
小麦根系细胞学观察体系的构建和应用
2021-12-01 08:47赵佳男王鹏程贾晓雯蒋钰婕许盛宝王晓明
麦类作物学报 2021年9期
赵佳男,王鹏程,贾晓雯,蒋钰婕,许盛宝,王晓明
(西北农林科技大学 农学院,陕西杨凌 712100)
小麦(TriticumaestivumL.)是世界上最重要的粮食作物之一,其高产稳产对于保障国家粮食安全至关重要。植物根系是水肥吸收的主要器官,并参与体内物质合成及转化[1]。小麦是须根系作物,其根系由初生根、次生根(不定根)以及再生的各级分支构成。其中,初生根一般由种子萌发出的一条主胚根和另外4~6条次生胚根组成,其对小麦根系建成和后期生长发育具有重要作用[2]。主胚根为中间最先突破种皮的初生根[3],随后在其两侧依次长出次生胚根,次生胚根数主要与材料和种子大小相关[4]。
成熟小麦胚根的纵向结构可以划分为根冠、分生区、伸长区和成熟区四部分[5]。根冠位于根尖的最顶端,具有保护分生组织感知重力、水分和养分的功能[6]。分生区细胞具有旺盛的分裂能力,是根系干细胞分裂分化出各根系组织的区域[7]。分生区内部靠近根冠的部位有一团活动缓慢的原始组织,称为静止中心,具有维持干细胞状态和特性的能力[8-9]。根系向上紧靠分生组织的区域称为过渡区,过渡区的细胞被证明具有独特的细胞学和代谢特性,它们能够感知不同的环境因素和内源信号,进而调控根系生长[10]。伸长区细胞分裂能力减弱,开始快速伸长,成为根尖深入土层的重要推动力。成熟区位于伸长区之上,是根部吸收水分和养分的主要部位。
成熟根尖的横向结构具有典型的放射状特征,即由外到内分别为表皮、皮层、内皮层、中柱鞘及中柱[7]。表皮位于最外层主要起到保护分生组织的作用。皮层位于表皮之下,其细胞通透性强,溶质及水分可以自由通过[11]。内皮层的细胞壁加厚,径向壁木栓化形成凯氏带,使根的吸收具有选择性[12]。中柱包括韧皮部和木质部,前者从枝条向根运输代谢物,而后者则向枝条输送水分和溶质[13]。木质部大导管位于中柱的中间,周围分布着原生导管和韧皮部。与双子叶模式植物拟南芥的根系横向细胞组成相比,单子叶植物根系除了皮层数和细胞来源有差异之外,其他细胞组成基本相似[12]。
目前,小麦根系解剖结构观察的常用方法有徒手切片法和石蜡切片法。徒手切片技术操作简单,不需要仪器设备,且没有经过化学试剂处理,可保持小麦根尖细胞自然色泽和结构状态,多适用于观察根尖横切结构[14]。用于小麦根尖纵截面观察时,切片厚度不好控制,无法保证切口水平,容易导致组织结构破损。石蜡切片厚薄均匀,清晰无皱褶,并可对材料进行连续切片和观察,是小麦横切结构观察最常用的方法[15]。但是观察纵向细胞结构时,石蜡切片纵切难度较大,细胞破损严重,且很难保证不同切片在根系的同一层,从而阻碍不同材料和不同类别根系的比较分析。因此,目前小麦根系的横向细胞结构研究远多于纵向,但纵向细胞结构对于研究根系发育的细胞学基础至关重要,其清晰准确的观察是该领域目前急需解决的技术难题。显微观察方面,电子显微镜分辨率及精度较高,利用其可以清楚观察到小麦根尖表皮结构,但其成本高,无法分析三维结构,且不能观察根系内部组成[16]。小麦根系内部显微观察的另一个难点在于样品较厚,光线难以穿透。激光扫描共聚焦显微镜(laser scanning confocal microscope,LSCM)可以对样品进行断层扫描和成像,且样品处理时的透明步骤有效解决了样品较厚的问题,从而实现了灵活、直观、无损伤的样品观察,并可以对多个光学切片进行叠加,分析细胞的三维空间结构,广泛应用于模式植物拟南芥的根系细胞学研究[17]。
本研究基于LSCM,优化了制片流程,调整了观察参数,构建了可以快速、清晰地对同一纵向切面进行观察的技术体系,并利用该技术体系对5个小麦品种的胚根纵向细胞结构进行了连续观察和比较,以期为小麦根系的生物学研究提供重要的技术支撑。
1 材料与方法
1.1 试验材料
本试验选取了的小麦品种农大1108、小偃22、烟农19号、碧蚂4号和西农88为供试材料。观察设备为激光共聚焦扫描显微镜(Olympus公司生产,型号IX83-FV1200)。
1.2 小麦幼苗的培养试验
采用水培方式,于西北农林科技大学人工气候室进行。人工气候室的条件设置为25/20 ℃,14/10 h,5 000 lx,湿度50%。根系培养实验设置3次重复。
1.2.1 种子处理
选择饱满、大小一致、无病害的种子在室温下用蒸馏水浸种两天后,用消毒液(绿伞)(消毒液∶蒸馏水=3∶2)对种子进行表面消毒10 min,然后用蒸馏水冲洗3~5次。在灭菌的培养皿(直径15 cm)中铺两层滤纸,加入5 mL蒸馏水润湿,将消毒后的种子均匀地摆放在滤纸上,培养皿用保鲜膜封口,并在保鲜膜上戳洞以防止种子进行无氧呼吸,再缓慢加入15 mL蒸馏水,将摆好种子的培养皿置于4 ℃冰箱,春化 3 d,确保种子发芽一致。
1.2.2 移苗
挑选长势一致的发芽种子移至发芽盒(12 cm×12 cm),每个发芽盒中放12粒,共三盒。发芽盒用保鲜膜覆盖(保鲜膜上留通气和供幼苗叶片伸出的孔),之后转入人工气候室培养14 d,用于后期观察分析。
1.3 试验观察
挑选茁壮无损伤小麦主胚根,截取根尖部位0.5 cm用于分生区细胞学观察;在主胚根长1/2处截取1 cm的根段,用于成熟区细胞学观察。样品处理步骤如下:
(1)固定:取固定好的根尖材料放入加有4%戊二醛固定液的1.5 mL离心管中,真空泵抽真空三次,压强大于0.08 MPa,每次30 min。抽真空后,室温下在固定液中过夜。
(2)脱水:固定完成后,将固定液吸出,按酒精浓度50%、65%、75%、85%、95%、100%进行梯度脱水,每个浓度处理30 min。
(3)透明:脱水后材料置于苯甲酸苄酯∶苯甲醇为2∶1的配比溶液中4 h,以备制片观察。
(4)观察:利用LSCM进行材料解剖结构观察。LSCM来自Olympus公司,型号为IX83-FV1200。选用488 mm激发光,将焦距对焦到根尖中柱所在层面,能够清晰地观察到截面细胞组织,观察拍照。
2 结果与分析
2.1 小麦根尖细胞学观察体系构建
LSCM在拟南芥根系细胞组织研究中的方法和参数已经非常成熟[18]。但是,由于细胞结构组成的差异,运用该体系直接观察小麦根尖细胞结构时,细胞轮廓不清晰,观察效果不佳(图1A、图1B)。因此,本研究对脱水、透明和放大倍数进行了一系列调整,发现脱水时间从每梯度20 min延长至30 min、透明时间从1 h延长至6 h后,利用细胞自发荧光,在488 nm波长激发光下,用显微镜逐层扫描至中间层(根系中央后生木质部导管切面清晰可见),调节显微镜放大倍数到20倍,调整激发光强度,最终可以完整清晰观察到小麦根尖的纵向细胞结构(图1C、图1D)。农大1108、小偃22、烟农19号、碧蚂4号和西农88的主胚根长分别为23.01、26.02、24.00、25.47和24.64 cm,差异较大(图1E)。

A:流程优化前小麦根尖分生区纵切结构图;B:流程优化前根尖成熟区纵切结构图;C:流程优化后小麦根尖分生区纵切结构图;D:流程优化后根尖成熟区纵切结构图;E:五个材料主胚根长度统计分析。A:Longitudinal section of wheat root tip meristem zone before procedure optimization;B:Longitudinal section of maturation zone before procedure optimization;C:Longitudinal section of wheat root tip meristem zone after procedure optimization;D:Longitudinal section of maturation zone after procedure optimization;E:Statistical analysis of radicle length of five materials.图1 制片流程优化前后小麦根尖分生区和成熟区的纵切结构Fig.1 Longitudinal section of wheat root tip meristem zone and maturation zone with or without section producing procedure optimization
2.2 小麦根冠的细胞学特征
根冠呈帽状处于根尖的顶端,由细胞壁较薄的细胞组成[18]。运用LSCM观察发现,供试小麦根冠边缘的薄壁细胞大致呈椭圆形,根冠中央的薄壁细胞大小近乎等径,总体形状不规则,排列松散,而基部细胞体积较小,细胞核大,细胞排列紧密有序,与该处细胞的旺盛分裂相一致(图2A~图2E)。根冠与后方的分生区之间可见明显的穹窿形分界线,将根冠顶端细胞与穹窿形分界之间的距离表示为根冠长。农大1108、烟农19号、西农88、碧蚂4号和小偃22根冠长分别为220.28、212.08、194.57、178.90和160.47 μm。进一步观察发现,根冠细胞有序排列为10~13层,其中农大1108为13层,小偃22、烟农19号及碧蚂4号均为11层,西农88为10层。每层细胞的纵向平均长度从上到下逐渐下降,不同材料间同一层细胞的长度存在显著差异(图2F~图2H)。这说明细胞层数和每层细胞长度的不同共同引起不同材料间的根冠长度差异。
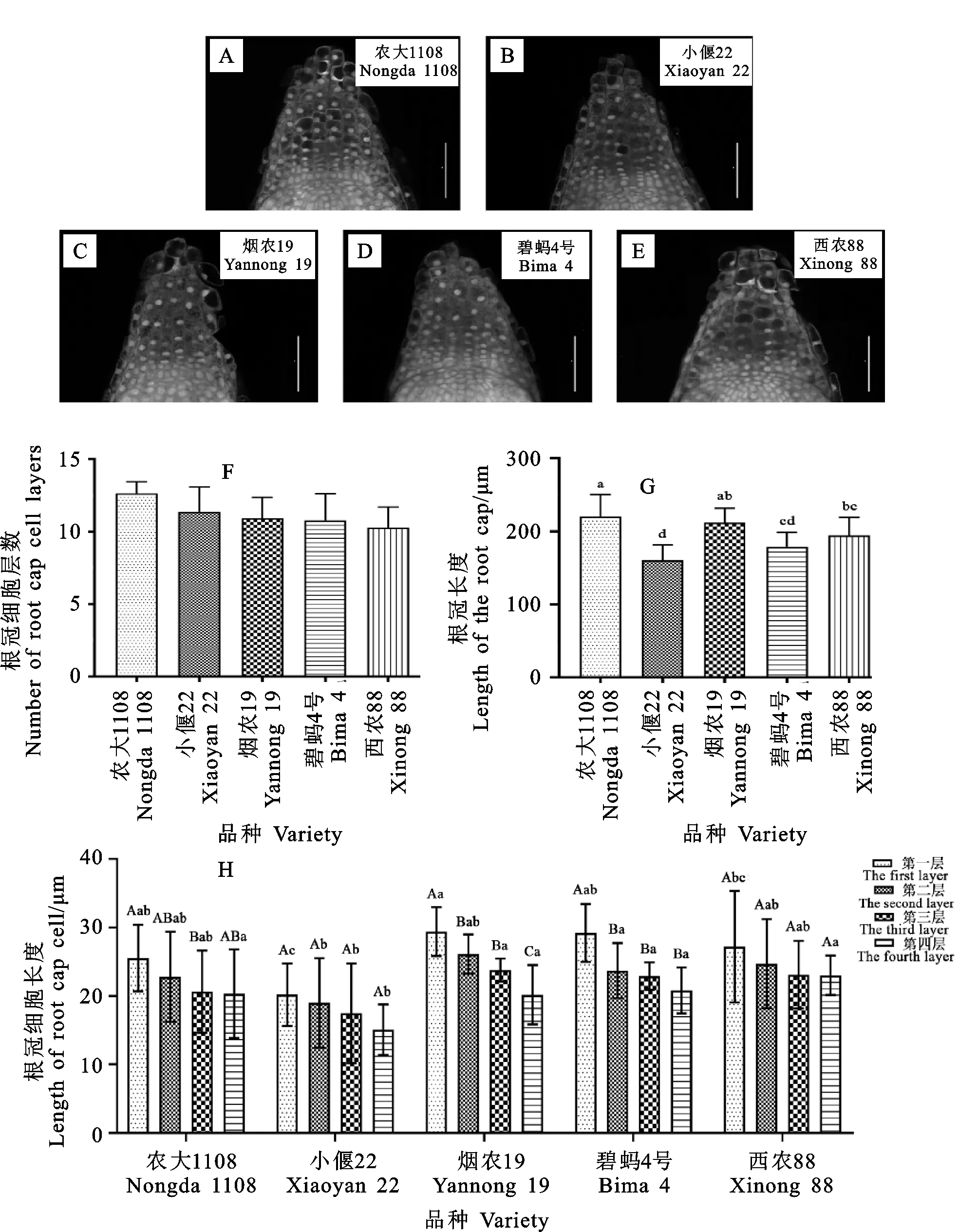
A~E:不同材料根冠纵切结构图(100×);F:根冠细胞层数统计;G:根冠长度统计;H:根冠每层细胞长度统计,不同大写字母表示不同层细胞长度间差异显著,不同小写字母表示不同材料间同一层细胞长度差异显著。A—E:Longitudinal section of root caps of different materials (100×);F:Statistics of cell layers of root cap;G:Statistics of root cap length;H:Statistics of cell length for each layer.The different capital letters represent significant differences among cell length of different layers.The different lowercase letters represent significant differences in cell length among different materials.图2 小麦根冠纵切结构观察与分析比较Fig.2 Observation and analysis of longitudinal section of the root cap
2.3 小麦根尖分生区的细胞学特征
在分生区靠近根尖的区域,细胞排列紧密、细胞核大且位于细胞中央,细胞个体小,横向尺寸大于纵向尺寸,呈扁长型紧密排成一列,这与该区域细胞的快速分裂状态相一致(图3)。此外,5个材料的穹窿型结构与导管起始细胞之间都具有三层细胞,且具有相同的后期分化模式,说明静止中心周围的细胞结构稳定。
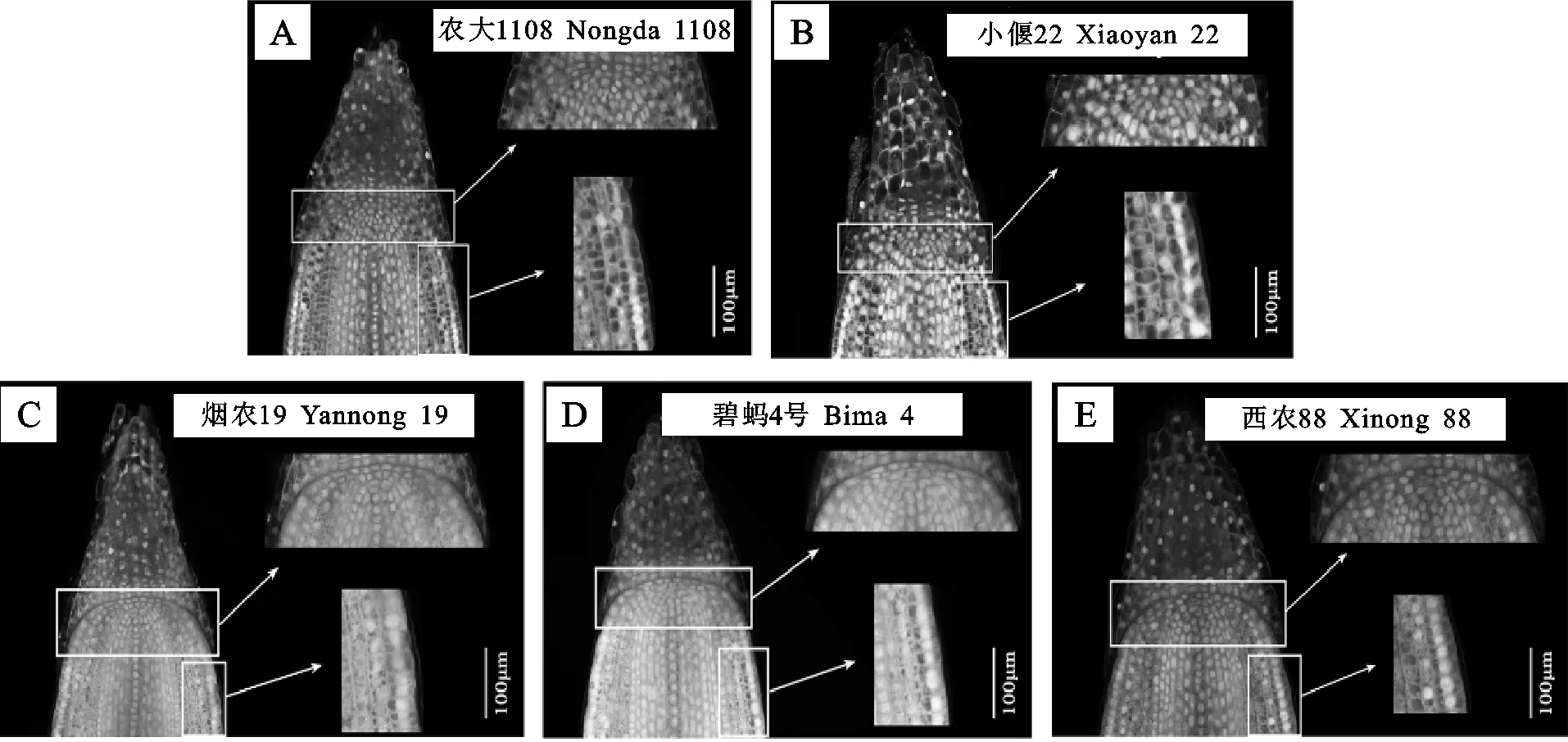
图3 静止中心及分生区细胞放大图(100×)Fig.3 Cytological structure of quiescent center and meristem cells(100×)
在分生区靠近伸长区的区域,细胞的纵向尺寸逐渐变大,并可清晰观察到一层细胞,其横向尺寸和纵向尺寸接近相等,细胞形态近似于正方形。本研究将该层细胞定义为分生区的上边界,并将其与分生区顶端穹窿形结构间的距离定义为分生区的长度(图4A~图4E)。农大1108、小偃22、烟农19号、碧蚂4号和西农88分生区长度的平均值分别为985.85、692.98、711.98、777.19和572.20 μm,农大1108分生区长度约为西农88分生区长度的两倍,说明分生区大小在不同材料间存在较大差异。通过测量,农大1108、小偃22、烟农19号、碧蚂4号和西农88分生区上边界的宽度分别是450.31、388.23、429.93、403.15和415.71 μm,材料间差异不显著(图4F)。
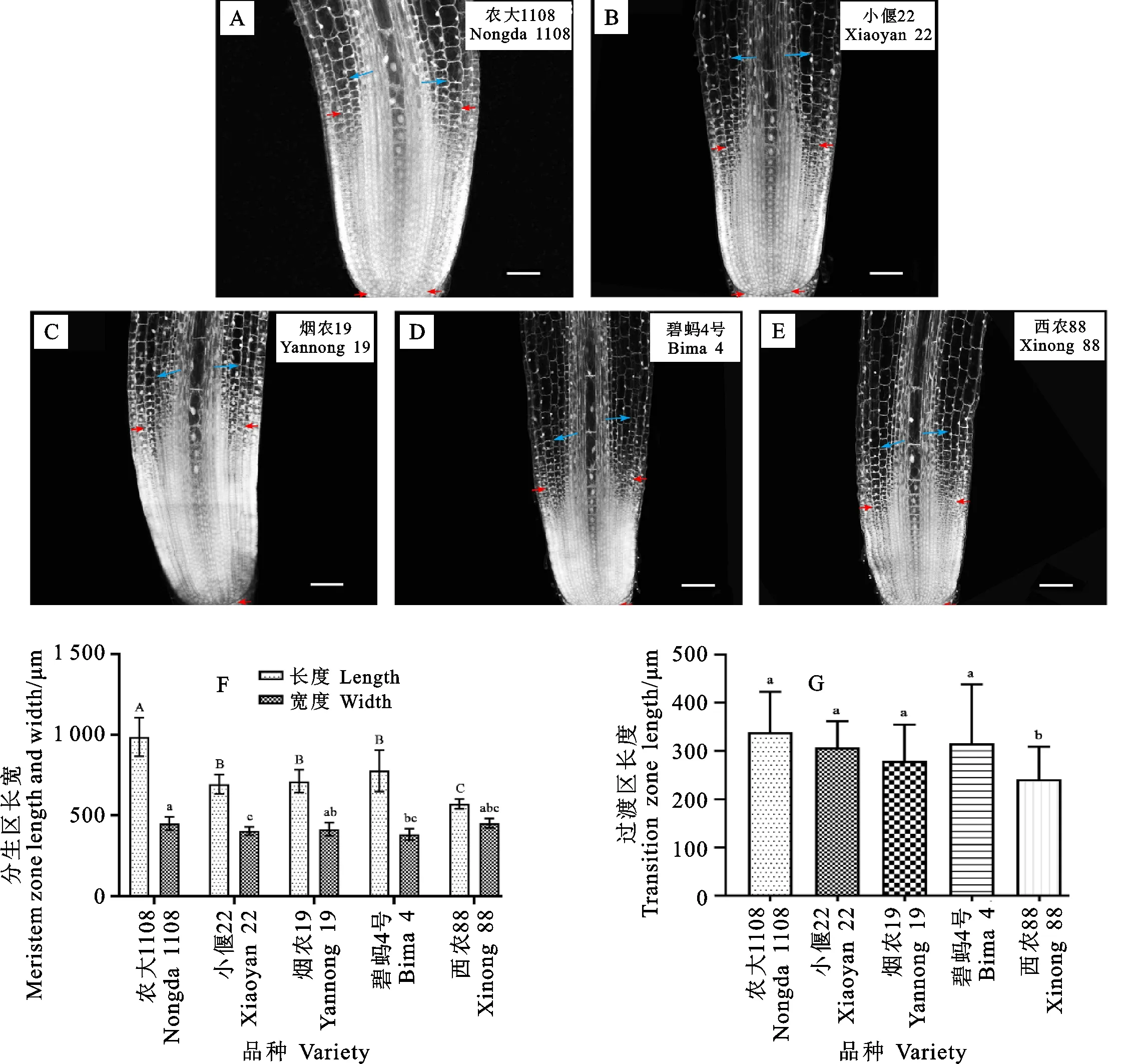
A~E:分生区纵切结构图,红色箭头表示分生区上边界,蓝色箭头表示过渡区上边界(100×);F:分生区长宽统计,不同大写字母表示材料间分生区长度显著差异,不同小写字母表示材料间分生区上边界处宽度差异显著;G:过渡区长度统计,不同字母表示材料间过渡区长度差异显著。A—E:Longitudinal section of the meristem zone.The red arrowhead indicates the upper boundary of meristem zone,and the blue arrowhead indicates the upper boundary of transition zone (100×);F:Statistics of the length and width of the meristem zone.The different uppercase letters represent significant differences of the length of the meristem zone among the five materials,and the different lowercase letters represent significant differences of the width of the meristem zone at the upper boundary;G:Different letters represent significant differences of the length of transition zone among materials.图4 小麦根尖分生区与过渡区纵切结构观察与分析比较Fig.4 Observation and analysis of longitudinal section of meristem zone and transition zone in wheat root tip
2.4 小麦根尖伸长区观察
与模式植物的过渡区相似[10,19],小麦胚根过渡区细胞也可分为两部分,其中靠近分生组织的部分长度略有增加,细胞核位于细胞中心,细胞形状仍近似等径,而远离分生组织的部分,细胞明显伸长。农大1108、小偃22、烟农19号、碧蚂4号和西农88的过渡区长度分别为191.76、214.46、214.89、176.52和133.74 μm,包含的纵向细胞层数分别为7、8、9、6和5层。材料间过渡区长度和细胞层数差异不显著,说明过渡区的细胞模式在材料间较为保守(图4G)。
紧邻过渡区的部分为伸长区,其细胞分裂能力减弱,但伸长和分化迅速,成为根尖深入土层的重要推动力[8]。类似于模式植物拟南芥根系伸长区的细胞结构[20],小麦根尖伸长区细胞的细胞核被挤向侧壁,细胞快速伸长,纵向尺寸迅速增加,细胞形状为矩形(图5A)。为了评估伸长区细胞的伸长速度,我们测量了紧邻过渡区的5~7个细胞的纵向长度,发现其伸长速度总体逐渐加快(图5B)。
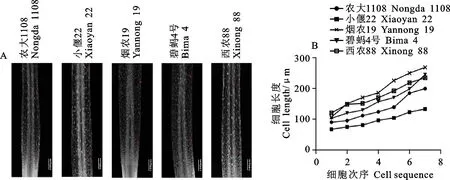
A:不同材料纵切结构图,红色五角星表示用于测量伸长速率的始末细胞;B:伸长速率统计。A:Longitudinal section of elongation zone.The red pentagons represent the beginning cell and ending cell used for measuring elongation rate;B:Elongation rate statistics.图5 伸长区纵切结构观察与分析比较Fig.5 Observation and analysis of longitudinal structure of elongation zone
2.5 小麦根尖成熟区观察
小麦胚根成熟区细胞结构和拟南芥、水稻和玉米的成熟区细胞结构相似,细胞形状呈规则矩形,每层细胞的长度大致均匀。横切观察发现,内皮层细胞直径远远小于皮层细胞[13],因此将与皮层细胞相邻的一层形态与皮层细胞完全不同的细胞层定位为内皮层,内皮层向内为中柱,向外为皮层,最后一层为表皮,由于内皮层细胞和中柱细胞的细胞壁出现木栓化,所以其荧光信号比相邻的皮层细胞更强(图6A)。据此标准,农大1108、小偃22、烟农19号、碧蚂4号和西农88的皮层数分别为4、3、3、2和3层(图6E)。
对各材料成熟区不同类型细胞长度和宽度的测量发现,虽然表皮、皮层及内皮层的细胞长度因材料不同存在差异,但皮层的细胞体积总体上最大,内皮层的细胞体积最小(图6B、6C)。同时需要注意的是,不同材料的趋势不同,小偃22、碧蚂4号和西农88的表皮细胞长度大于皮层细胞长度,而农大1108和烟农19号的皮层细胞长度大于表皮细胞长度。另外,细胞宽度在材料间也存在显著差异,农大1108的皮层宽度最大 (9.35 μm),西农88最小(5.48 μm)。这说明成熟区的细胞,不论是表皮、皮层还是内皮层,细胞大小在不同材料间存在较大变异,并且没有固定规律。
另外,统计分析表明,导管直径,中柱直径和根直径在不同材料间都存在差异,且导管直径的差异最大。导管和中柱占根直径的比例在材料间也存在差异,最大的为烟农19号,最小的为小偃22。但是需要注意的是,导管占中柱的比例 (22.33%~25.39%)在材料间差异不显著(图6D、6F),说明中柱和导管的直径存在协同变化的 现象。
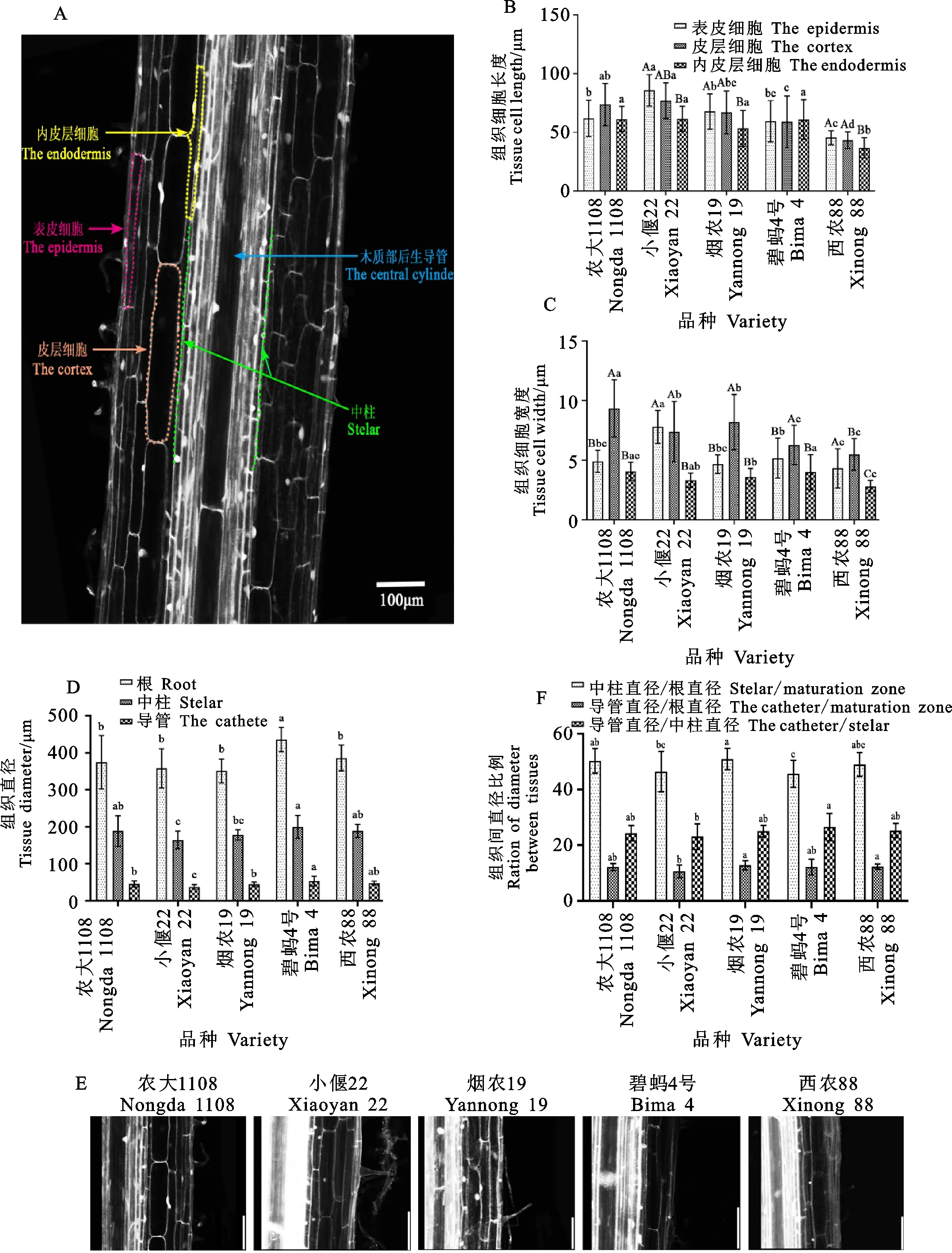
A:成熟区纵切示意图;B:成熟区表皮、皮层、内皮层长度统计;C:成熟区表皮、皮层、内皮层宽度统计;D:成熟区根直径、中柱直径、导管直径统计;E:不同材料成熟区皮层纵切结构图;F:导管直径占中柱直径比例,以及两者占根直径的比例。不同大写字母表示组织间显著差异,不同小写字母表示材料间显著差异。A:Longitudinal section of maturation zone;B:Statistics of the length of the epidermis,the cortex,and the endodermis the maturation zone;C:statistics of the width of the epidermis,the cortex,and the endodermis in the maturation zone;D:Diameter statistics of maturation zone,central column,and catheter;E:Longitudinal section of cortex in maturation zone of different materials;F:The ratio of the diameter of the catheter to the stellar,the ration of catheter to maturation zone,and the ration of the catheter to stellar.The different capital letters represent significant differences among tissues,and the different lowercase letters represent significant differences among materials.图6 小麦根尖成熟区纵切结构观察与分析比较Fig.6 Observation and analysis of longitudinal section of maturation zone in wheat root tip
3 讨 论
根系作为植物最重要的地下器官,具有固着植物,吸收、贮藏、运输水分和营养物质的功能[22]。对于小麦而言,根系形态和性能直接影响地上部分的生长发育和产量表现,在小麦整个发育过程发挥着重要作用[22]。
但目前快捷、清晰、准确的小麦根系细胞学观察技术缺乏,限制了小麦的根系生物学研究。本研究对脱水、透明和放大倍数进行了一系列调整,可以完整清晰观察到小麦根尖的纵向细胞结构,并通过对五个小麦材料胚根的纵向细胞结构进行观察和比较,证明该观察体系的简便和可靠性。在测序成本日益下降的今天,高通量、准确的表型鉴定成为作物学研究中的瓶颈,限制着农艺性状控制位点的定位和分析。本研究中的观察体系,可以快速准确获得包括根冠长度、分生区长度、过渡区长度、皮层细胞长度和宽度、中柱和导管直径在内的多项表型数据,可为定位和分析其遗传调控位点提供可靠的技术支撑。
根冠呈帽状,处于根部的顶端,由细胞壁较薄的细胞组成,对内部组织的保护和重力感知起关键作用。本研究观察发现,材料间根冠长度存在差异,并由细胞层数和每层细胞的长度差异共同造成,根冠长度是否与其保护作用和重力感知有关,还有待进一步研究。同时,我们观察到根冠边缘细胞结构松散,两侧细胞数量不对称,可能与根冠和土壤摩擦脱落有关。
分生区作为小麦根系中细胞分裂最旺盛的区域,其细胞一部分向根尖部分分裂,不断补充脱落的根冠细胞[23],另一部分向根基部分裂分化形成各种功能细胞和组织。另外,分生区长度能直接影响根系的生长[22],与根系长度具有显著正相关关系[19]。拟南芥分生区长度约为200 μm,玉米分生区距离根尖长2 500 μm。本研究发现,小麦分生区长度约为500~1 200 μm,远大于拟南芥,而小于玉米,说明分生区长度在物种间存在显著差异。另外,不同小麦材料的分生区长度也存在显著差异,为解析其遗传调控网络提供了表型依据。但是,分生区长度的差异具体是由细胞数目的差异,还是细胞体积的差异导致的,抑或是两者都有贡献,在目前的研究中并不能确定,因为在目前的放大倍数下,分生区细胞太小,无法计数,放大观察倍数后,分生区分处多个视野,无法有效合并。在拟南芥研究中,有通过连续方程和运动学方法量化细胞增值分裂,从而计算细胞分裂速率及细胞数目的报道[24-25];在玉米研究中推导分生区细胞数目的方法是将根尖连续切片,使用细胞计数器对所有切片细胞数目进行统计分析[26]。参考以上方法,结合本研究建立的观察体系以及发现的细胞体积形态、分生区长度、分界线宽度和横向细胞结构等结果,可以对小麦根尖分生区的细胞组成进行进一步的分析。
静止中心位于分生区顶端,由不太活跃的细胞组成,对维持其周围的干细胞状态至关重要[27]。Hanstein于19世纪中期提出,根尖原始细胞分为3层,分别分化为根冠、皮层和中柱,这种根据原始细胞所组成的层次来解释根尖结构的学说一直沿用至今。在拟南芥中,皮层与内皮层具有共同来源,表皮与侧根冠具有共同来源,中柱独立起源[28];在水稻中,表皮、皮层与内皮层具有共同起源,根冠与中柱分别是独立起源,这两种模式被认为是双子叶植物与单子叶植物的区别[12]。但小麦中因缺乏合适的观察体系,其静止中心的大小和细胞组成鲜有报道。本研究观察到小麦根尖原始细胞大致分化为三层,皮层与内皮层由一层分化而来,而表皮与侧根冠由两层独立分化而来,不仅与拟南芥中的组织模式不同,也与水稻中观察的表皮与皮层具有共同来源不符。但是,这种差异,还需要借助静止中心和起始干细胞特异表达基因和荧光标记,通过对转基因材料的细胞学观察等方法进行确定。
过渡区位于分生区与伸长区之间,可以响应环境变化快速调整生长速度和根系生长方向,是根系发育可塑性细胞的动态储库[21]。靠近分生区的部分其细胞具有分裂能力,且形态与分生区细胞相似,一直以来被认为是分生区的一部分[19]。后来的研究表明,靠近伸长区的过渡区细胞具有快速伸长的能力,因此过渡区作为一个独特的区域能使根细胞获得特殊的生理特性,如对不同环境因素的高度敏感性:重力、湿度、光和氧,这些信号介导根的向性并对根的形态发生都有重要的影响[29]。拟南芥过渡区长度大约为320 μm,本研究观察到的过渡区长度为200 ~350 μm,与拟南芥类似,在小麦材料间过渡区长度和纵向细胞层数差异也不明显,说明小麦根尖过渡区细胞结构比较稳定。
成熟区的细胞长度和宽度趋于稳定,细胞基本停止生长,内部的木质部是根部运输水分和无机盐的主要部位,部分表皮突起分化为根毛,大大扩大了根系的吸收面积[30]。前人研究表明,较少的皮层细胞层数和较大的皮层细胞体积,可以降低代谢消耗并提高营养元素向上分配的能力[25,31-32]。本研究结果表明,材料间细胞层数存在1~2层的差异,皮层细胞较其他类型细胞体积偏大,但在材料间,细胞大小没有固定规律,某种类型细胞体积更大,会引起根系哪些功能差异有待进一步研究。成熟区内的导管直径,中柱直径和根直径的大小,以及前两者所占根直径的比例,都会影响水分和养分在根系中的吸收运输[33-34]。本研究发现,皮层细胞体积、导管和中柱直径在不同小麦材料间存在显著差异,未来可以结合水分和养分吸收和利用实验,揭示这些差异与水分和养分利用效率的关系,为其遗传改良提供理论 基础。
猜你喜欢
现代园艺(2022年7期)2022-11-19
中华实验眼科杂志(2022年6期)2022-11-15
林业科技(2022年5期)2022-10-08
小学生学习指导(低年级)(2020年10期)2020-11-26
湖北农业科学(2019年22期)2019-12-23
首都体育学院学报(2019年5期)2019-10-18
作文大王·低年级(2017年11期)2017-12-05
中国当代医药(2017年17期)2017-07-25
学苑创造·A版(2017年1期)2017-01-19
支部建设(2016年18期)2016-11-28
