
毛竹叶片性状及其异速增长关系的海拔梯度效应
2021-10-29 13:24胡瑞财程建新蓝春宝陈双林郭子武
竹子学报 2021年1期
胡瑞财,徐 森,程建新,蓝春宝,陈双林,郭子武
(1.浙江省龙游县林业技术推广站,浙江 龙游324400;2.中国林业科学研究院亚热带林业研究所,浙江 杭州311400)
叶片性状是植物长期适应外界环境所表现出的适应性特征,与植物的生长对策和资源的获取、利用能力密切相关,能够反映植物适应环境变化所形成的生态适应策略[1],因此不同生境下同一植物或同一生境下不同植物的叶片性状通常差异较大。异速增长采用幂函数方程来描述植物构件特征间的相对变化关系,降低了采用构件特征绝对量或比例带来的波动性,是目前植物表型可塑性研究较为理想的方法[2]。植物构件间的异速增长关系主要受植物自身遗传性所决定,同时也受资源可用性和环境变化的影响,其差异反映了植物对环境资源的分配及利用状况[3-4]。海拔是综合性的生态因子,其改变通常会导致气象、土壤因子等发生剧烈变化,如海拔升高,光照增强、温度下降、空气湿度增加[5],土壤氮、磷含量降低,而碳含量增加等[6],进而形成生境条件的海拔特异性,致使植物性状产生明显的适应性调节。研究发现,随海拔升高,披针叶黄华(Thermopsis lanceolata)叶片表现出“小型化”的趋势,其通过降低叶面积和叶厚以适应高海拔的环境胁迫[7];而高海拔下山东银莲花(Anemone shikokiana)叶片厚度增加以减少水分散失[8]。可见,植物通过叶片形态塑性调节提高自身的生境适应和风险规避能力,且不同植物的生境适应策略存在明显差异,但不同海拔竹子叶片性状变化特征及其适应机制的研究较少[4],其相对生长关系适应性调节及其生态学意义亦亟待阐明,这对于明确竹子在与环境交互作用中形成的生存适应策略具有重要科学意义。
毛竹(Phyllostachys edulis)隶属禾本科竹亚科刚竹属,单轴散生型高大竹种,具有生长速度快,笋、材产量高,加工性能好等特征,是中国面积最大、分布最广、经济价值最高的竹种。我国现有毛竹林面积达443万hm2,分布于全国20余个省市区[9],且多生长于山地丘陵地区,资源垂直分布十分明显。目前海拔梯度上毛竹的研究仅局限于种群结构[10]、竹笋品质[11]、土壤理化性质[12]等方面,而海拔对毛竹叶片性状及其异速增长关系的影响尚不清楚,这极大地限制了对毛竹种群演替、扩张及稳定机制的环境阐释。为此本文以垂直分布明显、经营水平基本一致的3个海拔毛竹林为研究对象,测定了不同海拔1~3度立竹叶片形态指标,并分析其异速增长关系,试图探究海拔是否会对毛竹林立竹叶片性状产生明显影响,进而改变其相对增长关系。旨在明确试验区毛竹丰产培育适宜海拔,为毛竹林高效经营提供理论参考。
1 研究地概况
试验地位于浙江省龙游县沐尘乡(28°44′~29°50′N,119°06′~119°15′E)。该地区属亚热带季风性气候,四季分明,年平均降雨量1 620 mm,年平均气温17.4℃,年平均无霜期261 d,年平均相对湿度79%,年平均日照时数1 769 h,土壤主要为红壤。该地区毛竹资源丰富,现有毛竹林面积达0.5万hm2,且毛竹林资源垂直分布特征明显。试验地毛竹林大小年不明显,经营类型主要为毛竹材用林,主要经营措施为季节性伐竹、采笋、林下植被清理等,少有施肥。
2 材料与方法
2.1 样品采集与测定
2019年10 月,在试验区选取立地条件和经营水平基本一致的3个海拔[低海拔(230±10 m,L)、中海拔(570±10 m,M)和高海拔(870±10 m,H)]毛竹林,并在各海拔分别设置3块10 m×10 m样地,调查竹林结构(表1)。每块样地分别选取1度、2度和3度竹各6株,取竹冠上、中、下3个部位健康、完整的成熟叶片约200 g,置于冰盒中带回实验室,用于叶片性状分析。
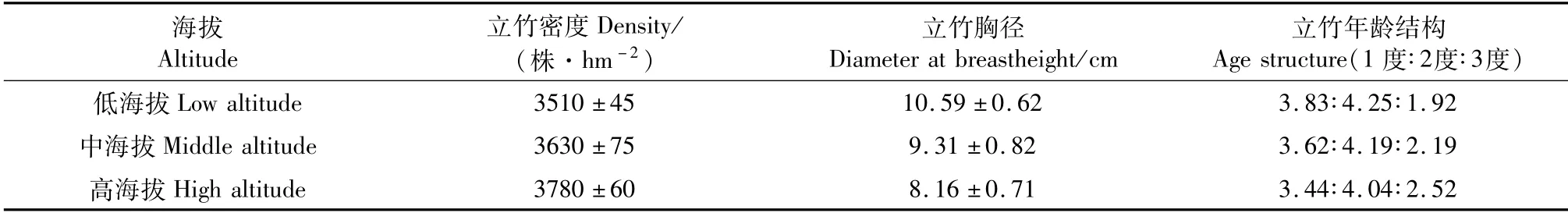
表1 试验毛竹林林分结构Tab.1 Summary of Phyllostachys edulis forest structure
将采集的样品洗净擦干后,每个处理每个重复随机选取15片完全伸展、无机械损伤的叶片,选用精度为0.01mm的游标卡尺在叶片中部测量叶片厚度,后采用万深LA-S植物图像分析系统测量叶长、叶宽和叶面积,计算长宽比,并以叶脉为对称轴测量叶片半叶宽度、半叶面积,计算叶宽和叶面积的波动不对称AF值。叶片形态指标测定后将叶片置于105℃杀青30 min,再65℃烘至恒重称其叶干重,并计算比叶面积(比叶面积=叶面积/叶干重)。
2.2 数据处理
试验数据在Excel 2010中进行数据处理和图表制作,采用SPSS 23.0进行单因素(One-way ANOVA)方差分析和邓肯(Duncan)多重比较(α=0.05),分析不同海拔1~3度毛竹叶片性状的差异;采用R 4.0.3统计软件的Smart软件包对不同海拔毛竹叶长、叶宽、叶面积进行标准主轴回归分析(Standardized Major Axis,SMA),异速增长方程为Y=a xb,经对数转化后为log y=loga+blogx,其中方程斜率b为异速增长指数,若|b|与1.0差异显著,表明x和y间呈异速增长关系,同时比较不同海拔斜率之间的差异性,若其无显著差异则计算共同斜率,并采用Wald检验分析不同海拔毛竹沿共同主轴是否发生明显位移。
叶片的波动不对称AF值计算方法如下:
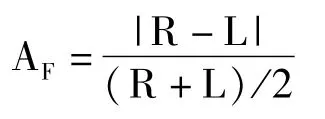
式中L为叶片左侧特征值(包括叶片左半宽和左半面积),R为叶片右侧特征值(包括叶片右半宽和右半面积)。
3 结果与分析
3.1 海拔对毛竹林立竹叶片形态性状的影响
由表2可知,随立竹年龄增加,同一海拔梯度毛竹林立竹叶长、叶宽、叶面积、叶厚和叶干重均呈升高趋势,且2、3度立竹间无明显差异,但均显著高于1度立竹。随海拔升高,同一年龄立竹叶长、叶宽、叶面积、叶厚和叶干重均呈升高趋势,其中低海拔立竹叶宽、叶厚和叶干重显著低于高海拔,而低海拔1、2度立竹叶长和叶面积显著低于高海拔,3度立竹间叶长和叶面积无明显差异;叶片长宽比不同海拔和不同年龄立竹间均无明显差异。可见,海拔对毛竹立竹叶片主要形态指标产生了明显的影响,但对叶片叶形指数(长宽比)影响较小,毛竹林叶片形态并无明显变化。
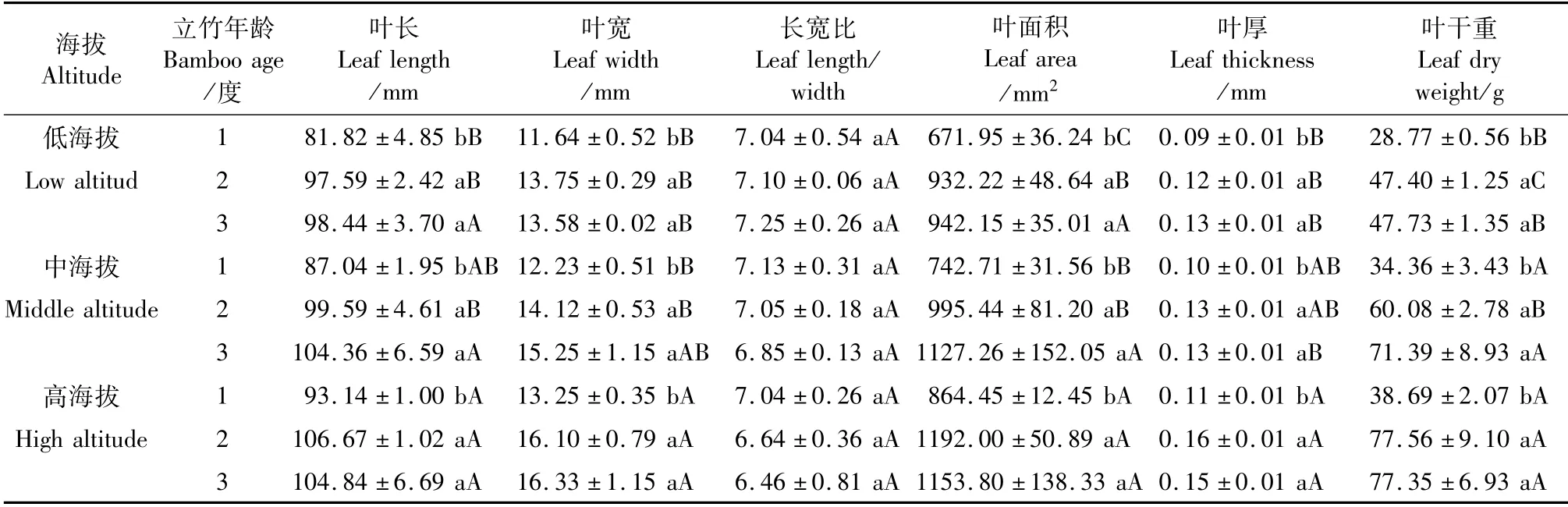
表2 不同海拔毛竹叶片性状Tab.2 Leaf morphological characters of Phyllostachys edulis at different altitudes
3.2 海拔对毛竹林立竹叶片比叶面积和波动性不对称的影响
由表3可知,随立竹年龄增加,同一海拔梯度毛竹林立竹比叶面积呈降低趋势,且2、3度立竹间无明显差异,但均显著低于1度立竹;高海拔叶宽和叶面积的AF值随立竹年龄增加均呈先升高后降低趋势,2度立竹AF值显著高于1度立竹,而与3度立竹间无明显差异。随海拔升高,同一年龄立竹比叶面积均呈降低趋势,其中低海拔2、3度立竹比叶面积显著高于中、高海拔,1度立竹间无明显差异;2度立竹叶宽AF值随海拔升高呈升高趋势,低海拔叶宽AF值显著低于高海拔;3度立竹叶面积AF值随海拔升高呈先降低后升高趋势,中海拔叶面积AF值显著低于低、高海拔。说明海拔对毛竹叶片比叶面积有明显影响,但对其叶片波动性不对称AF值影响较小。
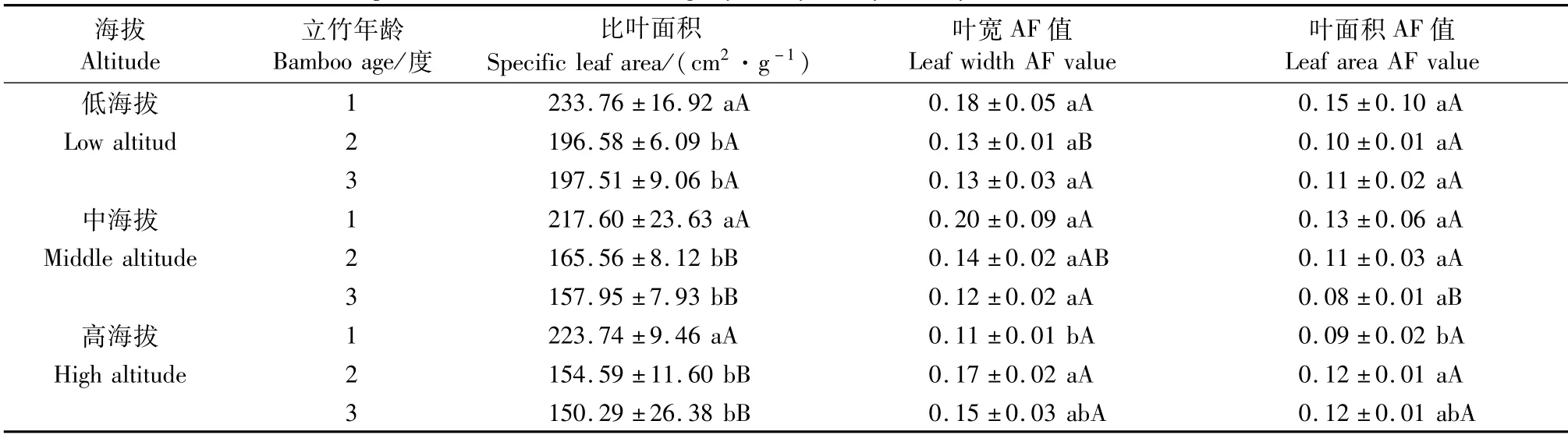
表3 不同海拔毛竹叶片比叶面积和波动性不对称性Tab.3 Specific leaf area and fluctuating asymmetry of Phyllostachys edulis leaf at different altitudes
3.3 海拔对毛竹叶长、叶宽、叶面积异速增长关系的影响
由图1可知,不同海拔毛竹林立竹叶长-叶面积、叶宽-叶面积和叶宽-叶长的回归斜率均显著大于或小于1.0,呈异速增长关系。随海拔升高,叶长-叶面积的斜率呈先降低后升高趋势,且不同海拔间差异显著;而叶宽-叶面积和叶宽-叶长异速增长指数不同海拔间并无显著差异,存在共同斜率,分别为0.571 2和1.106 8,截距分别为0.556 0、0.557 8、0.482 2和1.061 1、1.057 5、1.049 2,经Wald检验,低、中海拔叶宽-叶面积的截距无明显差异,均显著低于高海拔,而中、低海拔叶宽-叶长的截距无显著差异,且均显著低于高海拔,说明随海拔升高,毛竹叶宽-叶面积、叶宽-叶长截距均沿共同主轴发生明显负向位移,且以低海拔位移量最大。
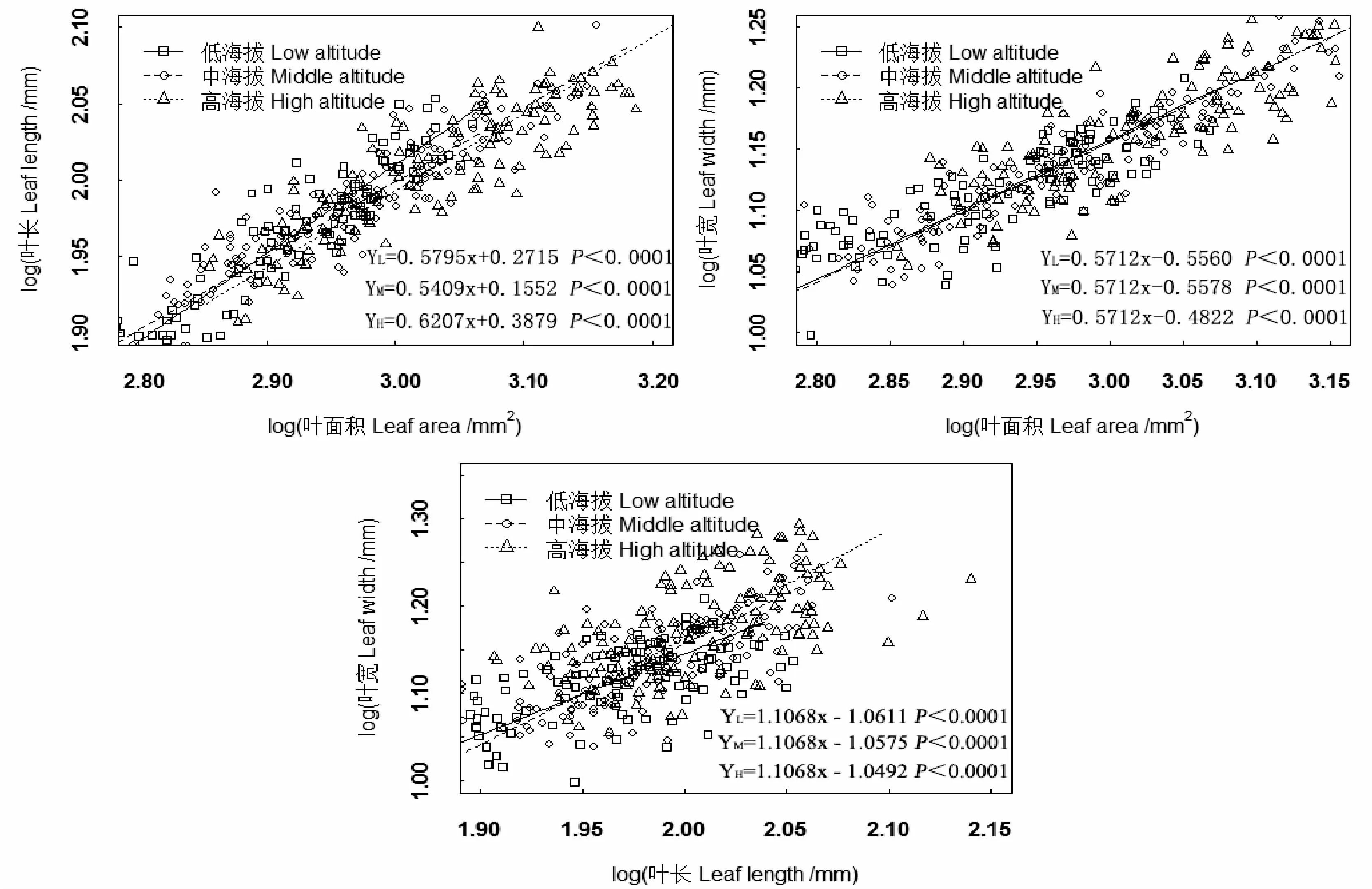
图1 不同海拔毛竹林立竹叶片叶长、叶宽与叶面积异速增长关系Fig.1 Allometric scaling between leaf length,leaf width and leaf area of Phyllostachys edulis at different altitudes
4 讨论
4.1 海拔对毛竹叶片性状的影响
叶片性状是植物与环境长期作用的结果,可以表征植物的生理生化特征[13-14],植物叶片性状在不同环境下表现出不同的生态对策,“性状-环境”之间的关系反应了植物生长和适应的最优“适应原则”[15-16]。本研究中,随立竹年龄的增加,同一海拔毛竹林立竹叶片叶长、叶宽、叶面积、叶厚和叶干重均呈升高趋势,说明立竹年龄对毛竹叶片形态特征有明显影响,随立竹年龄的增加,毛竹叶片呈现出变长、变宽、面积变大的趋势,但其叶形指数(长宽比)变化并不明显,即叶片形态相对稳定。随海拔升高,毛竹林立竹叶宽、叶厚和叶干重均呈升高趋势,且低、高海拔间差异显著。引起不同海拔叶片厚度明显变化的原因可能是水分和光照等的差异,海拔升高,降水量增加,而叶片厚度与植物的水分保存及利用密切相关[17],同时叶片厚度降低能提高植物在低海拔弱光下的光合作用能力[8,18]。叶面积大小直接影响植物对光的截取和对碳的同化能力,与植物的光合作用密切相关[19],随海拔升高毛竹叶面积呈升高趋势,且1、2度低海拔毛竹叶面积显著低于高海拔,这与辽东栎(Quercus liaotungensis)叶面积随海拔的变化规律基本一致[20],说明低海拔毛竹通过降低叶面积以减少水分散失,进而适应其干、热的环境条件。
比叶面积能够反映植物对资源的获取及利用能力,表征植物对生境的适应特征,通常比叶面积较高的植物具有较高的生产力[21]。本研究中,随海拔升高,同一年龄立竹比叶面积呈降低趋势,且低海拔2、3度立竹比叶面积显著高于中、高海拔,这说明低海拔毛竹林具有较高的生产力;而海拔升高,光照增强,温度下降,CO2分压降低,毛竹为维持自身的正常生长代谢,会将更多的能量和物质用于合成防御物质,因而比叶面积显著降低[22],这也与高海拔毛竹叶片厚度显著增加的研究结果一致。叶片波动性不对称反映了外界环境胁迫对叶片双边对称结构偏离的影响程度,是衡量外界环境压力大小或生物体适应能力的重要指标[23-24],海拔对叶片AF的影响主要通过资源和代谢物的分配来改变个体或群体的生理投资[25],低海拔2度立竹叶宽AF值显著低于高海拔,中海拔3度立竹叶面积AF值显著低于低、高海拔,而其他年龄立竹叶宽AF值和叶面积AF值不同海拔间无明显差异,说明海拔对毛竹叶片波动性不对称无明显影响,其双边对称结构较稳定,这与不同海拔毛竹叶片叶形指数相对稳定的研究结果一致,这说明毛竹具有较强的环境适应能力。
4.2 海拔对毛竹叶长、叶宽、叶面积异速增长关系的影响
异速增长是指某两个性状的相对生长速率出现明显差异的现象,虽其主要由遗传因素决定,但环境因子和栽培措施等的变化会引起叶片形态的可塑性变化,进而导致其异速增长关系的改变[26-27]。异速增长关系可以体现植物性状特点及其相互关系,同时也反映了植物不同性状对于环境资源变化所采取的生态策略[28]。本研究中,海拔对毛竹叶片异速增长关系有明显影响,叶长-叶面积、叶宽-叶面积和叶宽-叶长的异速增长指数显著大于或小于1.0,呈异速增长关系,这与Guo等[4]对不同海拔苦竹(Pleioblastusamarus)叶性因子的研究结果基本一致。随海拔升高,叶长-叶面积的异速增长指数呈升高趋势,且不同海拔间差异显著,而叶宽-叶面积和叶宽-叶长间则均存在共同斜率,且其截矩均表现为高海拔明显低高于低海拔,表明在给定叶面积下,高海拔具有较大的叶宽,这与随海拔升高,毛竹叶片呈现出变长、变宽的趋势的结果基本一致。异速增长关系的变化可以表征植物最高程度的适应性[3],毛竹叶长对海拔变化响应敏感,是可塑性较高的性状之一,说明海拔变化引起的温度、湿度和光照等的变化对叶长有较强的限制作用,同时也说明毛竹主要通过叶长的可塑性调节来应对海拔变化引起的环境异质性,以增强对环境资源的获取及利用能力,提高毛竹对异质环境下的适合度。
5 结论
综上可知,海拔对毛竹叶片性状及其异速增长关系产生了明显影响,毛竹可以通过其叶片性状的可塑性调节适应海拔变化引起的温度、光照、湿度和土壤状况等的综合变化,但不同年龄立竹对海拔的响应规律有所差异。随海拔升高,毛竹叶片长、宽和面积明显增加,叶片呈现变大、变厚的趋势,且叶长对海拔变化响应敏感;随海拔升高,毛竹比叶面积则呈显著下降趋势,即毛竹林生产潜力明显下降,但不同海拔毛竹叶片长宽比变化不大,即叶片形态相对稳定。综合分析表明,试验区中海拔具有相对较大的叶长、叶宽、叶面积和比叶面积,其生产潜力较高,是毛竹林培育的适宜海拔。
猜你喜欢
天津农林科技(2022年2期)2022-04-19
新疆农业科学(2020年1期)2020-02-14
亚热带植物科学(2019年2期)2019-07-22
江苏农业学报(2018年5期)2018-09-10
乡村科技(2018年8期)2018-06-27
湖南林业科技(2017年1期)2017-02-06
江苏农业科学(2016年10期)2017-02-05
