
茶尺蠖触角高丰度气味结合蛋白EoblOBP9、EoblOBP11的配基结合功能和模式差异研究
2021-10-16 05:53严玉婷吴帆张亚丽傅晓斌崔宏春韩宝瑜李红亮
茶叶科学 2021年5期
严玉婷,吴帆,张亚丽,傅晓斌,崔宏春,韩宝瑜,李红亮
茶尺蠖触角高丰度气味结合蛋白OBP9、OBP11的配基结合功能和模式差异研究
严玉婷1,吴帆1,张亚丽2,傅晓斌1,崔宏春3,韩宝瑜1,李红亮1*
1. 中国计量大学生命科学学院/浙江省生物计量及检验检疫技术重点实验室,浙江 杭州 310018;2. 中华全国供销合作总社杭州茶叶研究院,浙江 杭州 310016;3. 杭州市农业科学研究院,浙江 杭州 310024
茶尺蠖(Prout)是茶园重要的鳞翅目害虫,雄雌成虫能通过嗅觉系统感受和分辨外界环境的化学信息以助于其交尾和产卵地选择等行为。在此过程中,成虫触角高丰度表达的气味结合蛋白(Odorant-binding proteins,OBPs)可能发挥了重要作用。以茶尺蠖成虫触角高丰度表达的两个OBPs即OBP9和OBP11为对象,首先发现二者cDNA编码蛋白均含有OBPs家族的典型特征,即含有6个保守的半胱氨酸,等电点预测分别为酸性和碱性,表明二者在一级结构性质上存在差异;利用原核表达技术获得重组蛋白,并免疫小鼠制备相应的多克隆抗体,再利Western blot技术验证了二者在雄性成虫触角特异表达;通过荧光竞争结合实验分别测试二者与20种候选配基的结合力,结果表明二者均与两个性信息素成分(顺3,9-环氧-6,7-十八碳二烯与顺-3,6,9-十八碳三烯)及一种植物挥发物成分(反-2-己烯醛)呈现较强的亲和力。与此同时,二者也显示出不同的配基结合谱,如OBP9与1-戊烯-3-醇、苯甲醇、苯乙酮和苯甲醛亲和力较强,结合谱较宽;而OBP11与-萜品醇亲和力较强,结合更显专一,分子对接结果也支持该结论。此外,预测二者的活性位点均位于C端,也显示出二者配基结合的共性。研究结果表明,尽管这两个茶尺蠖OBPs均在成虫触角高丰度特异表达,但无论从结构性质上还是生化结合谱上,既有保守性又存在差异性,显示出茶尺蠖触角高丰度表达OBPs功能模式的丰富度和多样性,这为解释茶尺蠖嗅觉系统在面对复杂多变的外界环境的适应机制提供了理论依据。
茶尺蠖;触角高丰度;气味结合蛋白;原核表达
在复杂的自然环境中,昆虫的嗅觉系统在其识别和感知外界化学信息分子过程中发挥了重要作用。在昆虫的嗅觉感器淋巴液中,充满了大量的球性蛋白,如气味结合蛋白(Odorant-binding proteins,OBPs)。OBPs能将外界化学气味分子运输至外周神经树突细胞膜上的气味受体(Olfactory receptors,ORs),触发电信号传递至脑部中枢,从而引发昆虫相应的行为和生理反应[1]。由此可见,作为嗅觉系统的第一个参与者,OBPs发挥筛选和运载的重要作用。按在鳞翅目感受器上分布的不同,可将OBPs分为普通气味蛋白(General odorant-binding proteins,GOBPs)、信息素结合蛋白(Pheromone-binding proteins,PBPs)和触角结合蛋白(Antennal binding protein X,ABPX)等[2],其中PBPs和GOBPs分别在和转运植物挥发物和结合昆虫信息素方面能发挥重要作用。
信息素和植物挥发物在昆虫定位、取食、觅偶和产卵等行为方面起到了重要的调控作用[3]。自1959年Butenandt等[4]首次在家蚕()雌蛾中鉴定出性信息素蚕蛾醇(Bombykol)以来,已经鉴定出3 000多种昆虫的性信息素[5]。作为茶树中发生最普遍、为害最严重的鳞翅目害虫之一,茶尺蠖(Prout)幼虫具有暴食性,严重时可使茶树光秃,严重影响茶叶产量和质量[6-7]。目前化学防治不仅破坏生态平衡和污染环境,而且容易使害虫抗药性增加,从而引起农药超标,因此利用信息素、植物挥发物生态调控技术等已成为茶尺蠖绿色防控的重要手段[8-10]。
茶尺蠖性信息素组分为主要性信息素和次要性信息素,分别为顺3,9-环氧-6,7-十八碳二烯(3,9-6,7-epo-18: Hy)和顺-3,6,9-十八碳三烯(3,6,9-18: Hy)[11]。另外某些植物挥发物也对茶尺蠖有引诱作用,如茶尺蠖成虫对寄主植物薄荷、迷迭香、茶叶和吸毒草等气味的趋性显著[12]。茶尺蠖幼虫危害茶树后释放植物挥发物苯甲醇、顺-3-己酸己烯酯和顺-3-己烯醛对茶尺蠖都有吸引作用,且加入顺-乙酸-3-己烯酯可增强吸引力[9,13]。除了引诱作用外,某些植物挥发物能产生驱避作用,如高剂量薰衣草气味能驱避茶尺蠖[12];芳香类植物迷迭香会释放顺-3-马鞭草烯醇、松油醇等植物挥发物,能够驱避茶尺蠖成虫产卵[14]等。尽管上述性信息素和植物挥发物具有引诱或者驱避茶尺蠖的能力已得到共识,但是尚不清晰茶尺蠖对这些结构不同的化学信息分子的嗅觉感受机制,且昆虫触角中的OBPs是识别传递外界化学信息的第一个重要环节,因此有必要对茶尺蠖OBPs及其功能展开深入研究。
已报道茶尺蠖OBPs家族包括24个成员,其中OBP9和OBP11在茶尺蠖成虫触角中呈高丰度表达[15],表明二者可能与茶尺蠖成虫嗅觉感受功能密切相关。为了进一步明确这两个触角高丰度表达基因的生理功能,本研究克隆了这两个基因的cDNA序列,在获得其蛋白的基础上,研究重组蛋白与性信息素组分及供试植物挥发物的结合谱,并通过分子对接解析和比较其结合机制,为茶尺蠖触角高丰度表达嗅觉感受机理提供参考,也为后续拓展开发新型生态调控信息素剂型奠定理论基础。
1 材料与方法
1.1 材料与主要试剂
茶尺蠖幼虫采自浙江省杭州市余杭区茶园,收集20头新羽化后0~2 d的茶尺蠖雄蛾触角,利用TRIzol提取RNA,反转录成cDNA后于–20℃冰箱保存备用。同时收集20只雄蛾的头、胸、腹、足和翅等组织用于Western blot。
pMD18-T载体、T4连接酶购于TaKaRa生物公司;pET-32a(+)载体为实验室制备保存;1-NPN购自梯希爱(TCI)公司(纯度>97%);顺3,9-环氧-6,7-十八碳二烯和顺-3,6,9-十八碳三烯购自北京中捷四方生物科技股份有限公司(纯度>90%);气味分子标准品购自上海百灵威化学技术有限公司(纯度>95%);其他均为进口或者国产分析纯试剂。
1.2 基因扩增与进化树分析
根据OBP9、OBP11基因序列(GenBank登录号分别为KT327214.1和KT327216.1)设计cDNA全长引物。以茶尺蠖雄虫触角cDNA第一链为模板扩增,程序为94℃3 min;94℃45 s,60℃45 s(每循环降落1℃,至50℃后再20个循环),72℃45 s,共35个循环。目的片段纯化后连接到pMD18-T载体上,选取阳性克隆测序验证。获得的序列经BLAST(Basic Local Alignment Search Tool)分析后利用MEGA 7.0软件进行序列比对和进化树构建(邻接法)。
1.3 蛋白表达纯化与免疫印迹
利用限制性酶切H和I对pMD18-T/OBP9、pMD18-T/OBP11进行双酶切,目的条带胶回收后,用T4连接酶与pET-32a(+)质粒连接,构建重组质粒,筛选阳性单克隆进行序列鉴定,然后诱导表达。用镍柱ProteinIsoTMNi-NTA Resin对目的蛋白进行分离纯化,SDS-PAGE检测后透析获得目的蛋白,免疫小鼠获得OBP9、OBP11多克隆抗体。茶尺蠖触角、头、胸、腹、足和翅各取20 µL的总蛋白溶液经SDS-PAGE电泳进行分离后,用Bio-Rad Transblot (Bio-Rad)将蛋白从胶转移至硝酸纤维素膜上;封闭液封闭120 min,TBST洗膜4次;再用制备的抗体(1︰500,/)4℃冰箱孵育过夜,TBST洗4次;再用HRP标记的羊抗兔二抗(1︰500,/)和膜常温静置6 h;按照超敏型辣根过氧化氢酶DAB显色剂盒的说明对底物进行显色。
1.4 荧光竞争结合实验
利用荧光分光光度计(岛津RF5301,日本)测试重组蛋白OBP9、OBP11与1-NPN的荧光光谱,确定1-NPN是否适合作为荧光报告子。用荧光分光光度计记录配基与1-NPN竞争结合重组蛋白的荧光数据,根据Scatchard方程[16]计算配基与蛋白的解离常数。
1.5 分子对接
从Swiss-Model Workspace中的蛋白三维晶体数据库(PDB),预测获得OBP9、OBP11的蛋白3D结构。从NCBI的PubChem中下载获得配基的3D结构。将蛋白和配基的3D结构一起导入到Molegro Virtual Docker(MVD 4.1,free trial)软件中,寻找最佳对接模型,最后用Pymol软件画出3D分子对接图。
2 结果与分析
2.1 序列比对及进化树分析
本研究成功获得了茶尺蠖OBP9、OBP11 cDNA序列全长,二者开放阅读框大小分别为426 bp和429 bp,编码141和142个氨基酸。其中OBP9编码氨基酸序列含有酸性残基达到20%(27个),呈酸性(预测等电点pI=4.3);而OBP11编码氨基酸序列含有碱性残基达到38.7%(55个),呈碱性(预测等电点pI=8.1)。
序列比对结果表明,二者均含有6个保守的半胱氨酸,具有OBPs家族典型的特点(图1),可形成3个二硫键,两者的二级结构主要为-螺旋。系统进化分析显示(图2),OBP9和OBP11处在两个分支上,其中OBP9与苹果银蛾OBP5(OBP5)相似度最高,达68.09%,OBP11与水稻稻纵卷叶螟GOBP2(GOBP2)相似度最高,达66.94%,表明二者存在序列进化差异。

注:EoblOBP9和EoblOBP11用箭头标注。BmorOBP1(家蚕OBP1);CsupOBP1(二化螟OBP1);DpleOBP1(黑脉金斑蝶OBP1);OfurOBP2(亚洲玉米螟OBP2);CmedGOBP2(水稻稻纵卷叶螟GOBP2);EoblOBP11(茶尺蠖OBP11);AconOBP5(苹果银蛾OBP5);EoblOBP9(茶尺蠖OBP9);MsexASP3(烟草天蛾ASP3);SinfOBP16(大螟OBP16);DhouOBP1(柳杉毛虫OBP1);DpunOBP44(松毛虫OBP44);LtriOBP1(三叶斑潜蝇OBP1);AcerASP2(中华蜜蜂ASP2);EoblPBP2(茶尺蠖PBP2);EoblGOBP1(茶尺蠖GOBP1);EoblGOBP2(茶尺蠖GOBP2)。下同
2.2 原核表达、蛋白纯化与Western blot分析
利用原核表达技术构建载体,并诱导获得OBP9和OBP11重组蛋白,其中OBP9主要存在于感受态细胞裂解液的上清液中,而OBP11主要存在于包涵体中,经镍柱纯化后获得二者的重组蛋白分子量大小分别为35.9 kDa和36.3 kDa(含pET-32载体标签)(图3)。
提取茶尺蠖雄成虫不同组织蛋白,SDS-PAGE电泳显示茶尺蠖雄虫腹部和胸部的总蛋白含量较高(分别为图4-A、图4-C中的泳道4和7),触角和翅的总蛋白含量相对较低(分别为图4-A、图4-C中的泳道2和6)。Western blot(图4-B、图4-D)显示,无论是触角(泳道2)还是重组蛋白(泳道8),均呈现明显特异条带,其他泳道中几乎没有,表明OBP9、OBP11蛋白特异地表达于茶尺蠖雄成虫触角中。
2.3 荧光竞争结合分析
OBP9与OBP11重组蛋白最大激发波长均为282 nm,最大发射波长均为335 nm。在蛋白溶液中逐渐增加1-NPN,记录发射光谱最大强度,并根据Scatchard方程,计算出二者与1-NPN的结合位点数n分别为(0.924±0.168)和(0.874±0.107),均接近于1;解离常数1-NPN分别为(6.603±2.351)、(4.412±1.544) µmol·L-1(图5-A、图5-B)。荧光竞争结合实验表明,在21种候选测试配基中,有7种配基可使OBP9蛋白荧光强度下降到50%以下,有5种配基可使OBP11蛋白荧光强度下降到50%以下(图5-C、图5-D),表明OBP9具有较OBP11更加广泛的配基分子识别谱。

图2 EoblOBP9和EoblOBP11与其他昆虫气味结合蛋白的NJ进化树

注:泳道1为蛋白分子量标准;泳道2、3和7、8分别为未经过和经过IPTG诱导的含有pET-32a(+)/EoblOBP9和pET-32a(+)/EoblOBP11重组质粒的细菌裂解产物;泳道4、5和9、10分别为IPTG诱导过的细菌裂解上清液和包涵体溶解液;泳道6和11分别为透析后的EoblOBP9和EoblOBP11蛋白溶液
如表1所示,OBP9与植物挥发物1-戊烯-3-醇结合能力最强(D=2.293 µmol·L-1),与茶尺蠖主要性信息素成分顺3,9-环氧-6,7-十八碳二烯和植物挥发物苯甲醇结合能力也较强(D<2.5 µmol·L-1);OBP11与茶尺蠖次要性信息素成分顺-3,6,9-十八碳三烯的结合能力最强(D=2.338 mol·L-1),其次是反-2-己烯醛(D=3.032 µmol·L-1)。
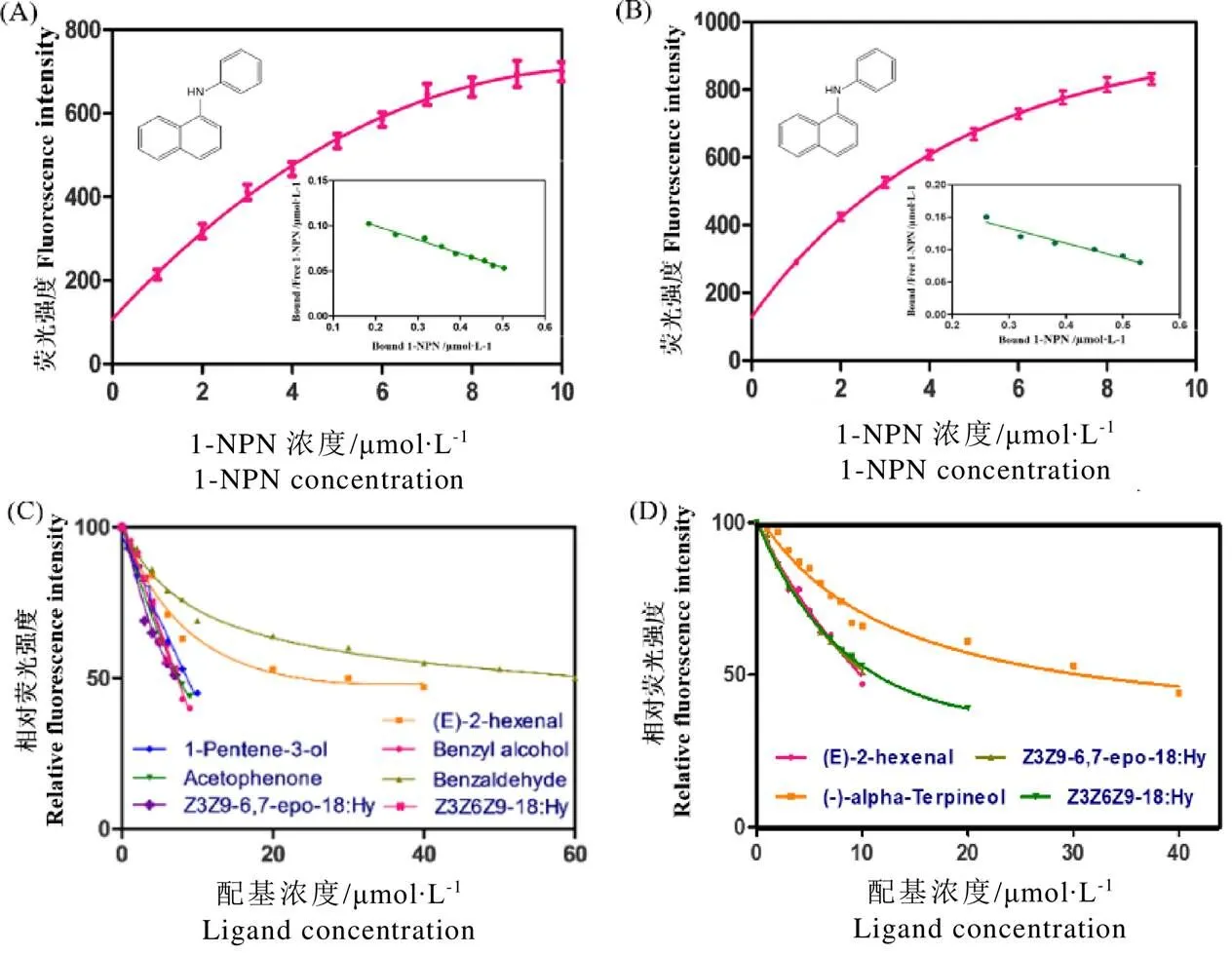
注:A和B:右上角图上有1-NPN的化学结构,根据Scatchard方程线性化荧光光谱数据,EoblOBP9、EoblOBP11与1-NPN的结合分析。C和D配基和1-NPN与EoblOBP9、EoblOBP11重组蛋白的竞争结合
2.4 茶尺蠖EoblOBP9和EoblOBP11与测试配体的分子对接分析
OBP9和OBP11预测的3D结构模型分别以冈比亚按蚊()的OBP1蛋白(2erb.1.A)和OBP20蛋白(4f7f.4.A)为模板进行构建,序列相似度分别为37.19%和37.82%,QMEAN值分别为–1.40和–0.77。
从分子对接结果看,OBP9与两个性信息素组分(顺3,9-环氧-6,7-十八碳二烯和顺-3,6,9-十八碳三烯)结合时提供主要能量的氨基酸残基有Ile33、Leu72、Tyr98、Arg101、Thr102、Leu137、Tyr138和Phe139(图6-A);OBP11与这两个性信息素组分结合时提供主要能量的氨基酸残基有Leu27、Gly31、Tyr71、Tyr95、Leu131、Met140、Phe141和Ala142(图6-B)。图6-C和图6-D显示OBP9与供试配基分子的结合腔较OBP11更大,暗示OBP9能容纳更多类型的气味分子和更宽的配基结合谱;而OBP11对气味的识别更具有专一性,结合谱较窄。对于植物挥发物来说,OBP9与苯甲醛、苯乙酮结合中提供更多能量为Tyr98、Thr102、Phe139,与反-2-己烯醛结合中提供更多能量为Tyr98、Leu137、Phe139,与苯甲醇的为Tyr98、Arg101、Leu137,与1-戊烯-3-醇的为Lys20、Asn56、Phe139;OBP11与反-2-己烯醛结合中提供更多能量为Val103、Leu131、Phe139,OBP11与-萜品醇结合中提供更多能量为Leu27、Phe141和Ala142。从分子对接三维图(图6-C、图6-D)来看,两个蛋白均有大量疏水性氨基酸在结合位点周围,从而形成疏水结合腔。此外,在所有测试配基中,发现除与顺-3,6,9-十八碳三烯结合无氢键外,其余配基与两个蛋白的对接模型中都含有氢键(图6-E、图6-F)。

表1 EoblOBP9和EoblOBP 11与候选配基的竞争结合分析

注:A是EoblOBP9与Z3, Z9-6R, 7S-epo-18: Hy、Z3, Z6, Z9-18: Hy、Benzaldehyde、(E)-2-hexenal、1-Pentene-3-ol、Benzyl alcohol、Acetophenone结合时发挥重要作用的氨基酸残基的能量值。图B是EoblOBP11与Z3, Z9-6R, 7S-epo-18: Hy、Z3, Z6, Z9-18: Hy、α-Terpineol、Dibutyl phthalate、(E)-2-hexenal结合中有重要作用的氨基酸残基的能量值。C图D:EoblOBP9和EoblOBP11与Z3, Z9-6R, 7S-epo-18: Hy分子对接的最佳模型,红色螺旋结构的3D蛋白结构与配体在疏水腔中相互作用。图E和图F:EoblOBP9和EoblOBP11与Z3, Z9-6R, 7S-epo-18: Hy的分子对接2D模型。黑色字体标注了关键氨基酸残基,青色虚线为氢键
3 讨论
昆虫OBPs是一类多功能蛋白,能作为载体结合和运输性信息素和气味分子到嗅觉神经树突末梢,引起昆虫嗅觉反应[17]。作为昆虫主要嗅觉感受器官,触角高丰度表达的OBPs极有可能参与昆虫嗅觉反应,本研究以触角高丰度表达的OBP9和OBP11为对象,来研究其与信息素和茶树挥发物的结合关系,并比较和解析了二者在配基生化结合功能的作用模式异同。
氨基酸序列和多序列联配分析显示,二者均具有昆虫OBPs典型序列特征(图1),且系统发育树显示二者能和多种昆虫OBPs具有较近的发育关系,然而二者也处在不同系统分支(图2)。有报道显示,OBP9和OBP11高丰度特异地表达于茶尺蠖成虫触角上[15],本研究利用Western blot技术进一步证实了两个蛋白能在茶尺蠖雄虫触角中特异表达,这表明二者极有可能参与了茶尺蠖雄成虫的嗅觉识别。在竞争性荧光结合实验中,两个蛋白均与顺3,9-环氧-6,7-十八碳二烯和顺-3,6,9-十八碳三烯这两种性信息素产生了较高的结合能力,甚至高于茶尺蠖PBP2[18],而且相较而言,OBP9与主要性信息素顺3,9-环氧-6,7-十八碳二烯结合能力强,而OBP11与次要性信息素顺-3,6,9-十八碳三烯结合能力强(表1),表明二者在参与茶尺蠖性信息素多元成分的识别过程中可能有互补的作用。
与供试茶树挥发物配基的结合实验中,OBP9和OBP11对不同的化学配基呈现极具规律性的配基结合谱。如对反-2-己烯醛,二者均会有较强的结和能力;然而,对1-戊烯-3-醇、苯甲醇、苯乙酮和苯甲醛,OBP9的亲和力较强,OBP11却不结合;对于-萜品醇,OBP11的亲和力较强,而OBP9却不结合(表1)。相比较而言,OBP9配基结合谱更宽,而OBP11更专一,分子对接结果也解释了可能的原因,即OBP9的结合腔较OBP11更大(图6-A和图6-B)。在前期研究中,发现茶尺蠖成虫触角中高表达的GOBP2也能与反-2-己烯醛、苯甲醛和苯乙酮等多种茶树挥发物结合[16],本研究中的OBP9与GOBP2显示了相似的配基结合谱,表明茶尺蠖OBPs具有与茶树挥发物亲和力强的共同特征。
在所测试的植物挥发性物质中,反-2-己烯醛作为植物受外界刺激诱导产生的一种绿叶性气味,不仅能对小贯小绿叶蝉()[19]和褐梗天牛()[20]产生引诱作用,而且还能刺激多音天蚕()雌蛾释放性信息素从而引诱雄蛾[21],可见反-2-己烯醛作为能够普遍引诱多种昆虫的重要植物活性成分,茶尺蠖这3种OBPs(GOBP2、OBP9、OBP11)与之都有较强的亲和力,表明茶尺蠖对该重要植物活性成分识别的高度协调性和一致性。对于另外的几种植物挥发物,如苯甲醛是在茶树受到茶蚜、茶尺蠖幼虫危害后可释放的,能吸引草瓢虫和天敌寄生蜂[22-23],只能与OBP9和GOBP2结合,而与OBP11不结合;-萜品醇是植物被寄主为害后自身产生的能够引诱寄生蜂等天敌[24],仅与OBP11的亲和力较强;而对其他的12种供试配基均不结合,表明不同茶尺蠖的OBPs对于不同的化学配基具有功能上的选择性,而这种选择性与蛋白的内部空间结构可能存在一定的关系。
对于等电点偏酸性的OBP9(pI=4.3)而言,本研究发现一个有趣的现象,即其结合位点除了多数的非极性氨基酸外,存在两个碱性氨基酸Arg101和Lys20(图6-A)。这种现象与东亚飞蝗()OBP1(pI=4.4)中的关键位点Arg10和His107的状况[25]非常相似,即在N端和C端分别有一个碱性氨基酸参与了配基结合,这个特征是否是所有酸性OBPs的共有的结合模式还需要进一步验证;而等电点偏碱性的OBP11(pI=8.1),只是和OBP9其他结合位点较类似,即活性中心分布着大量的非极性氨基酸残基(图6),而这些非极性残基形成的疏水性结合腔,能有效驱动与疏水性配基的结合[26]。此外,本研究显示,两个蛋白C端的部分残基,如OBP9的Leu137、Tyr138、Phe139和OBP11的Phe141、Ala142等,都参与了结合腔的生成(图6-A和图6-B),类似地情况也出现在冈比亚按蚊的OBP1[27]和茶尺蠖PBP2[28]中。另外,本研究发现,除了顺3,9-环氧-6,7-十八碳二烯外,OBP9与OBP11两个蛋白与其他亲和力较强的配基均预测能形成氢键,暗示氢键可能在二者与配基结合过程中发挥重要作用,这与草地螟()的GOBP1[29]和梨小食心虫()的GOBP2[30]与配基结合的情况也非常类似。
总而言之,本研究结果表明,作为两个茶尺蠖触角高丰度表达的OBPs,不仅显示了配基结合的普遍性规律,即与重要性信息素成分和关键植物挥发物成分的普遍高亲和力[1-2],而且也体现出了差异性的配基结合谱,这种差异与其空间结构密切相关[17],从而为茶尺蠖对植物寄主挥发物的嗅觉选择性机制提供了理论依据。
[1] Leal W S. Odorant reception in insects: roles of receptors, binding proteins, and degrading enzymes [J]. Annual Review of Entomology, 2013, 58: 373-391.
[2] Pelosi P, Zhou J J, Ban L P, et al. Soluble proteins in insect chemical communication [J]. Cellular and Molecular Life Sciences, 2006, 63(14): 1658-1676.
[3] Bruce T, Wadhams L J, Woodcock C M. Insect host location: a volatile situation [J]. Trends in Plant Science, 2005, 10(6): 269-274.
[4] Butenandt A, Beckmann R, Stamm D, et al. Über den sexual-lockstoff des seidenspinnersReindarstellung und Konstitution [J]. Zeitschrift für Naturforschg B, 1959, 14(4): 283-284.
[5] Symonds M, Elgar M A. The evolution of pheromone diversity [J]. Trends in Ecology & Evolution, 2008, 23(4): 220-228.
[6] 李喜旺, 刘丰静, 邵胜荣, 等. 茶尺蠖绿色防控技术研究现状及展望[J]. 茶叶科学, 2017, 37(4): 325-331.
Li X W, Liu F J, Shao S R, et al. Research progress and prospect of green control techniques of[J]. Journal of Tea Science, 2017, 37(4): 325-331.
[7] 高旭晖, 宛晓春, 杨云秋, 等. 茶尺蠖生物学习性研究[J]. 植物保护, 2007, 188(3): 110-113.
Gao X H, Wan X C, Yang Y Q, et al. Studies on the biological habits ofProut [J]. Plant Protection, 2007, 188(3): 110-113.
[8] Luo Z X, Li Z Q, Cai X M, et al. Evidence of premating isolation between two sibling moths:and(Lepidoptera: Geometridae) [J]. Journal of Economic Entomology, 2017, 110(6): 2364-2370.
[9] Sun X L, Li X W, Xin Z J, et al. Development of synthetic volatile attractant for malemoths [J]. Journal of Integrative Agriculture, 2016, 15(7): 1532-1539.
[10] 张帅琪, 冯博文, 张婧, 等. 灰茶尺蠖和茶尺蠖绿色防控技术研究进展[J]. 环境昆虫学报, 2020, 42(5): 1121-1138.
Zhang S Q, Feng B W, Zhang J, et al. Research progress on green control techniques ofWarren andProut [J]. Journal of Environmental Entomology, 2020, 42(5): 1121-1138.
[11] Yang Y Q, Zhang L W, Guo F, et al. Reidentification of sex pheromones of tea geometridProut (Lepidoptera: Geometridae) [J]. Journal of Economic Entomology, 2016, 109(1): 167-175.
[12] 江丽容, 刘守安, 韩宝瑜, 等. 7种寄主和非寄主植物气味对茶尺蠖成虫行为的调控效应[J]. 生态学报, 2010, 30(18): 4993-5000.
Jiang L R, Liu S A, Han B Y, et al. The adaptive strategies of insects to plant alleochemicals [J] Acta Ecologica, 2010, 30(18): 4993-5000.
[13] Sun X L, Wang G C, Gao Y, et al. Volatiles emitted from tea plants infested bylarvae are attractive to conspecific moths [J]. Journal of Chemical Ecology, 2014, 40(10): 1080-1089.
[14] Zhang Z Q, Bian L, Sun X L, et al. Electrophysiological and behavioural responses of the tea geometrid(Lepidoptera: Geometridae) to volatiles from a non-host plant, rosemary,(Lamiaceae) [J]. Pest Management Science, 2015, 71(1): 96-104.
[15] Ma L, Li Z Q, Bian L, et al. Identification and comparative study of chemosensory genes related to host selection by legs transcriptome analysis in the tea geometrid[J]. PLoS One, 2016, 11(3): e0149591. doi: 10.1371/journal.pone.0149591.
[16] 赵磊, 崔宏春, 张林雅, 等. 茶尺蠖普通气味结合蛋白EoblGOBP2与茶树挥发物的结合功能研究 [J]. 茶叶科学, 2014, 34(2): 165-171.
Zhao L, Cui H C, Zhang L Y, et al. Molecular binding characterization with tea plant volatiles of a general odorant-binding proteinGOBP2 in the tea geometrid,Prout (Lepidoptera: Geometridae) [J]. Journal of Tea Science, 2014, 34(2): 165-171.
[17] Pelosi P, Iovinella I, Zhu J, et al. Beyond chemoreception: diverse tasks of soluble olfactory proteins in insects [J]. Biological Reviews, 2017, 93(1): 184-200.
[18] 冯一璐, 傅晓斌, 吴帆, 等. 茶尺蠖信息素结合蛋白PBP2的基因克隆、原核表达及其结合功能[J]. 中国农业科学, 2017, 50(3): 504-512.
Feng Y L, Fu X B, Wu F, et al. Molecular cloning, prokaryotic expression and binding functions of pheromone binding protein 2 (PBP2) in the[J]. Scientia Agricultura Sinica, 2017, 50(3): 504-512.
[19] Chen K, Huang M X, Shi Q C, et al. Screening of a potential leafhopper attractants and their applications in tea plantations [J]. Journal of Environmental Science and Health Part B, 2019, 54(10): 1-7.
[20] 泽桑梓, 赵宁, 王大纬, 等. 反-2-己烯醛对褐梗天牛成虫引诱剂的作用效率[J]. 中国森林病虫, 2013, 32(3): 46.
Ze S Z, Zhao N, Wang D W, et al. Effect of ()-2-hexenal on attractant efficiency of longicorn beetle[J]. Forest Pest and Disease, 2013, 32(3): 46.
[21] 杜家伟. 昆虫信息素及其应用[M]. 北京: 中国林业出版社1988.
Du J W. Insect pheromone and its application [M]. Beijing: China Forestry Press, 1988.
[22] Maeda T, Ishiwari H. Tiadinil, a plant activator of systemic acquired resistance, boosts the production of herbivore-induced plant volatiles that attract the predatory mitein the tea plant[J]. Experimental and Applied Acarology, 2012, 58(3): 247-258.
[23] 陈宗懋, 许宁, 韩宝瑜, 等. 茶树-害虫-天敌间的化学信息联系 [J]. 茶叶科学, 2003, 23(s1): 38-45.
Chen Z M, Xu N, Han B Y, et al. Chemical communication between tea plant-herbivore-natural enemies [J]. Journal of Tea Science, 2003, 23(s1): 38-45.
[24] Zhu J, Cossé A, Obrycki J J, et al. Olfactory reactions of the twelve-spotted lady beetle,and the green lacewing,to semiochemicals released from their prey and host plant: electroantennogram and behavioral responses [J]. Journal of Chemical Ecology, 1999, 25(5): 1163-1177.
[25] Zheng J, Li J, Han L, et al. Crystal structure of theodorant binding protein [J]. Biochemical Biophysical Research Communications, 2015, 456(3): 737-742.
[26] Liu H, Duan H, Wang Q, et al. Key amino residues determining binding activities of the odorant binding protein AlucOBP22 to two host plant terpenoids of[J]. Journal of Agricultural and Food Chemistry, 2019, 67(21): 5949-5956.
[27] Wogulis M, Morgan T, Ishida Y, et al. The crystal structure of an odorant binding protein from: Evidence for a common ligand release mechanism [J]. Biochemical Biophysical Research Communications, 2006, 339(1): 157-164.
[28] Fu X B, Zhang Y L, Qiu Y L, et al. Physicochemical basis and comparison of two Type II sex pheromone components binding with pheromone-binding protein 2 from tea geometrid,[J]. Journal of Agricultural and Food Chemistry, 2018, 66(50): 13084-13095.
[29] Yin J, Zhuang X, Wang Q, et al. Three amino acid residues of an odorant-binding protein are involved in binding odours inL [J]. Insect Molecular Biology, 2015, 24(5): 528-538.
[30] Li G W, Chen X L, Li B L, et al. Binding properties of general odorant binding proteins from the oriental fruit moth,(Busck) (Lepidoptera: Tortricidae) [J]. PLoS ONE, 2016, 11(5): e0155096. doi: 10.1371/journal.pone.0155096.
Study on the Differences in Ligand-binding Function and Mode of the Antennal High-abundance Odorant-binding ProteinsOBP9 andOBP11 of the Tea Geometrid,Prout
YAN Yuting1, WU Fan1, ZHANG Yali2, FU Xiaobin1, CUI Hongchun3, HAN Baoyu1, LI Hongliang1*
1. College of Life Science, China Jiliang University/Zhejiang Provincial Key Laboratory of Biometrology and Inspection and Quarantine, Hangzhou 310018, China; 2. Hangzhou Tea Research Institute, China Coop., Hangzhou 310016, China;3. Hangzhou Academy of Agricultural Sciences, Hangzhou 310024, China
As an important lepidopteran pest in tea gardens, the male and female adults ofProut can perceive and distinguish the chemical information of the external environment through the olfactory system, which helps its behaviors such as mating and selection of oviposition locations. In this process, Odorant-binding proteins (OBPs) highly expressed in the antennae of adults may play an important role. In view of this, this study focused on the two OBPs high expressed in the antennae of the adult tea geometrid, namelyOBP9 andOBP 11. First, it was found that bothOBP9 andOBP11 contain six conserved cysteines, the typical characteristics of the OBPs family, while the isoelectric points are predicted to be acidic and basic, respectively, indicating their difference in the primary structural properties. Both recombinant proteins were obtained using prokaryotic expression technology, and the corresponding polyclonal antibodies were prepared by immunizing mice. It was verified that they were indeed expressed specifically in the antennae of male adults by Western blot. Their binding abilities with 20 candidate ligands were tested by fluorescence competitive binding experiments, and the results show that both of them were compatible with the two sex pheromone components (3,Z9-6,7-epo-18: Hy and3,6,9-18: Hy) and a plant volatile component (trans-2-hexenal) has a strong affinity. Meanwhile, two OBPs also showed different ligand binding spectra. For example,OBP9 had a strong affinity with 1-penten-3-ol, benzyl alcohol, acetophenone and benzaldehyde, indicating that the binding profiles were wider; whileOBP11 had a strong affinity with-terpineol with a more specific binding mode. Molecular docking analysis also supports this conclusion. In addition, it was predicted that the active sites of both proteins are located at the C-terminus, which also shows the commonality of ligand binding. In conclusion, this study shows that although both OBPs were specifically expressed in the adult tea geometrid with high abundance, they are both conservative and different in terms of structural properties and biochemical binding profiles. The general characters and significant differences are coexist, showing the high abundance and diversities of the OBPs. This can also explain an adaptation mechanism of the tea geometrid olfactory system in the face of the complex and changeable external environment.
Prout, antennal high-abundance, odorant-binding protein, prokaryotic expression
S571;S435.711
A
1000-369X(2021)05-643-11
2021-01-26
2021-04-05
国家自然科学基金(31772544、32000331)、浙江省自然科学基金(LQ21C030007)、科技部基础资源调查专项(2018FY100405)、浙江省重点计划项目(2020C02026)
严玉婷,女,硕士研究生,主要从事昆虫生化与分子生物学方面的研究。*通信作者:hlli@cjlu.edu.cn
(责任编辑:赵锋)
猜你喜欢
世界热带农业信息(2022年8期)2022-07-19
茶叶科学(2021年4期)2021-08-12
华人时刊(2020年19期)2021-01-14
小资CHIC!ELEGANCE(2019年40期)2019-12-10
小猕猴学习画刊(2019年4期)2019-05-08
扬子江(2019年1期)2019-03-08
爱你(2017年13期)2017-11-15
作文评点报·作文素材小学版(2016年9期)2016-03-18
农民致富之友·下半月(2015年6期)2015-07-09
中学英语之友·高一版(2008年1期)2008-03-20
