中国与日本(忍路)/韩国(济州岛)样本间遗传距离明显较大,均为0.017 7(日本与韩国两样本的COI序列相同)(见表2)。
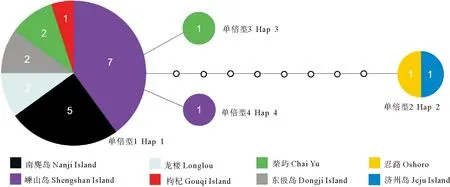
(图中数字为标本数;单倍型间的每个空心圆代表一个缺失的单倍型。 Numbers correspond to the number of specimens; each blank circle indicates a missing haplotype. )

表2 不同地点基于COI序列的遗传距离(未校正p-距离)Table 2 Averaged genetic distance (uncorrected p-distance) for COI sequences between different locations
3 讨论
本文观察的中国标本的形态与原始描述基本一致。与模式标本相比[4-5],本文标本存在以下细微差异:体型较小(体长15~29 mmvs.50~70 mm);体色黄绿色(见图2A~E),而模式标本体色白黄;多数标本眼的颜色与原始描述一致,呈朱红色,但也发现眼呈暗褐色的标本(见图2C);吻针和针座较小(主针长62~85 μmvs.96~130 μm;针座长59~69 μmvs.80~110 μm),该差异可能是因为本文标本个体较小。Sundberg等对大量样品的分析结果显示,纽形动物COI基因的种内变异上限约为3%[8],本文样品与韩国、日本标本间序列差异小于此值(见表2)。基于COI序列的TCS物种界定分析显示,中国、日本、韩国的23个标本归于同一单倍型网络,也表明他们隶属同一物种[15]。
除本文记录的分布点外,本种已知分布区包括日本下田(静冈)[4-5]、三崎(神奈川)、忍路(北海道)和佐渡岛(新潟)[6-7],韩国济州岛( 见表1)。宋鹏东等曾在大连记录本种[2],但近年来,作者及同事在辽宁至山东的黄渤海沿岸进行了多次调查采集,超过20个调查地点具有本种的典型生境(岩相潮间带,有大型藻类生长),但均未发现本种。三井卷曲纽虫是一暖水种,其在纬度较高的北海道忍路出现可能与对马暖流有关(柁原宏,个人通讯)。因此,本种在大连的分布可能系一误记。
COI序列的分析结果表明,三井卷曲纽虫的单倍型多样性和核苷酸多样性总体较低,特别是浙江、海南90%个体共享同一单倍型,其他个体与该主单倍型间也仅有一个碱基差异,提示其可能经历过瓶颈效应或种群是由少数个体通过奠基者效应而建立[16]。虽然济州岛与忍路间、浙江各采样点与海南龙楼间的地理距离明显大于浙江各采样点与济州岛间的距离,但浙江和济州岛的个体间遗传距离远大于济州岛与忍路、浙江与海南个体间的遗传距离(见表2)。遗憾的是,济州岛和忍路的样品仅有1个,海南样品仅有2个,包括模式产地在内的日本东海岸多个分布地均无可用于DNA研究的样品,现有数据尚不能探讨三井卷曲纽虫的地理分布格局的形成历史。
4 结语
形态研究和DNA序列分析表明,浙江和海南的标本与在日本、韩国报道的三井卷曲纽虫属于同种。在中国沿海三井卷曲纽虫分布于浙江、海南等暖水水域,前人在辽宁大连的记录可能为一误记。中国沿海三井卷曲纽虫的遗传距离、单倍型与核苷酸多样性均很低,但与韩国、日本2个样品间序列差异较大,提示有必要通过深度采样在种群水平上解析其地理分布格局的形成历史。
致谢:曾晓起教授、郑晓东教授等在样品采集过程中给予大量帮助,俄罗斯科学院远东分院Alexei V. Chernyshev教授提供GenBank序列样品的准确采集地信息,北海道大学柁原宏教授对研究物种在日本的分布给予进一步解释,谨此致谢!
猜你喜欢
中国化肥信息(2022年8期)2022-12-05
中国船检(2021年7期)2021-11-23
现代妇女(2019年12期)2019-12-19
小星星·阅读100分(高年级)(2018年3期)2018-03-26
环球时报(2017-10-31)2017-10-31
环球时报(2017-08-30)2017-08-30
诗林(2016年5期)2016-10-25
旅游世界(2015年9期)2015-09-29
初中生学习·中(2005年3期)2005-01-10
