
不同作物光合蛋白复合物的提取及比较分析
2021-10-21 01:34王儒情
作物杂志 2021年5期
王儒情 华 玮 刘 军
(1中国农业科学院油料作物研究所/农业农村部油料作物生物学与遗传育种重点实验室,430062,湖北武汉;2中国农业科学院研究生院,100081,北京)
粮油安全是涉及社会、国家与个人等多个层面的问题。近50年来,通过提高作物收获指数,粮油作物产量得到大幅提高[1]。然而当前作物收获指数已接近极限,大幅提高收获指数可能性并不大;此外,随着人口增长和城市化的推进,大幅增加耕地面积可能性也不存在[1-2];光合作用是植物维持生命的主要能量来源,光能转化效率理论值约4%,但是实际值仅约1%,光合作用仍有很大提升空间,进一步改良光能利用效率或成为未来大幅度提高作物产量重要的甚至是唯一可行的途径[3-5]。并且,改良光能利用效率还可以实现逆境胁迫条件下的作物增产[6]。光能捕获、吸收和传递等光反应在类囊体膜上进行,类囊体膜主要由光系统I(PSI)、光系统II(PSII)、细胞色素b6f(Cytb6f)复合体和 ATP 合酶复合体(ATPase)4种多亚基整合膜蛋白复合物组成[7]。因此,揭示不同作物类囊体膜复合物之间的异同点对改良光合作用具有重要的理论指导意义。
作物根据光合作用类型主要分为 C3型(如水稻、烟草)和C4型(如玉米、白花菜)[8];根据子叶数目分为单子叶作物(如水稻、玉米)和双子叶作物(如烟草、白花菜);根据染色体倍性分为二倍体作物(如烟草、水稻)和多倍体作物(如油菜、大豆、花生)。目前关于作物光合作用机制的研究已从形态结构、生理生化和细胞水平深入到蛋白质与基因的分子水平,而类囊体膜作为植物光反应的场所,探究其组成特征对分析作物的光合特性、改良育种以及研究植物的发育进化都起到关键作用。
叶绿体的类囊体膜复合物是疏水性大分子膜蛋白,为分离这类疏水性强的大分子膜蛋白,有研究者[9]建立了蓝绿温和凝胶电泳系统(blue native polyacrylamide gel electrophoresis,BN-PAGE),利用该技术可真实反映复合物的大小、数量、组成及相对丰度等。BN-PAGE一方面依靠考马斯亮蓝G250对蛋白的结合,为蛋白质提供负电荷,利用电场力分离不同蛋白复合物;另一方面,可根据蛋白分子量的大小,利用分子筛效应进行分离,在研究叶绿体类囊体膜复合物的组成、生物发生中具有十分重要的作用[10]。目前BN-PAGE最常用于分析双子叶拟南芥和单子叶水稻类囊体膜组成,而关于玉米、油菜等的研究较少[11-12]。
本研究以双子叶 C3光合模式植物拟南芥和本氏烟草、单子叶C3光合模式作物水稻和C4光合玉米、双子叶 C4光合白花菜、C3-C4光合中间型Moricandiasuffruticos(MS)二倍体物种以及多倍体作物油菜、大豆和花生为材料,运用 BN-PAGE技术手段,从生化分子水平解析C3型与C4型、单子叶与双子叶、二倍体与多倍体不同类型作物光合蛋白复合物的差异,为改良光合作用提供理论支撑。
1 材料与方法
1.1 试验材料
选用拟南芥、本氏烟草、水稻、玉米、MS、白花菜、油菜、大豆和花生共9种植物为材料,分别编号1~9,拟南芥为Col-0生态型,水稻为籼稻。拟南芥、本氏烟草、MS、白花菜和油菜种子均由本课题组保存,其中拟南芥、烟草、MS和白花菜种植于中国农业科学院油料作物研究所温室,培养条件为温度22℃,光照强度约100μmol/(m2∙s),光周期为16h/8h(光/暗)。油菜也种植于该温室,培养条件为白天温度 24℃,夜间温度 22℃,光照强度约 200μmol/(m2∙s),光周期为 16h/8h(光/暗)。水稻、玉米、大豆和花生取自油料作物研究所试验田。为确保不同物种之间取样的一致性和结果的可比性,根据不同作物的生长发育周期,叶片取样时以3周龄的拟南芥为参照,分别对应烟草、水稻、玉米、MS、白花菜、大豆、油菜和花生第3周、8周、6周、4周、4周、3周、9周、4周的新生完全展开叶。
1.2 类囊体膜蛋白样品的制备
按照Järvi等[13]的方法稍做改动进行9个植物类囊体蛋白的提取制备。将材料从-80℃冰箱中取出,在研钵中倒入少许液氮研磨至粉末状后,加入低渗裂解液[50mmol/L Hepes-KOH(pH 7.5),330mmol/L山梨醇,2mmol/L EDTA,1mmol/L MgCl2,5mmol/L抗坏血酸,0.05% BSA,10mmol/L氟化钠],转至冰上继续研磨至浆液均匀,3层Miracloth滤布过滤,于4℃、8000g离心10min得到类囊体膜沉淀,用shock缓冲液[50mmol/L Hepes-KOH(pH 7.5),5mmol/L 山梨醇,5mmol/L MgCl2,10mmol/L氟化钠]洗涤 1次,用储存缓冲液[50mmol/L Hepes-KOH(pH 7.5),100mmol/L山梨醇,10mmol/L MgCl2,10mmol/L氟化钠]重悬洗涤1次。再次加入100μL储存缓冲液并按Porra等[14]的方法进行叶绿素含量定量。用适量预冷样品缓冲液[25mmol/L BisTris-HCl(pH 7.0),20%(W/V)甘油,10mmol/L氟化钠]将叶绿素浓度调整为1.0mg/mL,加入等体积含2%(W/V)去垢剂DM的样品缓冲液,轻轻用手指弹匀至悬浊液慢慢变为澄清,黑暗处冰浴增溶10min,4℃、18 000g离心20min后吸出上清液即为类囊体膜蛋白溶液。加入1/10体积上样缓冲液[100mmol/L BisTris-HCl(pH 7.0),2.5%考马斯亮蓝G250,500mmol/L 6-氨基己酸,30%甘油],然后上样电泳。
1.3 一向BN PAGE
按照 Liu等[15-16]的方法进行一向 BN PAGE,并稍做改动。根据用户手册,采用XCell SureLock mini-cell和Native PAGETM Novex 4-16% Bis-Tris mini gel进行蓝绿温和胶电泳。电泳中先使用含0.01%考马斯亮蓝 G250的阴极缓冲液[50mmol/L Tricine,15mmol/L BisTris-HCl(pH 7.0)]电泳至分离胶 1/3处时更换为含 0.001% G250的阴极缓冲液,阳极缓冲液始终为50mmol/L BisTris-HCl(pH 7.0)。为了保证蛋白复合物的完整性,于 4℃冰箱中进行电泳以保证低温环境。先25V电泳2h,随后每隔0.5h电压升高25V,直至电压升至150V,整个电泳过程约5h。
1.4 二向SDS-PAGE
灌制 10cm×10cm、厚度 1.5mm 的 12% SDSPAGE凝胶。一向BN-PAGE结束后,立即切下进行二向电泳,放入6mL平衡液[100mmol/L Tris HCl(pH 6.8),8mol/L 尿素,5% SDS,5% β-巯基乙醇,20%甘油]中,75℃水浴20min,在室温摇床上平衡20min。将处理后的胶条推入胶板紧贴分离胶表面,用10mA/板的恒流进行电泳。电泳结束后,将凝胶放入染色液,摇床上室温缓慢摇晃2h;清水冲洗干净,加脱色液室温脱色至背景干净即可。
1.5 免疫印迹
将分离的蛋白质电转移到 PVDF膜上,5%脱脂奶粉室温封闭1h,加入对应的一抗,于4℃下过夜结合,再和带有 HRP标签的羊抗兔抗体(1︰15000)反应 30min后,加入 ECL室温黑暗孵育5min后显影拍照并保存。除二抗(上海碧云天生物技术有限公司,货号A0208)外,所有抗体均购自瑞士Agrisera公司。
2 结果与分析
2.1 蓝绿温和胶电泳技术制备植物类囊体膜复合物方法的优化
采用4种不同浓度的非离子型去垢剂十二烷基-β-D-麦芽糖苷(n-Dodecyl-β-D-Maltopyranoside,DM)增溶拟南芥类囊体膜蛋白,发现DM终浓度为1%时类囊体膜增溶效果更好(图1a和b)。通过蓝绿温和胶电泳,有10条清晰的条带被分离出来。通过文献查阅和经验分析,条带Ⅰ为NDH-PSⅠ,Ⅱ为 PSⅡ的超级复合物(PSⅡSC),Ⅲ为 PSⅡ的二聚体(PSⅡD)和PSⅠ的单体(PSⅠM),Ⅳ为FoF1-ATPase,Ⅴ为 PSI核心亚基(PSⅠcore),Ⅵ为ATPase/Cytb6f复合物,Ⅶ为CP43缺失的PSⅡ核心复合物,Ⅷ为LHCⅡ的三聚体(LHCⅡT),Ⅸ为LHCⅡ的单体(LHCⅡD),而条带Ⅹ是未参与复合物形成的游离蛋白[17-18]。
与1% DM相比,0.5% DM增溶类囊体膜蛋白复合物条带模糊,说明增溶剂含量太少,不足以达到充分增溶效果;而2% DM的条带Ⅰ和Ⅲ明显变细,说明增溶剂浓度太高,引起复合物的降解。一向电泳染色后,分别用12%和15% SDS-PAGE进行二向电泳并染色。复合物得到较好分离,共得到52个蛋白点(图1c)。经过对比发现,15%凝胶分离时,蛋白点主要集中在凝胶上部,而12%分离胶蛋白点分布较为均匀,因此12%凝胶浓度分离效果优于15%。
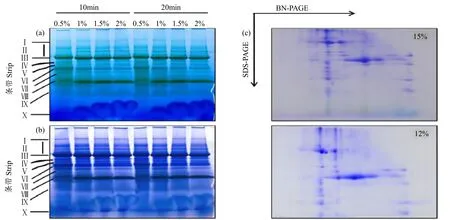
图1 BN/SDS-PAGE电泳分析模式植物拟南芥类囊体膜蛋白复合物Fig.1 BN/SDS -PAGE analysis of thylakoid membrane protein complexes in the model plant Arabidopsis
2.2 一向电泳比较分析不同作物类囊体膜复合物的差异特征
采用优化后的BN-PAGE体系提取9种植物的类囊体膜复合物,并进行BN-PAGE一向电泳。与烟草和水稻等C3作物相比,玉米和白花菜等C4作物PSⅡSC分离出更多条带,且条带更清晰,这与C4植物光能转化效率高的事实相一致(图2a)。而烟草和水稻中 PSⅡD含量高于玉米和白花菜。同样是 C3作物,与单子叶水稻相比,双子叶烟草的PSⅡSC分离出更多条带;而玉米和白花菜都属于C4型光合作用植物,与单子叶作物玉米相比,白花菜除了分离出较多的PSⅡSC,在PSⅡSC上游还存在几条蛋白条带,根据文献[12]和经验推测是PSⅠ-NDH复合物。
植物多倍体现象十分常见,多倍体植物具有细胞体积大、叶片宽厚、抗逆性和适应性强等特点[19]。但是通过一向电泳分析,多倍体作物油菜、大豆和花生的光合复合物并不高于二倍体作物烟草和水稻。拟南芥和MS的 PSⅡcore远高于其他作物。ATPase和Cytb6f在水稻中最多,玉米和白花菜中其次。此外,在烟草、水稻和玉米中都分离到PSⅡSC,拟南芥PSⅡSC的条带几乎不可见(图2a和b),可能是因为拟南芥的光合复合物对温度等外界因素更敏感,在提取和电泳过程中降解严重,所以在相同条件下拟南芥光合复合物几乎不可见[12]。

图2 BN-PAGE电泳分析不同作物类囊体膜蛋白复合物Fig.2 BN-PAGE analysis of thylakoid membranes in different crops
2.3 一向免疫印迹比较分析不同作物类囊体膜复合物的差异特征
为了进一步确认类囊膜复合物的丰度和位置,并比较不同作物类囊体膜差异性,用4大复合物的标志性抗体进行一向免疫印迹试验[20]。结果(图3)表明,不同作物类囊体膜复合物的蛋白含量具有很大差别。首先,C4作物玉米和白花菜中D1蛋白在PSⅡSC中含量高于C3作物水稻和烟草(图3a),而 PSⅡcore中水稻和烟草又高于玉米和白花菜。但是MS作为一种C3―C4中间型植物,无论是与C3还是C4植物相比,PSⅡSC中D1蛋白含量都显著降低。其次,在单子叶植物和双子叶植物中,单子叶作物水稻和玉米中 PSⅡD分别低于双子叶植物烟草和白花菜。同时发现多倍体作物油菜、大豆和花生PSⅡD含量高于烟草等二倍体作物。
Cytb6f在电子传递、调节光合电子传递链的电子速度与流向、PSⅡ与 PSⅠ之间的状态转移等方面具有十分重要的作用[21]。一向免疫印迹试验(图3b)表明,水稻中Cytb6f复合物最多,油菜和MS中含量较少。与C3作物水稻相比,玉米中Cytb6f含量下降;然而白花菜中 Cytb6f却高于烟草。同时,还发现同样是C4植物,白花菜中Cytb6f含量高于玉米,而同样是C3植物,单子叶水稻Cytb6f含量高于双子叶作物烟草。多倍体作物大豆和花生的Cytb6f复合物表达量高于拟南芥和烟草。在多倍体油料作物油菜、大豆和花生中,油菜中Cytb6f含量明显低于大豆和花生。
叶绿体自身基因psaa编码的光反应中心蛋白PsaA作为原初反应中心在PSⅠ中发挥重要作用。免疫印迹结果(图3c)表明PsaA在玉米中表达量显著高于其他作物。同时发现多倍体作物大豆和花生中PsaA含量并不高于二倍体,意味着多倍体和二倍体之间PSⅠ丰度可能并不具有显著性差异。有意思的是,与大豆和花生相比,同样是多倍体油料作物油菜中PsaA含量较低。与C3双子叶作物烟草相比,C4双子叶作物白花菜中PSⅠ核心亚基PsaA在PSⅠM含量更高,但是在PSⅠcore中则较低。
叶绿体 ATP合酶最终使光合生物实现了光能向化学能的转化[22]。图 3d表明不同作物 CF1β(AtpB)的差异性表现在2个方面:一是ATPase复合体的存在形式;二是ATPase复合体的丰度。一方面玉米和水稻等单子叶ATPase复合体主要以F1―ATPase复合物形式存在,而拟南芥,烟草和白花菜等双子叶植物主要以FoF1―ATPase形式存在;另一方面,单子叶作物水稻和玉米中ATPase复合体丰度比拟南芥,烟草、白花菜等双子叶植物高,这可能是单子叶和双子叶作物之间的差异性引起的。
2.4 免疫印迹分析类囊体膜四大复合物及其组成亚基
一向免疫印迹进一步确认了类囊膜复合物丰度和位置的差异性,但是不同作物对应亚基总含量是否有差别尚不清楚。因此,用常规免疫印迹分析各作物中四大复合物总含量。作为 PSⅡ中周转最快的蛋白,D1含量和PSⅡ含量成正相关关系。结果(图4a)表明,水稻中D1含量最高,说明水稻中PSⅡ含量最高。免疫印迹结果表明水稻中Cytb6f复合体最多,这与一向免疫印迹结果类似(图4b)。还发现同样是C4植物,白花菜中Cytb6f含量高于玉米,而同样是C3植物,单子叶植物水稻Cytb6f含量高于双子叶植物烟草。多倍体作物大豆和花生的 Cytb6f复合物表达量高于拟南芥和烟草。在多倍体油料作物油菜、大豆和花生中,油菜中Cytb6f含量明显低于大豆和花生。有趣的是,不同作物之间 Cytb6f复合物的核心亚基 Cyt f分子量差异很大。同时,玉米中PsaA表达量显著高于其他8种作物,并且C4作物白花菜中PsaA含量高于C3作物烟草(图4c)。同时,多倍体作物油菜、大豆和花生中PsaA总含量与二倍体植物拟南芥和烟草相当。ATP酶复合物核心亚基CF1β在单子叶水稻和玉米中表达量高于双子叶作物烟草和白花菜,这点与一向BN-PAGE免疫印迹结果类似(图4d)。
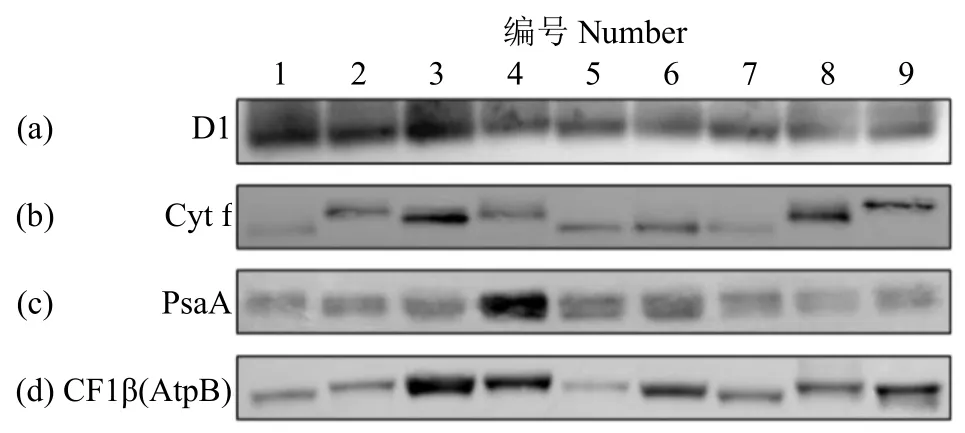
图4 免疫印迹分析不同作物光合蛋白Fig.4 Immunoblotting analysis of photosynthetic proteins in different crops
2.5 二向SDS-PAGE分离各复合物亚基
为了进一步验证类囊体膜的哪种复合物含量在C3向C4、双子叶向单子叶进化中发生更显著的变化,9种作物类囊体膜复合物经过BN-PAGE分离后,用二向SDS-PAGE变性电泳继续分离复合物的组分[15]。每个蛋白亚基按分子量大小在胶内分开,经考马斯亮蓝染色后得到清晰的蛋白点(图5)。其中在 SDS-PAGE凝胶上拟南芥共分离得到蛋白点33个、烟草43个、水稻40个、玉米45个、MS 30个、白花菜45个、油菜30个、大豆32个、花生32个。
二向考染得到了与一向免疫印迹类似的结果(图5)。首先,C4植物玉米和白花菜的D1分别表达量高于水稻和烟草。然而MS作为C3―C4中间型植物,无论是与C3植物相比,还是与C4植物相比,D1蛋白含量都明显降低。其次,双子叶烟草与单子叶水稻中D1表达量相当,白花菜比玉米中D1蛋白含量高。同时发现油菜、大豆和花生等多倍体植物中D1高于拟南芥和烟草等二倍体植物。并且二向电泳染色也发现玉米中PsaA含量比较高。考染结果还表明,单子叶植物水稻和玉米ATPase含量高于双子叶植物拟南芥和烟草等。

图5 二向BN/SDS PAGE分析不同作物类囊体膜复合物组成亚基Fig.5 Analysis of thylakoid membrane components in different crops by 2D BN/SDS PAGE
3 讨论
蓝绿温和胶电泳系统是一种非常有效的可以根据分子质量的不同而将完整的蛋白质复合物区分开的可靠方法[23]。不仅可以通过一向染色和免疫印迹试验进一步确定复合物的位置和丰度,还可以在变性条件下进一步通过 SDS-PAGE电泳进行二向电泳,分析复合物中单个亚基丰度。本研究以模式植物拟南芥为材料对BN-PAGE进行优化,基于研究结果,认为该体系同样适用于其他作物。在拟南芥中通过使用不同浓度梯度非离子型膜蛋白增溶剂DM结合不同增溶时间梯度,发现当DM浓度1%、增溶时间10min时效果相对最佳,具体表现为不同分子量大小的光合复合体条带分离完全且各条带均清晰可见,特别是可以分析完整的大分子量超级复合体和二聚体(图1a和b)。在使用这一优化的技术体系对水稻、玉米和大豆等其他不同作物进行BN-PAGE时可以分离出与模式植物拟南芥同样的各复合物条带,达到非常相似的预期效果(图2),说明通过控制相同的试验条件来分析差异的优化技术方法对于研究不同植物的光合性状具有一定的通用性。但仍有改进空间,如二向蛋白电泳还可增加蛋白质上样量,使用 SDS-Urea-PAGE凝胶等改进分辨率,并且经二向SDS-PAGE电泳后,还可以进行质谱鉴定。
本研究分析和解释不同类别植物间光合蛋白复合物是否存在差异性,选取9种代表性植物,借助蓝绿温和胶技术手段,从生化分子水平解析C3型与C4型、单子叶与双子叶、二倍体与多倍体不同作物其光合蛋白复合物的差异性,为定向改良光合作用提供理论参考。结果表明,与C3、双子叶和二倍体作物相比,C4、单子叶和多倍体作物中PSⅡ核心蛋白D1在超级复合物中分别表现出高丰度,也就是超级复合物含量比较高(图3a)。光能的捕获和传递通过光合复合物进行,该结果可能在一定程度解释了C4、单子叶和多倍体作物光合效率比较高的原因。结果还表明,虽然不同作物中Cytb6f含量不具有明显规律性,但是差异性很大,暗示Cytb6f复合物也是光合作用改良靶点。并且已有研究[24]表明,过表达CytC6基因可促进电子传输进而提高大田中烟草生物量。免疫印迹和考染结果都表明单子叶植物水稻和玉米ATPase含量高于双子叶植物拟南芥和烟草等。这或许是单子叶植物生物量高于双子叶植物的原因。在光能转化过程中,类囊体膜电子传递为碳同化提供还原力,因此,类囊体膜复合物上电子传递速率将直接影响碳同化进而影响光合作用。也就是说,只要过表达的基因可以促进电子传输就可以提高田间作物生物量。本研究主要围绕C3型与C4型、单子叶与双子叶以及二倍体与多倍体作物之间光合特性的比较展开,没有进行不同作物及其亚种之间光合性状比较,然而许多作物的不同亚种,如水稻的籼稻和粳稻在形态特征和农艺性状等方面有所不同,探究籼稻和粳稻的光合性状差异对于通过遗传改良选育高光效、高产、高抗水稻具有重要的潜在的应用价值,对于其他作物不同亚种的改良同样具有借鉴意义,后续需要对其他重要农作物及其亚种之间的光合性状差异进行进一步研究,比较分析不同作物间光合蛋白复合物差异为作物光合性状的改良和光合效率的提高提供参考。
4 结论
运用蓝绿温和胶(BN-PAGE)技术手段证明了不同类别作物光合复合物及其相应代表性亚基在蛋白丰度和组成形式等方面具有显著不同。与C3、双子叶、二倍体作物相比,C4、单子叶和多倍体作物中PSⅡ核心蛋白D1在复合物中的含量均升高;水稻中Cytb6f含量最高,不同作物之间Cytb6f核心亚基Cyt f丰度差异较大;玉米中PSⅠ核心亚基PsaA的含量显著高于其他作物,且C4作物高于C3作物;单子叶植物中ATP酶复合物核心亚基CF1β含量显著高于双子叶植物。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17
原子与分子物理学报(2021年2期)2021-03-29
中成药(2018年7期)2018-08-04
生命科学研究(2018年1期)2018-05-29
中成药(2018年3期)2018-05-07
中成药(2018年1期)2018-02-02
作文周刊·小学三年级版(2017年15期)2017-06-22
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06
