
水稻OsFTL12和OsFTL13基因双突变体的创建及新型编辑类型的鉴定
2021-10-22 02:03周诗晨孙丽娜兰志鹏王馨翊杨晓颖梁闪闪栾维江
天津师范大学学报(自然科学版) 2021年4期
郑 瑞,周诗晨,孙丽娜,田 瑶,王 鑫,兰志鹏,王馨翊,杨晓颖,梁闪闪,栾维江
(1.天津师范大学生命科学学院,天津300387;2.天津师范大学天津市动植物抗性重点实验室,天津300387)
水稻从营养生长到生殖生长转变是水稻生命活动中的关键转折点,这个转变的时间一般称为开花时间(flowering time),开花过程是由植物自身基因和环境(光照和温度等)相互作用决定的[1].影响水稻开花时间的主要环境因素是光周期,植物对光周期能够做出规律性应答.植株体内有着复杂的光周期调控通路,其中各基因相互作用,共同决定水稻的开花时间.成花素基因(FLOWERING LOCUS T,FT)是植物光周期调控开花途径中的一个关键基因,在多种植物中能够促进开花[2].水稻同源的FT-like基因家族包含13个成员,即OsFTL1~OsFTL13,其中,OsFTL2(Heading date 3a,Hd3a)和OsFTL3(RICE FLOWERING LOCUS T1,RFT1)的研究较为深入[3-4].Hd3a是拟南芥FT的同源基因,其作用机理已被揭示清楚[3].Hd3a蛋白在水稻叶片中合成,经过茎韧皮部转运到达茎顶端分生组织(shoot apical meristem,SAM),2分子的Hd3a蛋白先在SAM细胞的细胞质中与2分子的14-3-3受体结合,然后进入细胞核与2分子的OsFD1蛋白结合,形成开花激活复合体(florigen activation complex,FAC).之后,FAC结合到OsMADS15基因启动子序列上调控基因表达,进而促进水稻开花[5].成花素蛋白属于磷脂酰乙醇胺(phosphatidylethanolamine-binding protein,PEBP)蛋白家族,在进化上比较保守[6].本实验室前期构建了OsFTL12的过量表达载体,通过农杆菌介导的遗传转化法获得了转基因植株,田间观察发现OsFTL12过量表达转基因植株呈现晚抽穗表型[7].将FT-like基因家族成员进行氨基酸序列比对,发现在系统发育树上,OsFTL12和OsFTL13处于同一分支,二者的氨基酸序列同一性高达66.49%,暗示二者可能具有相似的生物学功能.为了进一步研究这2个基因的功能,本研究创建了OsFTL12和OsFTL13基因双突变体.
CRISPR-Cas9(clustered regularly interspaced short palindromic repeats,CRISPR-associated 9)技术是近些年发展起来的新一代基因编辑技术.该技术发挥功能的原理:CRISPR簇经过转录加工形成成熟的crRNA,与tracrRNA和Cas形成复合体,结合匹配到靶DNA序列并切割目标DNA链,生物体在进行DNA链断裂修复时产生碱基插入或缺失,这种插入或缺失主要是被编辑的生物体内源DNA序列的插入或缺失[8].与锌指核酸内切酶(zinc finger endonuclease,ZFN)和类转录因子效应物核酸酶(transcription activator-like effector nuclease,TALEN)相比,CRISPR系统具有成本低、效率高、操作简单等优点[9],因此得到迅速发展和应用,成为研究基因功能的一种工具.
本研究采用CRISPR-Cas9载体系统,将OsFTL12和OsFTL132个基因的靶点序列融合到CRISPR-Cas9载体中,通过农杆菌介导的水稻遗传转化法,获得转基因植株.对转基因植株进行鉴定分析,获得OsFTL12和OsFTL13基因双突变体,为研究OsFTL12和OsFTL13基因功能奠定基础.
1 材料与方法
1.1 实验材料
以粳稻品种中花11(Oryza sativaL.ssp.Japonicacv.Zhonghua11)为背景材料,获得CRISPR-Cas9编辑材料.
1.2 CRISPR-Cas9载体构建
首先在OsFTL12和OsFTL13基因的外显子上设计用于CRISPR-Cas9编辑的靶点.以含有启动子和gRNA的质粒作为模板,分别加入靶点引物,通过PCR获得各个融合有靶点的表达盒(启动子-靶点-sgRNA).PCR程序为:94℃预变性5 min;94℃变性30 s,62℃退火30 s,72℃延伸30 s,30个循环;72℃充分延伸7 min.将所有靶点表达盒和CRISPR-Cas9载体用BsaI酶切,利用T4-DNA连接酶在16℃水浴条件下连接过夜,连接产物经过热激转化,挑取阳性单克隆摇菌后,提取质粒进行酶切鉴定,获得同时连接3个靶点的重组载体,重组载体测序确认后备用.所设计靶点序列:FTL12-TG1为5′-GAGGCCTGTGTTTAATGGCA-3′;FTL12-TG2为5′-GCAGGTATTCCCTCAAGGTT-3′;FTL13-TG为5′-GCGTGTGATCGGAGATGTCC-3′.
1.3 转基因植株的获取及分子检测
将正确的CRISPR-Cas9重组载体导入农杆菌中转化水稻品种中花11,产生CRISPR-Cas9编辑的突变体.待植株生长健壮,收集各植株叶片,使用CTAB法提取DNA,以此作为模板,用载体中特异的潮霉素基因(Hyg)引物进行PCR扩增,PCR程序同1.2中的程序,检测重组载体是否导入水稻中.Hyg引物序列:HygF为5′-GGAGCATATACGCCCGGAGT-3′;HygR为5′-GTTTATCGGCACTTTGCATCG-3′.
1.4 突变体植株编辑类型鉴定
为了明确编辑突变体发生了何种类型的突变,分别在OsFTL12和OsFTL13基因的靶点上下游设计检测引物,以单株DNA为模板进行PCR扩增,将PCR产物进行琼脂糖凝胶电泳检测后,送至生工生物工程(上海)有限公司进行测序,从而确认各编辑突变体发生的编辑类型.检测引物序列:FTL12-TestF为5′-GCCTGTTAGCATTTGTAGAG-3′;FTL12-TestR为5′-GTCACCATCCTGTCATATTG-3′;FTL13-TestF为5′-GTCCCAACACAACACAAACC-3′;FTL13-TestR为5′-AAGTGGGTTTAGCGGGTTAG-3′.同时,在FTL12-TG1靶点上下游设计一条新的检测引物(野生型中目标片段大小为206 bp),用于检测发生了较大片段缺失或者插入的编辑类型.引物序列:FTL12-XF为5′-TAGTTGGAGATGTGTTAGAC-3′;FTL12-XR为5′-GACTCTTTTGGATCGGGTAT-3′.用该引物进行PCR扩增后,产物用3%琼脂糖凝胶电泳进行检测,直观检测在编辑突变体中发生的缺失或插入.
2 结果与分析
2.1 靶点选择及引物设计
借助网站(http://skl.scau.edu.cn/home/),在目标基因编码序列中依照GC含量50%~70%、长度20 nt、含有NGG(protospacer adjacent motif,PAM)识别序列的原则筛选靶点.OsFTL12和OsFTL13均含有4个外显子和3个内含子,如图1所示,二者的编码序列总长分别为522 bp和558 bp.本研究在OsFTL12基因上选择了2个靶点,第1个靶点位于第1个外显子,命名为FTL12-TG1,第2个靶点位于第2个外显子,命名为FTL12-TG2;在OsFTL13基因的第1个外显子上选择了1个靶点,命名为FTL13-TG.将3个靶点序列分别进行Blast序列比对,结果显示各靶点均特异.
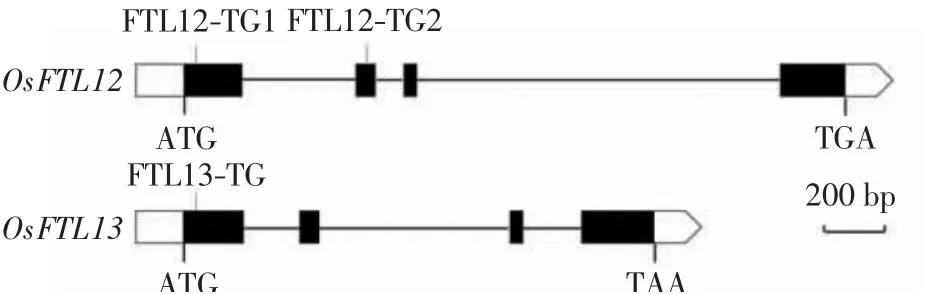
图1 OsFTL12和OsFTL13基因结构及靶点位置Fig.1 Gene structure and target sites position of OsFTL12 and OsFTL13
2.2 CRISPR-Cas9载体构建
将靶点同启动子和gRNA序列融合,利用PCR扩增分别得到3个靶点表达盒,产物经过琼脂糖凝胶电泳验证,目标片段大小正确,如图2(a)所示.将3个靶点表达盒同时连入CRISPR-Cas9空载体获得重组载体,重组载体进行酶切鉴定,结果显示切出了预期大小的目的片段,如图2(b)所示,表明含有3个靶点的sgRNA已成功连入了目的载体中.将重组载体送往生工生物工程(上海)有限公司测序,结果显示正确,重组载体可用于后续实验.
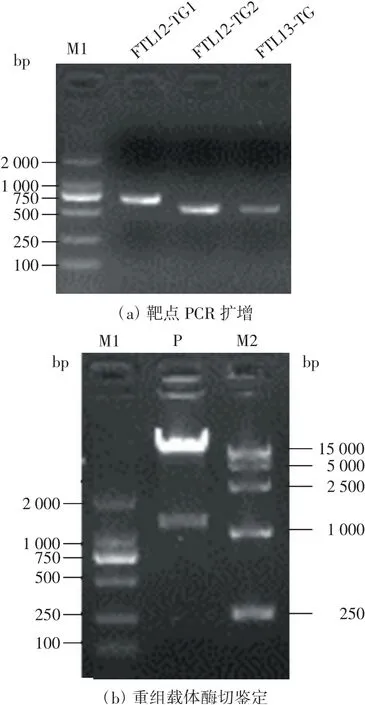
注:M1为DL2000 DNA Maker;M2为DL15000 DNA Maker;P为重组载体.
2.3 转基因植株的获取及分子检测
将验证正确的CRISPR-Cas9重组载体通过电击转化进农杆菌感受态(EHA105)中,通过农杆菌介导的遗传转化方法将CRISPR-Cas9重组载体导入水稻品种中花11中,共得到9株转基因当代(T0)植株.用载体中特异的标记基因引物对获得的转基因植株进行PCR鉴定,结果表明有8株为转基因阳性植株,如图3所示.

图3 T0代转基因植株分子鉴定Fig.3 Identification of T0 generation of transgenic plants
2.4 突变体植株编辑类型分析
通过CRISPR-Cas9系统获得的突变体编辑类型多样,为了检测获得的8株转基因阳性植株是否发生了编辑,用OsFTL12和OsFTL13特异检测引物分别扩增出包含OsFTL12和OsFTL13靶点的基因片段,对产物进行测序,结果如图4所示.
总之,车载光伏远程控制空调是汽车制冷发展的一个重要方向,由于其在应用上的优点突出,在未来拥有着广阔的应用前景。


图4 T0代突变体的编辑类型分析Fig.4 Analysis of T0 generation of editing mutants
测序结果表明,8株转基因阳性植株中有3株双编辑突变体,命名为m1、m2和m3.其中,OsFTL12的突变均发生在FTL12-TG1靶点上,编辑类型有3种.m1是一个纯合突变体,在PAM前3 bp处发生1 bp缺失,如图4(a)和图4(h)所示.m2也是一个纯合突变体,在PAM前17 bp处发生19 bp缺失,经3%琼脂糖凝胶电泳检测发现,野生型中扩增出了206 bp的目的片段,而在突变体中扩增出了较小的187 bp的片段,说明该突变体的确缺失了19 bp,如图4(b)、图4(c)和图4(h)所示.m3是一个大片段插入的双等位突变体,OsFTL12基因靶点序列扩增产物经过琼脂糖凝胶电泳检测后发现有2个条带,将其分别进行回收测序,结果显示m3的一条同源染色体上在PAM前3 bp处发生1 bp缺失,该链用m3′表示;另一同源染色体上发生了376 bp的大片段插入,该链用m3″表示,如图4(d)、图4(e)、图4(f)和4(h)所示.将m3″的测序序列在NCBI(www.ncbi.nlm.nih.gov)上进行Blast比对,结果表明这个大片段来自于CRISPR-Cas9载体,具体发生的编辑类型为,在PAM前3 bp处缺失10 bp水稻OsFTL12基因序列,紧接着插入376 bp的载体部分序列.用BioXM2.6将这376 bp与部分CRISPR-Cas9载体序列进行比对,结果完全一致,如图4(j)所示.
3个突变体中,OsFTL13的编辑类型仅有一种,均在FTL13-TG靶点PAM前4 bp处纯合缺失1 bp,如图4(g)和图4(i)所示.
2.5 突变体的氨基酸序列比对
OsFTL12基因编码序列522 bp,编码的蛋白质含有173个氨基酸.OsFTL13基因编码序列558 bp,编码的蛋白质含有185个氨基酸.用BioXM 2.6软件将获得的3株突变体OsFTL12和OsFTL13序列进行翻译,结果显示,3个突变体OsFTL12和OsFTL13基因的翻译过程均因移码造成提前终止,最终生成不同程度截短多肽.m1、m2、m3′和m3″的OsFTL12编码的截短蛋白分别包含46、40和46和39个氨基酸,OsFTL13编码的截短蛋白均包含36个氨基酸,如图5所示.

图5 突变体的氨基酸序列比对Fig.5 Comparison of amino acid sequence in different mutants
2.6 双等位缺失插入突变体m3的遗传分析及突变体植株的抽穗期统计
将m1、m2和m3转基因当代植株自交产生的种子进行播种,得到OsFTL12和OsFTL13不同编辑类型的T1代植株.以T1代单株DNA为模板,用FTL12-XF和FTL12-XR引物进行靶点序列扩增.测序结果显示,m3的T1代植株靶点序列得到分离,种植的29个单株中,纯合10 bp缺失和大片段插入的有10株,即图6(a)中的2、4、5、7、8、11、14、18、21和29;纯合1 bp缺失的有6株,即图6(a)中的12、15、17、19、24和28;双等位编辑的13株,即图6(a)中的1、3、6、9、10、13、16、20、22、23、25、26和27.说明突变类型能稳定遗传,可以用于基因功能分析.
经过测序确定各单株的编辑类型,统计OsFTL12和OsFTL13发生编辑单株的抽穗期,同野生型进行比较,结果如图6(b)所示.由图6(b)可以看出,与野生型相比,m1和m2抽穗期未发生显著性变化,m3的抽穗期提前了1.7 d.
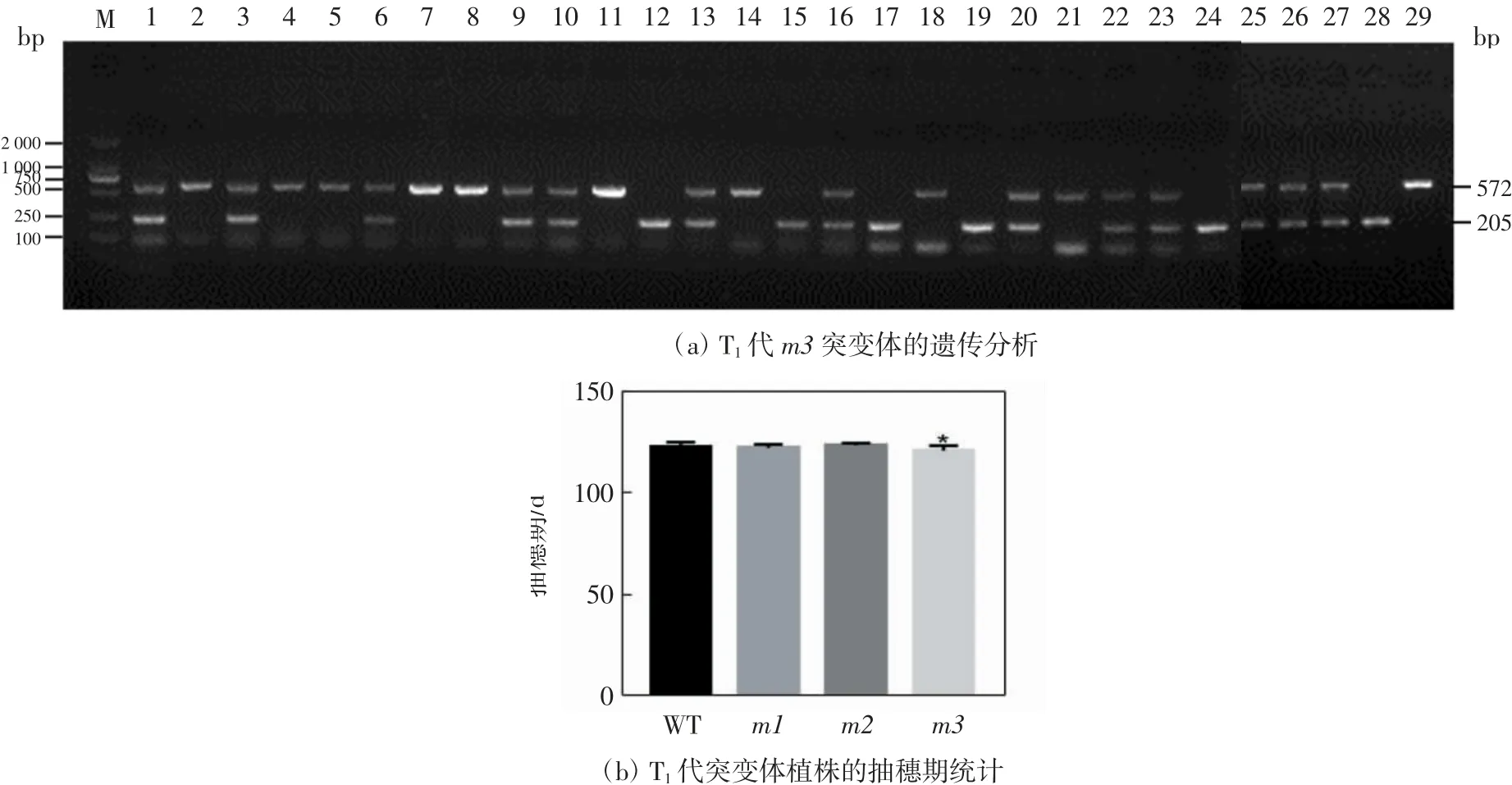
图6 T1代m3突变体的遗传分析及突变体植株的抽穗期统计Fig.6 Inheritance analysis of m3 mutants and statistics of heading date of mutants in T1 generation
3 讨论与结论
本研究利用CRISPR-Cas9编辑技术创建了水稻OsFTL12及OsFTL13基因的双突变体,并对突变体的编辑类型进行分析.产生的双突变体中,OsFTL12基因发生了3种类型的编辑,分别为纯合1 bp缺失、纯合19 bp缺失和双等位突变.尤其是在双等位突变体中发生了鲜见的CRISPR-Cas9载体序列插入的编辑类型.OsFTL13基因仅有一种类型的编辑,即1 bp缺失.这些双突变体的创建为全面研究水稻成花素家族基因功能奠定了材料基础,尤其新编辑类型的产生拓展了人们对CRISPR-Cas9编辑类型的认识.
虽然CRISPR-Cas9编辑技术简单有效,但在实际应用中存在较大风险,如脱靶(off-target)的可能[11].可以通过一些软件进行脱靶效应评估[12],也可以进行全基因组测序来确定脱靶位点.通过各种改良措施来降低脱靶率一直是研究热点,然而目前脱靶现象仍不可避免,这种脱靶的风险主要是由于CRISPR-Cas9编辑类型丰富复杂.通常CRISPR-Cas9编辑可能会产生所编辑基因的序列插入、缺失和替换[13],很少发生载体片段的插入类型.在本研究中,创建CRISPR-Cas9编辑突变体的过程中意外得到了一株插入CRISPR-Cas9载体片段(376 bp)的突变体,并且该突变体可以稳定遗传到后代,表明了CRISPR-Cas9编辑类型的复杂性及多样性.将靶点序列与插入的CRISPR-Cas9载体序列进行比对,发现二者没有同源性,说明新型插入突变体应该不是通过同源重组产生的.这也意味着这株特殊突变体很有可能是由错误切割导致的:CRISPR载体在进行基因组目标序列切割时,也切下了自身的一段序列,被切断的基因组在修复过程中,错误引入了这一大片段载体序列.事实上,靶序列可能在匹配目标基因组时发生错误结合,从而导致错误切割基因组.Zuccaro等[14]在尝试利用CRISPR-Cas9技术纠正人胚胎EYS位点突变导致的致盲性状的过程中,发现CRISPR-Cas9编辑导致频繁的染色体丢失.本研究中特殊突变体的获得也暗示了CRISPR-Cas9系统编辑的复杂性和不可控性,它不仅可能会切割与靶点序列完全匹配的基因序列,也会对匹配度不高的序列进行切割.这种不可控性严重限制了其在基因治疗中的应用和发展.
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
亚热带农业研究(2022年1期)2022-08-08
中国农学通报(2022年12期)2022-06-01
中老年保健(2021年3期)2021-12-03
中国种业(2021年11期)2021-11-25
农业科技通讯(2021年1期)2021-03-06
中国生殖健康(2020年7期)2020-12-10
中国农业科技导报(2020年3期)2020-03-15
山西农业科学(2020年2期)2020-02-29
