
对热带玉米种质群体控制双亲混合选择适应性的评价
2021-10-21 01:34张艳茹杨子和焦金龙吴元奇
作物杂志 2021年5期
张艳茹 杨子和 杨 荣 韩 建 焦金龙 赵 丽 吴元奇
(1四川农业大学玉米研究所,611130,四川成都;2雅安市农业科学研究所,625000,四川雅安;3蓬安县农业农村局,637800,四川南充)
玉米是我国重要的作物,用途广泛,可以用于粮食、能源和饲料等。长期以来中国玉米种质主要集中在来自美国的 Lancaster、Reid和国内塘四平头、旅大红骨4种类型[1-2],为了加速品种选育,常用的做法是把少数几个骨干自交系或其衍生系作为育种的基础材料反复地利用,导致近十几年来玉米育种遗传增益逐渐降低,使玉米对病虫害和逆境表现出脆弱性,减少了提高杂种优势的潜力和增强抗逆能力的可能性,影响玉米生产的可持续发展[3]。因此,解决种质基础狭窄问题的有效方法就是拓宽玉米种质基础,将热带、亚热带玉米种质导入温带玉米种质中,增加玉米遗传多样性。热带、亚热带种质在干旱、瘠薄等严酷的环境条件下经过长期的自然和人为选择,具备了耐旱、耐高温阴雨、茎秆坚韧、抗倒性强和抗病虫害等优良特性,且与其他种质存在地理隔离,遗传交流较少,因而拥有丰富的遗传变异[4]。因此,通过引进外来种质,创建优良群体和杂种优势模式[5],对我国玉米育种具有重要意义。
热带种质对光具有较强敏感性,为了使热带和亚热带种质适应温带日照条件,必须进行适应性选择[6],减少不利效应。要改良外来种质适应性,必须保持较大的有效群体和适当降低选择强度,才能够保持群体遗传变异性,通常采用混合选择法满足上述要求,该方法不仅可以保持群体中优良变异,同时可以防止近交衰退,包括籽粒产量、开花时间、果穗长度、株高、籽粒含水量等。美国学者20世纪60年代开始引进并驯化了一批具有热带血缘的玉米种质群体,目的是为了丰富玉米带的种质基础。有研究[7-10]对美国近年来应用热带玉米种质资源情况进行了总结,指出热带种质资源的应用极大地丰富了玉米育种材料的遗传基础,并选出一批符合美国玉米带育种目标的自交系,经杂交试验表明这批自交系有较好的适应性和较高的配合力。近年来美国利用 50%的热带种质与 50%的温带种质杂交,极大地拓宽了美国玉米的遗传基础,依阿华州立大学对引进的热带玉米种质群体进行6轮混合选择,改良得到群体 BS16,不仅适应在美国地带生长,并且具有较高的产量和配合力[11]。
国内热带和亚热带玉米种质导入利用的研究始于20世纪80年代初。当时从墨西哥(CIMMYT)和泰国等引入了Tuxpeno、Suwan1、Across和Pool33等一批热带和亚热带材料,将温带材料与之杂交组建了温热杂种群体,并对其进行改良,直接或间接地选育出了一批新的自交系,并且开始构建新的杂交模式,通过适应性改良、农艺性状和配合力测定,筛选出适合我国利用的优良群体,如中综5号、中综6号、中综7号、BSSS和BS13等[12-13],拓宽了我国的玉米种质基础。四川农业大学选育的S37、山西省农业科学院育成的太113自交系、河南农业大学选育的 87-1、吉林省农业科学院选育的 8501和沈阳市农业科学院选育的沈118、沈218、沈219等自交系都含有热带、亚热带种质血源,很多自交系成为了育种单位主要的骨干自交系。
刘代惠等[14]、黎裕等[15]和董玲等[16]经过分析与整合,已经形成了较为系统的热带、亚热带玉米种质的利用途径。其中群体改良和轮回选择是改良利用热带、亚热带种质的有效方法。对外来种质进行适应性改良时,一般采用适度规模的控制双亲的混合选择法,该方法可钝化热带和亚热带种质对光周期的敏感性,同时最大程度地保留其原有的遗传变异。具体操作是在抽丝、散粉期间对雌穗和雄穗进行套袋,即对雌穗和雄穗同时做了选择。中国农业科学院通过5轮混合选择,改良了2个亚热带优质蛋白玉米群体Pool33和Pool34在我国北方的适应性,选育出优质蛋白玉米自交系 CA375和杂交种中单9409。从1997年开始,采用混合选择法改良CIMMYT群体,经过3年3轮光周期驯化,其主要农艺性状和产量性状得到改良,Pob43和Pob21等群体基本适应了我国南方及河南等地区的生态条件。郭向阳等[17]通过对玉米Tuxpeno轮回选择来拓宽我国玉米的种质基础。
本研究利用1份非洲农家地方种,将合成基础群体 AC0以及经 2轮控制双亲混合选择改良的群体作为供试材料,评价玉米群体主要性状的遗传变异,为该群体的有效利用提供参考依据。
1 材料与方法
1.1 材料来源
基础玉米材料是来自西非中南部国家贝宁的1份农家地方种,属于热带种质。贝宁全境地处热带,全年高温。
1.2 群体选择方法和程序
2017年3月,在四川雅玉科技有限公司新津育种基地种植,淘汰发育不良、结实率低的个体,单株自交,选取穗部表现良好的果穗,等量取穗中部的饱满玉米籽粒,混匀后组成AC群体。同年9月,在海南乐东育种基地种植AC群体,选取最早抽丝的植株,人工控制混合授粉,收获时自然淘汰不孕株和穗腐病严重的果穗,混合脱粒为AC0群体。
2018年3月,在四川新津育种基地种植AC0(S1)群体2000~3000株,并采用控制双亲混合选择的方法,选取2块面积相同地块分别种植1000~1500株,人工控制授粉,选取雌雄协调的植株进行授粉,每一轮群体内的花粉来源不少于250株,对AC0群体进行改良,选取结实率较高玉米穗,淘汰不良种质,获得AC1(S2)群体2000~3000株。同年9月,在海南育种基地种植AC1群体,采用相同的改良程序对AC1群体进行改良,得到AC2(S3)群体2000~3000株,在2018年9月AC0群体已经完成2轮选择。
2017年3月,在四川雅玉科技有限公司新津育种基地,根据AC0群体幼苗长势、雌雄协调性、穗型、行粒数、粒型、结实性等农艺性状选出21份优良植株,自交且单株收获,获得S1;同年9月,在海南育种基地种植S1,继续让S1自交,收获后得S2,共21份;2018年3月,在新津育种基地种植S2,继续让S2自交,淘汰11份后收获S3,获得10份,后续再进行选育,自交至少6次以上得到较纯的自交系,用于构建杂种优势模式,增加种质多样性。
1.3 试验设计
2019年 3月在四川农业大学崇州(103°07'-103°49' E、30°30'-30°53' N)基地和安岳(104°56' E、29°40' N)基地种植AC0、AC1和AC2群体,采用随机区组排列,3次重复,10行区,行长4.5m,行距0.7m,密度为57 000株/hm2,常规大田管理。待AC0、AC1、AC2群体株高稳定,3个群体在崇州的株高、穗位高分别调查149株、159株、292株。收获时在崇州、安岳各群体的穗数调查分别为171株、170株、145株,对收获植株进行室内考种,调查穗长、秃尖长、穗行数、行粒数、穗粗、轴粗、粒深、穗重、轴重和单株产量。
1.4 数据分析
以小区平均值进行方差分析;采用DPS 7.55软件处理分析数据。按公式计算单株各性状的变异系数。
按公式∆Gi(%)=(Xsi–Xi)/Xi×100 计算各性状的遗传增益[18],式中∆Gi为i性状的遗传增益,Xsi为改良群体i性状的平均数,Xi为某基础群体i性状的平均数。
2 结果与分析
2.1 群体性状的方差分析
从表1可以看出,基础群体AC0以及改良群体AC1、AC2群体的株高、穗位在群体间差异达到显著水平,其余性状均未达到显著水平,说明性状遗传不稳定,易受环境的影响,基础群体和其改良群体部分性状间存在遗传差异。
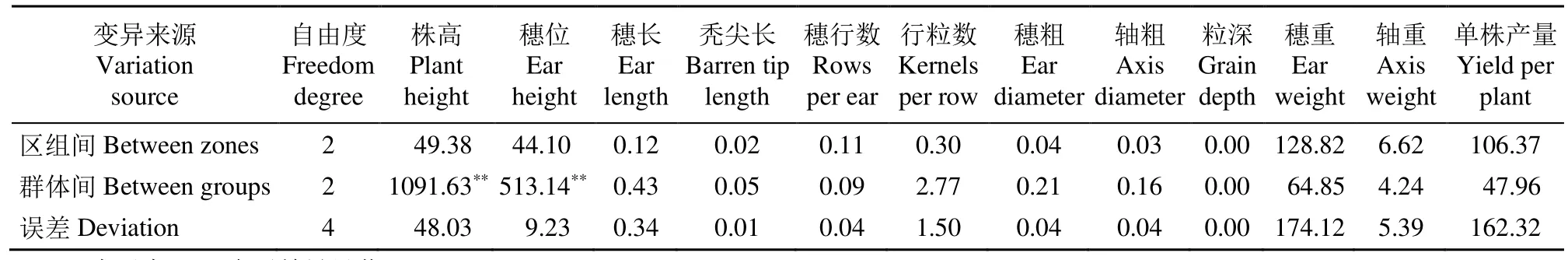
表1 群体各性状方差分析结果Table 1 The results of the analysis of variance of the various symptoms of the groups
2.2 混合选择方法对群体主要农艺性状改良效果评价
2.2.1 株高和穗位的改良效果评价 从表2可以看出,经过2轮控制双亲混合选择改良后,基础群体AC0株高和穗位高呈极显著增加趋势。株高在AC2为最大值,与 AC0间有极显著差异,遗传增益为11.60%,;穗位性状与株高相同,在AC2为最大值,且 AC0与 AC2间差异达到极显著水平,其遗传增益为16.18%。
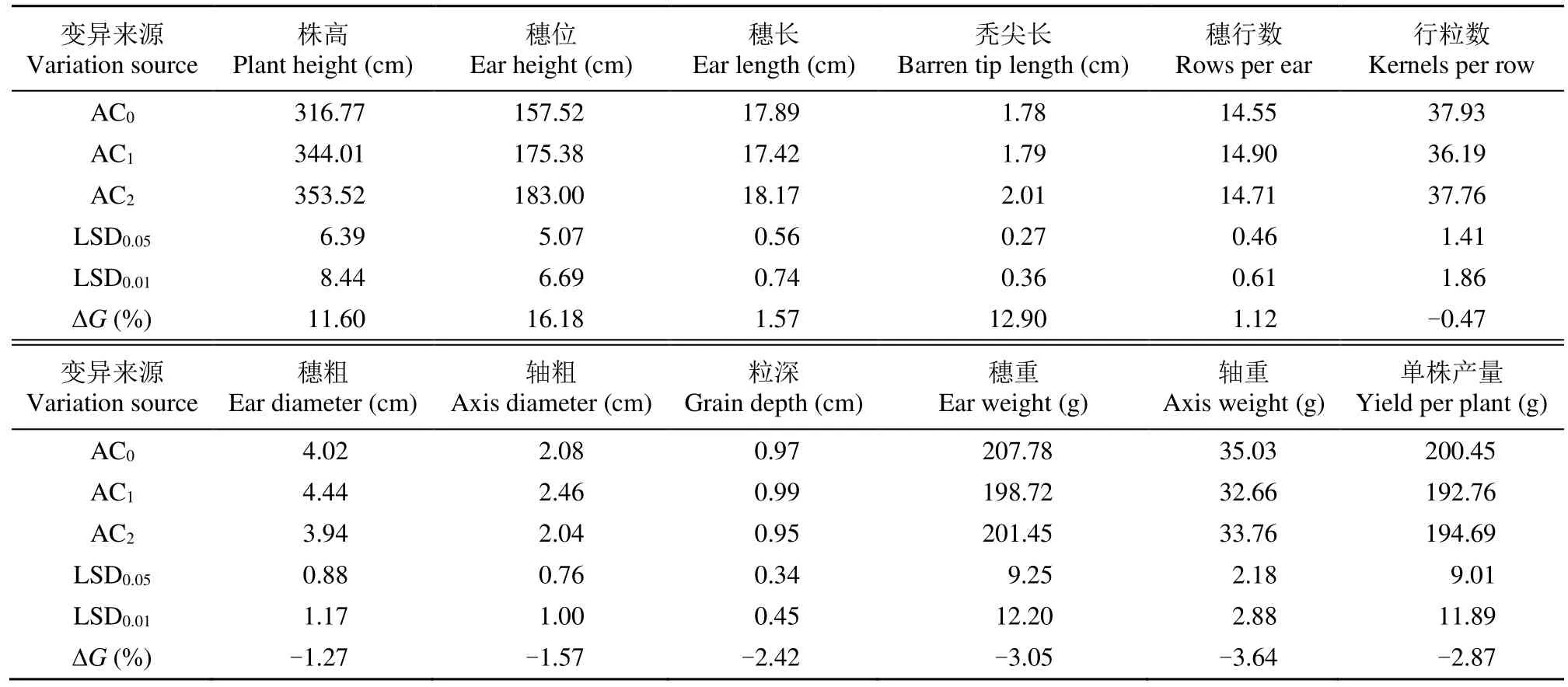
表2 主要农艺性状群体间的均值比较Table 2 Comparison of the averages the main agronomic traits among of groups
2.2.2 穗长和秃尖长的改良效果评价 从表 2可以看出,经过2轮控制双亲混合选择改良后,群体AC0穗长和秃尖长有一定程度增加。其中最大的穗长表现在AC2,AC2与AC1达到极显著性差异,遗传增益为1.57%,表现为有效遗传;秃尖长最大值同样出现在 AC2,无显著性差异,其遗传增益为12.90%,表现为无效遗传。
2.2.3 穗行数和行粒数的改良效果评价 从表2可以看出,经过2轮控制双亲混合选择改良后,对于基础群体AC0,穗行数有一定程度的增加,行粒数略微减少。穗行数最大值出现在AC1,但无显著性差异,遗传增益为1.12%,表现为有效遗传;行粒数最优值出现在AC0,与AC1有显著差异,遗传增益为-0.47%,表现为无效遗传。
2.2.4 穗粗、轴粗和粒深的改良效果评价 从表2可以看出,经过2轮控制双亲混合选择改良后,基础群体AC0穗粗、轴粗和粒深均略微降低。穗粗在AC1有最优值,与AC0和AC2差异不显著,AC2最小,遗传增益为-1.27%,表现为无效遗传;轴粗最大值也在AC1出现,分别与AC0、AC2差异不显著,遗传增益为-1.57%,表现为无效遗传;粒深最大值同样是AC1,与AC0和AC2差异不显著,遗传增益为-2.42%,表现为无效遗传。
2.2.5 穗重、轴重和单株产量的改良效果评价 从表 2可以看出,经过 2轮控制双亲混合选择改良后,基础群体AC0穗重、轴重、单株产量也均略微降低。穗重最优值在AC0,但与其他群体无显著性差异,遗传增益为-3.05%,表现为无效遗传;轴重最大值也在AC0,与AC1达到显著差异,遗传增益为-3.64%,表现为无效遗传;单株产量最大值同样在AC0,与其他群体无显著差异,遗传增益为-2.87%,表现为无效遗传。
2.3 改良群体性状的遗传变异分析
变异系数在一定程度上可以揭示群体内个体间的遗传变异,图1中AC0基础群体经过2轮控制双亲混合选择后,在崇州,穗位、轴粗、穗粗、粒深、轴重、单株产量性状在群体间的变异系数在改良群体间减小,表明各性状在改良群体AC2趋于稳定,但穗长、穗重、行粒数、株高、秃尖长和株高性状的变异系数在 AC0、AC1、AC2群体间增加,表明性状在改良群体AC2遗传不稳定。在安岳,穗长、行粒数、穗重、株高、穗位和秃尖长性状的变异系数 AC0基础群体相比在 AC2代有所降低;穗行数、穗粗、轴粗、粒深、单株产量5个性状的变异系数在群体间都有所增加。
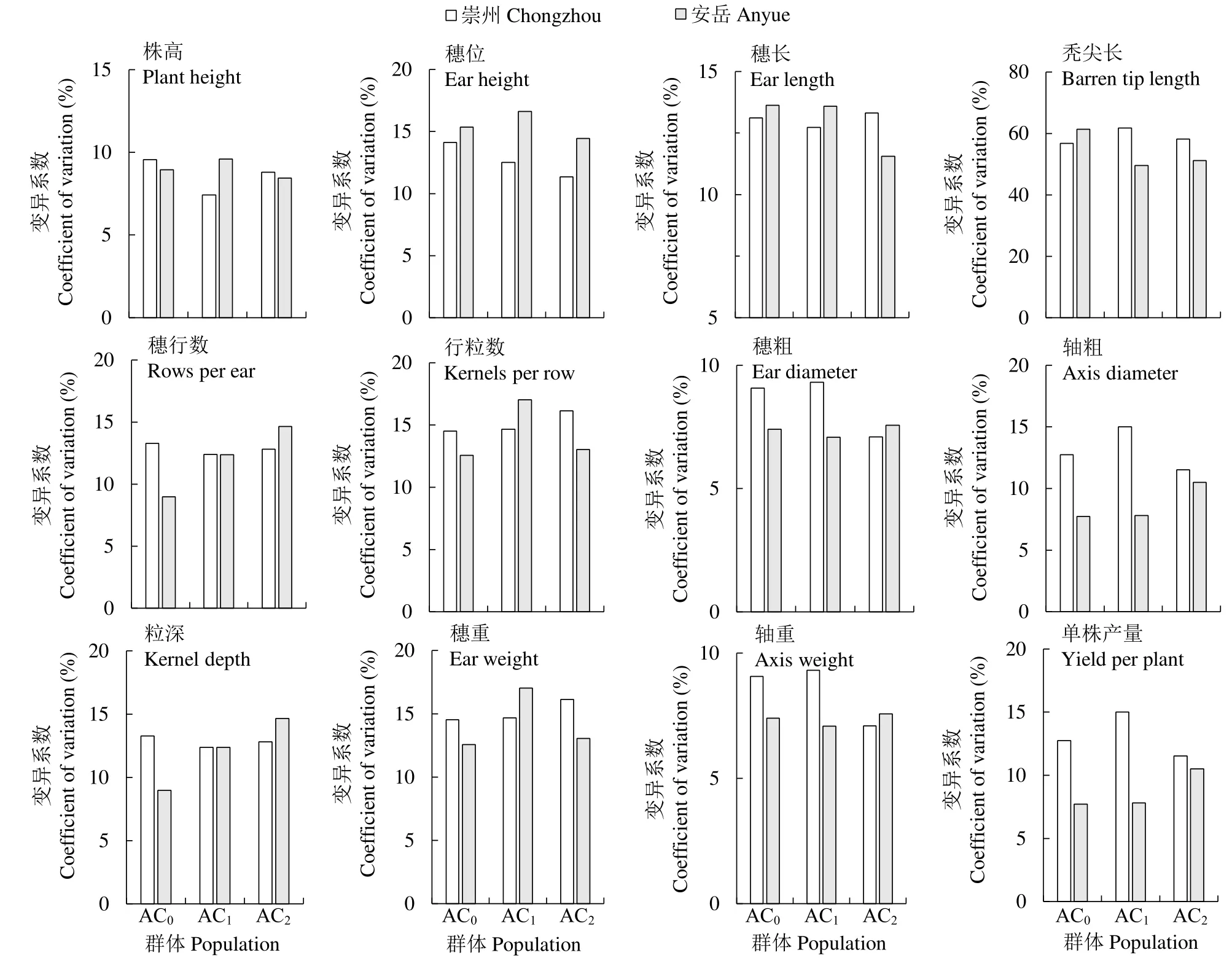
图1 主要农艺性状的变异系数Fig.1 Coefficients of variation of the main agronomic traits
不同地点各性状变异系数基本都有差异,说明其受环境影响较大,轮回群体次数较少,各性状不能够稳定遗传。由于作物性状有质量性状和数量遗传,质量性状为单个主基因控制,呈不连续变异,能够明确分组,数量性状为多个基因控制,在遗传分离群体中表现为连续性变异且不能够明确分组。数量性状相比质量性状,在不同环境中变幅较大,如经典数量遗传学表明,株高是由多基因控制[19],前人研究[20]表明,玉米产量的构成是与许多农艺性状相关,由多基因控制,并受环境和基因型共同影响,因此产量部分性状在不同地点间表现变化幅度较大。
3 讨论
通过群体间性状方差分析可知,AC0群体的株高、穗位高、穗粗、轴粗在AC1、AC2群体中相差较大,说明选取的自交种农艺性状受环境影响较大,遗传不稳定。AC0基础群体通过2轮选择后,大多农艺性状均值都相对AC0群体有所变化,证明改良措施是有效的,选取的自交种遗传效应较强,这与庄铁成等[21]认为早期混合选择对外来种质产量性状的改良进展较小类似,但它们均值变化幅度不大,且某些性状在AC1出现最优值,可能与在四川新津和海南乐东育种基地2种生态环境下选择有关。试验结果也表明在改良过程中应及时鉴定改良效果,以便根据育种需要和目标性状的表现,及时调整群体改良的选择强度,以提高群体改良的效果。
群体改良中由于群体间遗传背景的不同,表现出不同的遗传特性,导致不同群体改良用途不同。张德贵等[22]研究表明,控制双亲混合选择方法能够有效改良群体的部分性状,有的生育期得到了改良,有的产量及其相关性状没有明显的改良,改良效果因材料的不同而异。本研究结果表明,经过2轮控制双亲混合选择后,基础群体AC0自身穗长和穗行数性状得到一定程度的改良,但也伴随着株高、穗位高、秃尖的升高,这与李芦江等[23]研究中P4群体、P5群体经过 2轮控制双亲混合选择后株高、穗位改良效果趋势一致,并且认为株高和穗位升高可能是光周期敏感引起的,随着改良不断推进,应该会有所降低,其需要后期增加轮回次数,进一步对群体中优良性状的选育,为后期组合杂交种奠定基础。如杨致远等[24]通过外来种质的渗入,选出CP15-2为优良种质,可以用于改良生育期、产量和含水量等性状。
热带、亚热带种质由于其强烈的温带光周期和温度敏感性,会掩盖优良基因表达和特异性状的表现,采用混合选择法直接改良热带、亚热带种质难度较高、耗费时间较长,但可以减少优良种质的丢失,因此需要较大的改良群体,打破不良基因的连锁,确保群体内遗传的多样性,本试验通过2轮改良,群体较多农艺性状的增益效应都呈现负值,单株产量的变异系数增大,说明以AC0为基础群体的改良,没有达到增产的目的,可能由于环境和群体大小的影响,其优良基因未能表现出来,后续应增加改良群体和次数,发掘优良性状,达到增产和增加优良种质的目的。
4 结论
选用的非洲种质有一定的育种潜力,但因为改良轮次较少,目前还看不出大多数性状的改良趋势。在今后的改良过程中,将增加改良轮回次数和群体选择强度,同时兼顾其他性状,集聚群体的优良基因,形成综合性状较好的育种群体。
猜你喜欢
作物学报(2022年8期)2022-05-29
作物杂志(2022年6期)2022-02-03
东坡赤壁诗词(2020年5期)2020-11-06
湖南农业大学学报(自然科学版)(2020年4期)2020-08-28
北京农学院学报(2019年1期)2019-02-22
山西农业大学学报(自然科学版)(2018年12期)2018-12-04
河南农业科学(2017年4期)2017-04-12
小学生时代·综合版(2016年4期)2016-11-19
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年6期)2016-04-16
