
乳腺癌细胞系MCF-7中阿霉素的代谢物分析
2021-09-08 07:19陈雨蕉惠人杰朱景宇
中国药理学通报 2021年9期
陈雨蕉,惠人杰,王 叙,朱景宇,贾 磊,金 坚,陈 蕴
(江南大学药学院药物设计与分子药理研究室,江苏 无锡 214122)
阿霉素(doxorubicin,DOX)是一种传统的蒽环类化疗药物,乳腺癌是其适应症之一。阿霉素的作用机制是通过细胞膜并嵌入DNA,导致DNA断裂和干扰DNA复制[1],从而发挥其抗肿瘤作用。体内药代动力学表明,约50%的阿霉素以原型药形式排出人体,蒽环类药物的体内代谢是通过羟基化、形成半醌或脱氧苷元进行的,从而增强或抑制蒽环类药物的抗癌活性[2]。已报道代谢产物有阿霉素醇、阿霉素半醌、阿霉素羟基苷元、阿霉素脱氧苷元和阿霉素醇苷元[3]。其中,7-脱氧阿霉素酮和阿霉素醇已被证明与心肌细胞的心脏毒性有关[1]。
经典的药代动力学可以获得机体对药物处置的基本信息,包括药物在机体内的吸收、分布、代谢和排泄的数据。然而越来越多数据表明,传统的体内药代动力学不能完全解释药物对肿瘤的药理作用,一些药物在靶细胞中无法积蓄导致药物治疗效果差[4]。药物的细胞内处置过程和结合靶点的浓度是观察靶器官治疗反应的决定因素[5]。基于血浆药物浓度的经典药代动力学研究可能不足以预测体内反应,无法从肿瘤靶细胞角度解释药物分子作用的行为特征,现有研究缺乏靶细胞对于药物处置的认识,需要扩大药代动力学范围,对其进行进一步研究[6]。
人乳腺癌细胞系MCF-7是阿霉素适应症乳腺癌公认的成熟的细胞模型,在MCF-7的耐药研究中发现其耐阿霉素耐药株MCF-7/DOX相比野生细胞MCF-7/WT能能耐受更高浓度药物,且代谢阿霉素速率更高[7],有特殊的胞内药物代谢物,故选择其作为研究阿霉素靶细胞内处置的靶细胞模型。开发了一套肿瘤靶细胞处置药物的分析方法,MCF-7/DOX阿霉素耐药株作为靶细胞模型,使用超高效液相色谱-四级杆串联飞行时间质谱联用仪(UPLC-Q-TOF-MS/MS)分析研究阿霉素胞内药物代谢物,通过高分辨率多级质谱解析推断未知代谢物结构。
1 材料与方法
1.1 实验材料
1.1.1细胞 野生型人乳腺癌细胞系MCF-7/WT,购自美国菌种保藏中心ATCC细胞库;阿霉素耐药细胞MCF7/DOX为本实验室由MCF-7/WT构建培养[8]。
1.1.2试剂 盐酸阿霉素(纯度>98%,货号:MB1087)购自大连美仑生物科技有限公司;色谱级甲醇和甲酸购自美国TEDIA公司;实验用水为超纯水;其余溶剂均为分析纯;CellTiter-Blue细胞活力试剂盒购自无锡莱弗斯生物公司。
1.1.3仪器 ACQUITY UPLC超高效液相色谱仪(沃特世,美国);Xevo G2 Q-TOF四级杆串联飞行时间质谱仪(沃特世,美国)。
1.2 实验方法
1.2.1细胞培养 配置含阿霉素培养基培养MCF-7/WT,使用CellTiter-Blue试剂盒测定细胞对阿霉素的IC50,从IC50的1/10开始,低浓度诱导其耐药,待细胞适应且稳定生长后逐步增加药量。直到多药耐药细胞系MCF-7/DOX耐药倍数稳定在100倍以上,耐药倍数=IC50(MCF-7/DOX)/IC50(MCF-7/WT)。两种细胞系都使用添加了10%血清、1%青霉素-链霉素抗生素和2 kU·L-1胰岛素的DMEM培养基培养,耐药细胞MCF-7/DOX额外添加2.0 mg·L-1的DOX以维持其耐药性,并在实验前撤药7 d以防止药物对实验结果的干扰,生长环境为37 ℃及5% CO2。
1.2.2细胞存活率检测 用Cell Titer-Blue试剂测定细胞给阿霉素后24 h存活率,按照说明书操作,用酶标仪测量荧光值。未经DOX处理的细胞作为对照,每个浓度设置3个复孔,于EX 560 nm, EM 590 nm 进行荧光检测,使用软件GraphPad Prism7计算细胞存活率。
1.2.3液相色谱-串联质谱样品的制备 MCF-7/WT野生型细胞和MCF-7/DOX耐药细胞分别在T75细胞培养瓶中培养至汇合度80%,后更换含阿霉素培养基培养24 h,阿霉素浓度分别为7.7 mmol·L-1和370 mmol·L-1,在此条件下两种细胞存活率都在70%以上。将细胞用胰酶消化收集后,用PBS缓冲液洗涤离心3次后除尽PBS。使用乙醇:0.3 mol·L-1盐酸 ∶100%三氯乙酸=2 ∶2 ∶1的提取液提取阿霉素靶细胞代谢物,每3.8×106个细胞用1 mL提取液重悬并用400 W冷水浴超声1 h破碎细胞壁提高提取效率,后在12 000g,4 ℃下离心30 min后收集上清液过0.22 μm有机滤膜,存放于-80 ℃以待进一步的UPLC-MS/MS分析。
1.2.4UPLC-MS/MS条件
1.2.4.1 UPLC色谱条件 色谱柱使用ACQUITY UPLC BEH C18 Column(1.7 μm,2.1 mm×50 mm),柱温25 ℃。流动相使用甲醇(A相)和0.1%(V/V)甲酸水(B相)。使用梯度洗脱法,从10% A开始,随后在9 min内线性增加到95% A,保持到11 min后在11.5 min降回10% A,保持到15 min进行柱平衡,总时间为15 min,流速为0.2 mL·min-1,进样量为3 μL,二极管阵列检测器(PDA)检测波长范围设置为200-800 nm。
1.2.4.2 Q-TOF-MS/MS质谱条件 电喷雾离子源(ESI),正离子模式扫描:扫描范围m/z100-1 500,毛细管电压3.0 kV,锥孔电压40 V,锥孔气流速60.0 L·h-1,脱溶剂气(N2) 流速600 L·h-1。碰撞能量20 eV。
1.2.5分子对接分析 将阿霉素及其代谢物与DNA进行分子对接,以进一步了解阿霉素及其代谢物与DNA之间的作用机制。DNA晶体结构(PDB ID:2DND)取自PDB数据库(http://www.rcsb.org/);阿霉素三维结构从Pubchem Compound数据库(https://www.ncbi.nlm.nih.gov/pccompound/term=)检索并下载;使用Schr dinger2017基于阿霉素结构绘制出m/z=606和m/z=641两个代谢物的结构。使用Protein preparation wizard模块对DNA结构进行预处理,然后使用LigPrep模块优化3个小分子,使用Receptor grid generation模块以DNA晶体结构中的配体为中心生成格点文件,最后应用Glide模块进行对接,对接采用Standard precision(SP)精度,其余参数默认。
2 结果
2.1 细胞存活率测定MCF-7/DOX和MCF-7/WT细胞阿霉素处理24 h存活率如Fig 1,其中X轴为阿霉素浓度的对数值、Y轴为细胞存活率,取细胞给阿霉素24 h后存活率约70%作为给药浓度,MCF-7/WT为7.7 μmol· L-1,MCF-7/DOX为370 μmol·L-1。

Fig 1 Determination of cell survival rate of MCF-7/WT andMCF-7/DOX treated with various concentrations of DOX for 24 h
2.2 UPLC-MS/MS分析MCF-7耐药型与野生型阿霉素代谢物的差异对MCF-7/WT和MCF-7/DOX细胞裂解物提取液进行UPLC-Q-TOF-MS/MS分析。羟基化的三元醌环是蒽环类化合物红色至橙色的特征发色团,包括阿霉素在内的蒽环类药物在可见光范围内的480 nm处有特征吸收[1],获得480 nm处色谱图,如Fig 2所示,阿霉素保留时间为5.64 min。虽然两种细胞代谢物中阿霉素色谱峰的吸收强度大致相等,但在耐药细胞色谱中观察到许多未在野生细胞色谱中发现的代谢物峰。其中,保留时间为6.17 min、6.58 min的两种物质分别对应m/z574和m/z588,其结构已在前期发表的文献中进行了报道[9],为阿霉素的结构基础上的氧化-烷基化修饰结果,修饰位点在10号位,分别加上氧甲基和氧乙基。本研究在此基础上又发现了两种新的代谢物(M1和M2),保留时间分别为6.02 min和7.87 min,m/z分别为606和641。其中M1与Wang等[9]研究中的代谢物可能存在结构联系,而M2可能与细胞中氨基酸代谢相关。
2.3 MCF-7/DOX中的阿霉素代谢物的质谱裂解分析Fig 3显示了阿霉素标准品和来自耐药细胞的阿霉素代谢物(M1和M2)的一至四级质谱裂解过程。一级图谱MS1显示阿霉素m/z544、阿霉素代谢物M1m/z606及M2m/z641 (Fig 3A、E、I),阿霉素的二、三、四级图谱(MS2、MS3、MS4)分别通过锁定m/z544、397、379获得碎片(Fig 3B、C、D),代谢物M1的二、三、四级图谱(MS2、MS3、MS4)分别通过锁定m/z606、441、395获得碎片(Fig 3F、G、H),代谢物M2的二、三、四级图谱分别通过锁定m/z641、494、476获得碎片(Fig 3J、K、L)。
通过多级质谱分析,推断阿霉素的质谱裂解途径为544→397→379→321。阿霉素MS2(Fig 3B)显示有m/z397和379两个强峰,对应于其D环上的糖苷基团的掉落(Δm/z=147),以及碎片离子进一步脱水所得的峰(Δm/z=165)。虽然代谢物M1、M2与阿霉素有不同的分子量,但是它们与阿霉素有相同的碎片丢失。分别锁定母离子m/z606、m/z641进行二级质谱碎裂得到MS2谱图(Fig 3F、J),显示两个代谢物结构碎片中有Δm/z=147或165的丢失,(M1:m/z606→441,Δm/z=165;M2:m/z641→494,Δm/z=147)。这个结果表明阿霉素与M1、M2结构中糖苷基团结构基本一致,这也是其作为阿霉素代谢物的重要证据。
代谢物M1(m/z606)的MS2显示m/z395和377两个强峰(Fig 3F),对应阿霉素MS2碎片中m/z397和379有一个不饱和度的差异(Fig 3B),MS3结果显示,395和377信号很强(Fig 3G),它们很可能与阿霉素的397和379这两个碎片有相关性。随后选择MS3中的碎片m/z395进行碎裂,MS4显示m/z377强峰,验证了其来源于碎片m/z395。多级质谱显示M1离子m/z395的碎裂方式与m/z397(DOX)类似,通过多次脱水都可以得到离子m/z321。
代谢物M2的MS2显示有碎片m/z641、494、476和458,与阿霉素碎片m/z544、397、379和361比对,都有Δm/z=97的差异,推测代谢物M2与阿霉素主体结构一致,并具备一个稳定的附加基团。m/z494的MS3显示476和458信号很强(Fig 3K),裂解行为与阿霉素中的379和361碎片离子相似。m/z476的MS4显示319碎片离子且信号强(Fig 3L)。代谢物M1与Wang等[9]推断的m/z=574和m/z=588的代谢物结构相当类似,在它们的多级质谱中都观察到相同的m/z395、377和321,并遵循相同的碎裂过程(606→441→395→377→321)。代谢物M2与阿霉素相差97分子量,并且在多级质谱的碎裂过程中始终保持该质荷比差异(M2/DOX:641/544→494/397→476/379,Δm/z=97),可以判断其为阿霉素相关代谢产物。
2.4 阿霉素的代谢物的结构推测根据阿霉素和代谢物质量碎裂模式的异同,推导了代谢物的化学结构及其质谱碎裂途径,如Fig 4所示。
从质量数差异推测代谢物M1为已报道代谢产物m/z588的羟基化产物,其代谢过程同已报道的m/z588过程相似,首先脱去糖苷基团和一分子水,随后m/z441的碎片离子产生二级离子m/z395,根据高分辨质谱的数据,推断其为CH4O(m/z=32.03)的丢失。对m/z377的次级碎片离子研究表明,其产生m/z349、321时连续丢失两次CO(m/z=27.99),发生了类似C=O基团的级联断裂。为了保证糖苷基团完整掉落,7、8位置不能被修饰,且氧化及一系列甲基化位置若发生在14位则化合物结构不稳定,故修饰位置只可能在10位。m/z606在m/z588基础上继续氧化,增加的氧化修饰位置极大可能在D环侧链的13位,C=O双键还原后加上羟基。
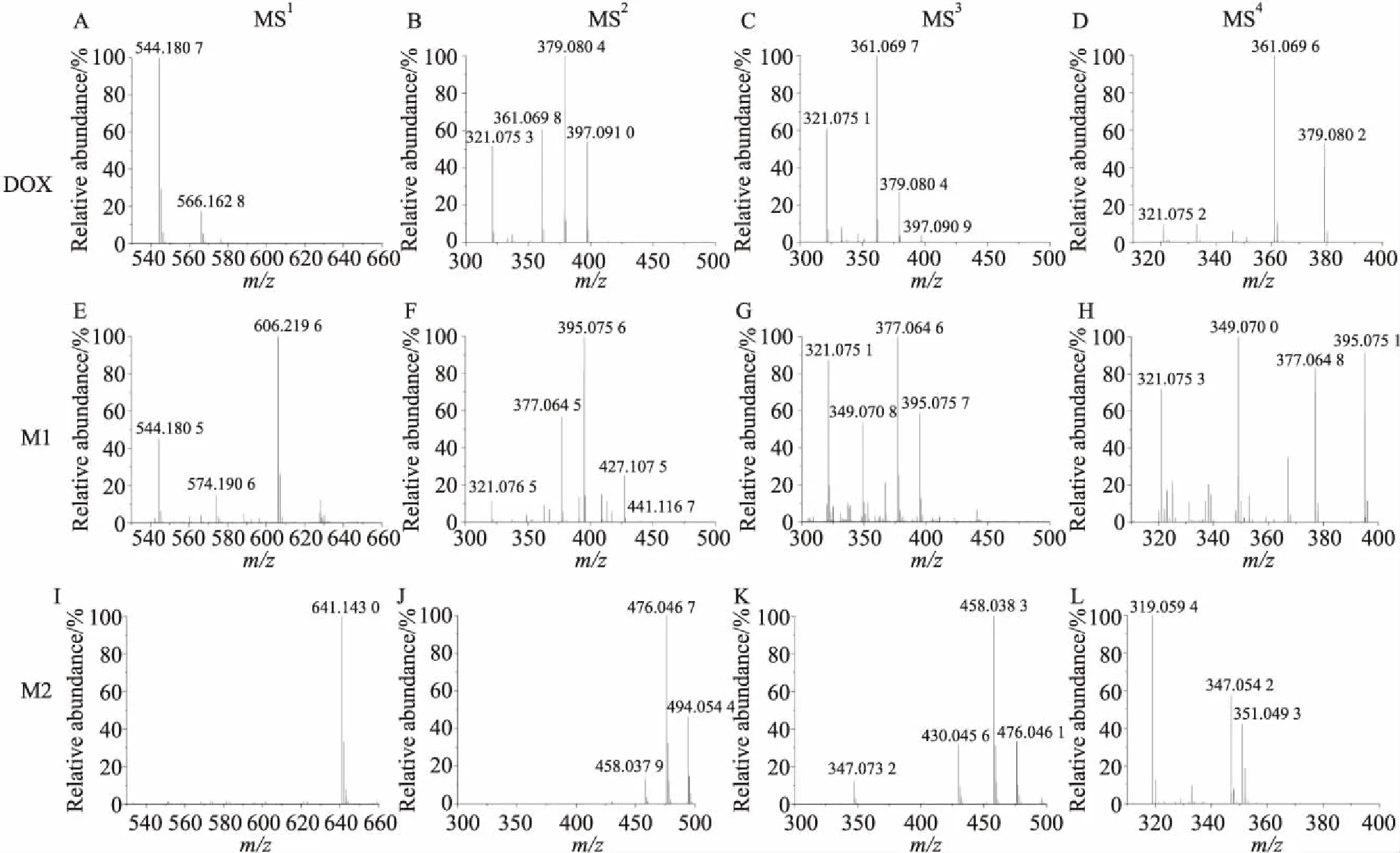
Fig 3 MSn of DOX and its metabolites M1 and M2
而代谢物M2与阿霉素质量数相差97,并在裂解过程中保持该质量差异,最终产生与蒽环基本结构m/z321具有一个不饱和度差异的碎片m/z319。该结果表明,在蒽环的某个活性位点新增了一个质量数为97的化学基团,且结合牢固,但由于未有更多裂解碎片信息,暂时还未获得更多碎片信息以准确判断该化学基团结构。
根据分子量推测,很可能为脯氨酸与阿霉素上羟基脱水缩合后的残基质量(m/z=97.06)。为了保证糖苷基团的完整掉落,修饰位置不可能在糖苷基团上,若在D环侧链末端羟基位置上,则不可能出现m/z458、430的碎片代谢物,因空间位阻效应也不能在6位羟基,因此脯氨酸修饰位置极大可能在11位羟基。
2.5 分子对接分析阿霉素及其两种代谢物与DNA的对接打分结果显示,阿霉素与DNA双链对接打分最好(-9.369 kcal·mol-1),m/z=641稍低(-8.358 kcal·mol-1),m/z=606最低(-7.191 kcal·mol-1),该结果表明相较于其代谢产物,阿霉素与DNA具有更强的亲和力。阿霉素与DNA对接的三维结构(Fig 5A)显示,阿霉素以一种平行于DNA双链结构的结构嵌入DNA双链中,该模拟结果与阿霉素嵌入DNA,导致DNA断裂和干扰DNA复制的实验机制相吻合,这种平行的结合方式使得阿霉素能够更深入的结合进DNA,从而产生较高的结合能力。而代谢物m/z=606及代谢物m/z=641的3D对接结果(Fig 5B、C)显示,这种平行结合方式在代谢物M1和M2与DNA的结合中丢失,相较阿霉素嵌入DNA的结合位点,代谢物M1和M2与DNA的结合位点均呈现出一定程度偏离,脱离了DNA双链,造成结合能力的下降。二维作用图表明(Fig 5a,b,c),阿霉素及其代谢物与DNA发生作用的位置主要在9位和14位的羟基,以及糖苷基团的氨基取代基上。3个小分子的糖苷基团都和DNA的碱基发生了作用,而差异之处主要是羟基和碱基发生的作用,m/z=606的代谢物的羟基没有和碱基发生作用,从而降低了结合能力,因此对接得分最低。而阿霉素的9位、14位以及蒽环结构的羟基分别与碱基发生了作用,而m/z=641的代谢物只有9位和14位羟基与碱基相互作用。综上所述,阿霉素与DNA具有独特的平行结合方式,该方式使得阿霉素与DNA较其代谢产物产生了更多的相互作用力,最终造成了阿霉素与DNA的高结合力。
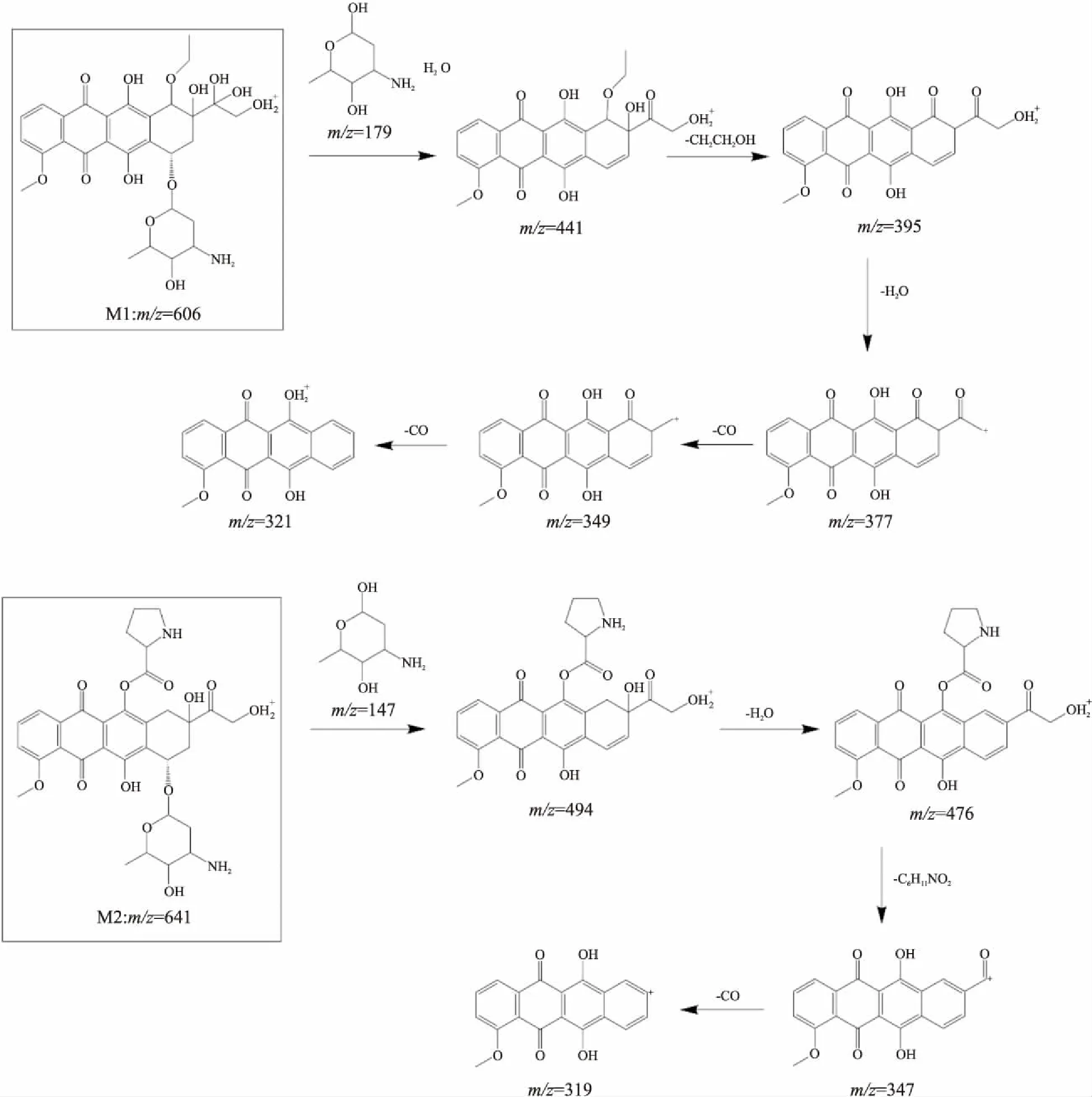
Fig 4 Fragmentation process and structure speculation of DOX and its metabolite M1(m/z=606) and M2(m/z=641)
3 讨论
临床使用中,阿霉素存在肿瘤细胞耐药以及心脏毒性的问题[10]。有研究发现阿霉素在 MCF7/WT野生型细胞中主要定位于胞核,而在 MCF7/DOX耐药细胞中则蓄积较少且主要定位于胞质[11]。核积聚的程度与这些细胞对阿霉素细胞毒性效应的敏感性直接相关。过去对阿霉素代谢的研究是基于肝脏的代谢,然而,很少见到靶细胞对阿霉素处置的研究报道。
本研究开发了一种靶细胞内微量代谢物的UPLC-Q-TOF-MS/MS分析方法,以鉴定靶细胞MCF-7耐药细胞和敏感细胞中的阿霉素代谢物,试验过程中未发现敏感细胞中的药物代谢物,对于耐药细胞中的阿霉素代谢产物,选用高分辨率多级质谱对其进行结构推导,发现了2个新的微量靶细胞阿霉素代谢物。新代谢物结构与体内药代动力学研究报道不一致,而与Wang等[9]发现的靶细胞阿霉素代谢物相关联,且质谱裂解过程具有共性,故推测靶细胞内的一系列相关代谢酶对阿霉素进行了修饰,而这些代谢过程与体内代谢途径有差异。
通过多级质谱对代谢物进行结构解析,计算机模拟分子对接比较了阿霉素与其靶细胞代谢物和DNA的结合能力,相对于阿霉素,这两个代谢物在DNA结合位点均呈现出一定程度偏离,且与DNA的亲和力相比阿霉素有所下降,提示靶细胞可能对阿霉素进行了特殊修饰,使得阿霉素代谢为药效更低的阿霉素代谢物,说明肿瘤靶细胞有独特的药物代谢途径,尤其存在于阿霉素耐药细胞中,可能与阿霉素的耐药机制有关联。

Fig 5 Molecular docking results of doxorubicin and its metabolites M1 and M2 with DNA
阿霉素体内生物转化的主要途径是由NADPH依赖性羰基(CBR)和醛酮(AKR)还原酶家族(统称为羰基还原酶)介导的,这些酶催化阿霉素代谢物阿霉素醇(DOXOL)的形成[12]。根据目前的质谱推测结果,不同于体内代谢过程,肿瘤靶细胞内阿霉素代谢过程可能有羟甲基酶、羟基化酶及氨基酸代谢相关酶参与。羟基化酶已经在红细胞、肝脏和肾脏中进行了广泛的研究[13],但未见其在肿瘤靶细胞中的研究。有研究表明,miR-142-3p的过表达可能抑制自噬靶向高迁移率族蛋白1(HMGB1),增强乳腺癌细胞对阿霉素的化学敏感性[14]。本课题组已有研究利用构建药物分子示踪探针,在包含20 000多种人类蛋白的芯片上筛选阿霉素结合蛋白,统计得出401个阿霉素潜在作用靶点[15]。将这些潜在靶点与耐药靶细胞阿霉素代谢物的代谢途径相结合,进一步研究阿霉素与这些鉴定的细胞靶标的确切相关性及其药理作用,就有可能从靶细胞药物代谢途径的角度解释阿霉素耐药机制,丰富对靶细胞对药物处置的认识。
由于细胞代谢产物的复杂性、药物代谢物含量低,很难分析和纯化代谢产物。受限于现有UPLC-MS/MS分析技术的灵敏度,MCF-7敏感细胞中是否有类似的阿霉素代谢物产生尚不清楚,阿霉素代谢物的代谢途径是耐药细胞特有还是靶细胞固有还有待进一步研究。由于阿霉素的特殊蒽环结构使其和代谢物便于观察,本研究针对阿霉素的靶细胞代谢物进行了研究,未来,我们还将使用基于阿霉素代谢物开发的方法来检测其他抗肿瘤药物的靶细胞代谢物,进一步完善方法学,使其成为可行和质量可控的方法,以补充现有药理学研究方法。
猜你喜欢
新传奇(2022年51期)2023-01-04
分子诊断与治疗杂志(2022年11期)2022-12-23
现代临床医学(2022年4期)2022-09-29
理化检验-化学分册(2020年5期)2020-06-15
农药科学与管理(2019年5期)2019-08-13
中国免疫学杂志(2019年3期)2019-03-13
神州·中旬刊(2019年1期)2019-02-12
医学研究杂志(2015年5期)2015-06-10
质谱学报(2015年5期)2015-03-01
中国药理学通报(2014年2期)2014-05-09
