
小麦TaGeBPL基因的克隆及表达分析
2021-12-01 09:06王蒙蒙张香宇徐晓敏王长有吉万全
麦类作物学报 2021年9期
王蒙蒙,张香宇,张 露,徐晓敏,王 琦,王长有,吉万全,2,张 宏,2
(1.西北农林科技大学农学院,陕西杨陵 712100;2.农业部作物基因资源与种质创制陕西科学观测实验站,陕西杨凌 712100)
表皮毛是由表皮细胞发育而来,是植物表皮组织的一种特化结构,可增加表皮层厚度,减少水分、热量的散失,也会分泌某些化学物质来防御病虫、微生物的侵害,对植物生长发育具有重要保护作用[1-2]。表皮毛被认为是研究植物细胞分化调控的理想模型[3],同时也具有抗虫、抗病、抗逆等功能[4]。因此,研究与植物表皮毛相关的基因不仅为细胞分化和植物发育提供参考,对小麦遗传改良及育种工作也具有借鉴意义[5]。
近年来,随着拟南芥表皮毛发育调控机制研究的不断深入,多个参与调控植物表皮毛发育的相关基因已被报道,如GL1[6]、TTG1[7]、GL3[8]、TTG2[9]和GL2[10]。其中,GL1是第一个被克隆到的与表皮毛发育相关的关键基因,其增强子序列对基因发挥功能具有重要作用[11]。研究表明,表皮毛细胞发育与其他形态建成类似,是一个严格受时空调控的过程,需要不同的转录因子相互作用完成[12-13]。GeBP(GLABROUS1-enhancer binding protein)型转录因子作为其中转录因子之一,通过识别并结合GL1基因增强子来调控GL1基因的表达,进而控制表皮毛的生成[14]。GeBP是植物中特有的一类转录因子[15],截至目前,除了拟南芥、水稻等模式植物外,番茄中亦有GeBP转录因子的报道[14,16],但在小麦中还未见到GeBP转录因子的相关报道。
拟南芥中GeBP转录因子是一种具有DNA结合活性的核蛋白,主要在叶原基、分生组织以及维管组织中表达[15],GPL1、GPL2、GPL3在内的GeBP/GPLs家族成员都含有保守中央区域和亮氨酸拉链C端区域,并且在体内可形成二聚体[17],但目前其功能未知。前人研究发现,GeBP蛋白与CPR5(Pathogenis-related gene 5)蛋白存在一定的联系,暗示GeBP基因发挥其功能可能与CPR5基因有关[18]。CPR5是一个多效性基因,在细胞壁代谢[19]、细胞增殖[20]、细胞衰老[21]、细胞死亡[22]、植物抗病性[23-24]方面中发挥重要功能,而GeBP可通过调节CPR5基因来发挥作用[18]。此外,遗传学分析也表明,GeBP发挥自身功能在一定程度上依赖于CPR5[18]。在病菌胁迫过程中,CPR5是调控植物细胞防御的重要因子之一[25]。Perazza等[18]对gebp gpl1,2,3缺失突变体转录组结果数据分析中发现,GeBP/GPLs和CPR5转录组数据存在部分重叠,CPR5通路中发现了大量的GeBP/GPLs依赖基因,GeBP/GPLs基因控制着CPR5调控的部分基因,作用于CPR5通路下游,GeBP/GPLs的功能虽是CPR5功能的一个子集,但GeBP/GPLs和CPR5之间的转录组分析数据、表型和遗传关系均表明,两者参与了类似的过程。例如,gebp gpl1,2,3缺失突变体、VP16:GPL2过表达突变体、cpr5缺失突变体这三个株系接种丁香假单胞菌(Pseudomonassyringaepvtomato)DC3000后,三个株系中病原体的生长量较对照组野生型均明显下降,说明有关基因激活了病原体的应答通路,抑制病原体的入侵,并且gebp gpl1,gebp gpl2,gebp gpl3缺失突变体和VP16:GPL2过表达突变体株系中病原菌应答标记基因PR1和PR5的转录水平也高于对照组野生型,表明GeBP/GPLs是PR基因的抑制因子,由此也证明GeBP转录因子在病原菌防御途径中发挥着一定作用[18]。
本课题组前期对小麦自发斑点细胞坏死RILs(N13039H/N)转录组学数据分析时发现,GeBP同源基因在两个不同材料中的表达水平差异显著[26]。鉴于小麦GeBP家族转录因子参与生物、非生物胁迫以及对病原菌的抵抗防御反应尚不清楚,因此,本研究克隆了小麦中一个GeBP类基因,并对其进行生物信息学分析、亚细胞定位、表达特性分析以及酵母转录激活活性验证,以期为研究小麦GeBP家族基因功能提供参考和依据。
1 材料与方法
1.1 试验材料
供试材料西农N13039H、抗白粉病近等基因系N9134R、感白粉病近等基因系N9134S、本氏烟草(Nicotianabenthamiana)以及小麦白粉菌(Blumeriagraminisf.sp.tritici,Bgt)E09菌株均由西北农林科技大学农学院提供。
1.2 材料处理
将均匀一致、成熟饱满的N9134R和N9134S籽粒种于盆中,置于20 ℃/12 ℃(光照16 h/黑暗8 h)人工培养箱中培养,待小麦长至两叶一心期时,接种E09菌株。在接种0、12、24、36、48和72 h后分别取适量叶片提取RNA,置-80 ℃保存。
1.3 基因组DNA、总RNA的提取及cDNA第一链的合成
利用Trizol法[27]提取西农N13039H、抗白粉病近等基因系N9134R、感白粉病近等基因系N9134S各样品的总RNA。用PrimeScriptTMⅡ 1st strand cDNA Synthesis Kit试剂盒(TaKaRa,大连)合成cDNA第一链。
1.4 目的基因的克隆
根据自发斑点细胞坏死RILs(N13039H/N)的转录组测序数据,利用Ensembl Plants数据库(http://plants.enseml.org/index.html)将搜索到的候选基因(TraesCS3D02G037800)序列为参考序列,利用Primer Premier 5.0设计3D染色体上目标基因特异引物TaGeBPL-F/R(表1),并以西农N13039H cDNA为模板进行PCR扩增。扩增程序:94 ℃ 3 min;94 ℃ 30 s,60 ℃ 50 s,72 ℃ 2 min,38个循环;72 ℃ 10 min。利用1.5%琼脂糖凝胶电泳检测PCR扩增产物,将与预期片段长度一致的目的条带回收、纯化,使用DNA连接酶将目标片段插入pMD19-T载体(TaKaRa,大连),并转化至大肠杆菌感受态细胞DH5α。平板过夜后,利用菌落PCR检测阳性单克隆并测序验证。测序由奥科鼎盛生物科技有限公司完成。

表1 本研究用到的引物信息Table 1 Primers used in the study
1.5 生物信息学分析
通过NCBI(https://www.ncbi.nlm.nih.gov)和Ensembl Plants中BLAST程序对TaGeBPL基因的核酸序列及氨基酸序列进行比对;用ExPASy(http://web.Expasy.org/protparam/)对TaGeBPL蛋白进行理化性质分析;用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和SWISS-MODEL(https://swissmodel.expasy.org/interactive/CKkukm/models/)对蛋白质的二级和三级结构进行预测;用TMpred(http://www.cbs.dtu.dk/services/TMHMM/)和SignalP(http://www.cbs.dtu.dk/services/SignalP-3.0/)进行跨膜区和信号肽预测;用ProtScale(https://web.expasy.org/protscale)和NetPhos(http://www.cbs.dtu.dk/services/NetPhos)对蛋白的亲疏水性和磷酸化位点进行预测;用Plant CARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析基因启动子区域的顺式作用元件;用cNLS Mapper(http://nls-mapper.iab.keio.ac.jp/cgi-bin/NLS_Mapper_form.cgi)和Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2)预测编码蛋白的核定位信号和亚细胞定位;用MEME Suite(http://meme-suite.org/index.html)分析氨基酸保守motif;用NCBICD-Search(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)分析蛋白的保守结构域;用MEGA 7.0软件,采取最大似然法构建系统进化树,Bootstrap值为1 000。
1.6 TaGeBPL基因的表达分析
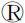
1.7 亚细胞定位
1.8 酵母转录自激活活性验证
与GFP同源重组引物设计原理相同,用Premier 5.0设计引物pGBKT7-TaGeBPL-F/R(表1),以pMD19-T::TaGeBPL质粒为模板进行PCR扩增。使用限制性内切酶EcoRⅠ和BamHⅠ对pGBKT7载体进行双酶切,琼脂糖凝胶电泳检测后对PCR扩增产物和酶切产物进行回收、纯化和同源同组,并转化大肠杆菌感受态细胞DH5α,挑取阳性单克隆。根据奥科鼎盛生物科技有限公司测序结果,提取质粒后获得pGBKT7::TaGeBPL重组载体。
获取重组载体后,以重组载体pGBKT7::TaGEBPL为试验组、共转载体pGBKT7-53+pGADT7-T为阳性对照、空载体pGBKT7为阴性对照,分别转化至酵母菌株Y2HGold中,取适量菌液涂布在SD/-Trp培养基上,封口倒置,置于30 ℃黑暗培养箱中培养48~96 h,观察菌落生长情况。待菌长至2~3 mm,挑取单克隆于 1 mL的SD/-Trp液体培养基中震荡培养(220 r·min-1,30 ℃)至OD600约为0.1,以100、10-1、10-2和10-3倍数稀释菌液,各取2 μL分别点涂在SD/-Trp、SD/-Trp/X-α-Gal、SD/-Trp/-His/-Ade培养基上,置于30 ℃黑暗培养箱中培养 48~96 h,观察菌生长情况,进行转录因子自激活检测。
2 结果与分析
2.1 目的基因的扩增
以小麦西农N13039H的cDNA为模板,以TaGeBPL-F/R为特异性引物进行PCR扩增,得到一条与预期目标相一致的条带(图1),将扩增产物回收纯化后连接至pMD19-T载体,转化至大肠杆菌DH5α,并进行篮白斑筛选、PCR检测和测序,最终获得一条长度为1 527 bp的片段。利用Ensembl Plants基因组数据库将所得序列进行Blast比对分析,结果显示与中国春TraceCS3D02G037800最接近,相似度高达 99.1%,位于小麦3D染色体上,而与小麦3A和3B染色体组上的TraceCS3A02G042900、TraceCS3B02G040600基因的相似度仅为 84.1%和91.6%。目的基因初步分析得出,该基因共编码508个氨基酸(图2),推测其分子式为C2342H3835N695O811S10,理论分子量为55 025.98 Da,理论等电点为6.33。
2.2 TaGeBPL蛋白理化性质和结构域分析
利用ExPASy对目的基因编码蛋白质的理化性质进行分析,发现该蛋白氨基酸序列中赖氨酸和谷氨酸含量较高,脂肪指数为50.12,平均亲水性为-1.153,属于亲水性蛋白,其不稳定系数为49.31,同属于不稳定蛋白。利用SOPMA和SWISS-MODEL预测该蛋白的二级、三级结构,发现该蛋白是由四种构象组成。SignalP和TMpred预测结果显示,该蛋白不含有信号肽和跨膜结构域。NetPhos预测结果表明,该蛋白氨基酸序列含有较多的磷酸化位点。cNLS Mapper预测结果显示,该蛋白含有两个单一型NLS和一个双分型NLS(图2)。Cell-PLoc预测结果显示,该蛋白定位于细胞核内,与NIL预测结果一致。NCBI CD-Search保守结构域分析表明,该蛋白含有DUF573结构域,位于氨基酸第164至247位点之间,属于GeBP转录因子家族,因此将该基因命名为TaGeBPL。
M:DL2000;1~2:PCR扩增产物。M:DL2000;1-2:PCR amplification products.图1 PCR扩增结果Fig.1 PCR amplification results

黑色下划线表示DUF573结构域;方框部分表示核定位信号区。Black underline represent DUF573 domain;Boxes indicate the region of nuclear localization signal.图2 TaGeBPL蛋白的氨基酸序列Fig.2 Amino acid sequence of TaGeBPL protein
2.3 蛋白同源性及系统发育分析
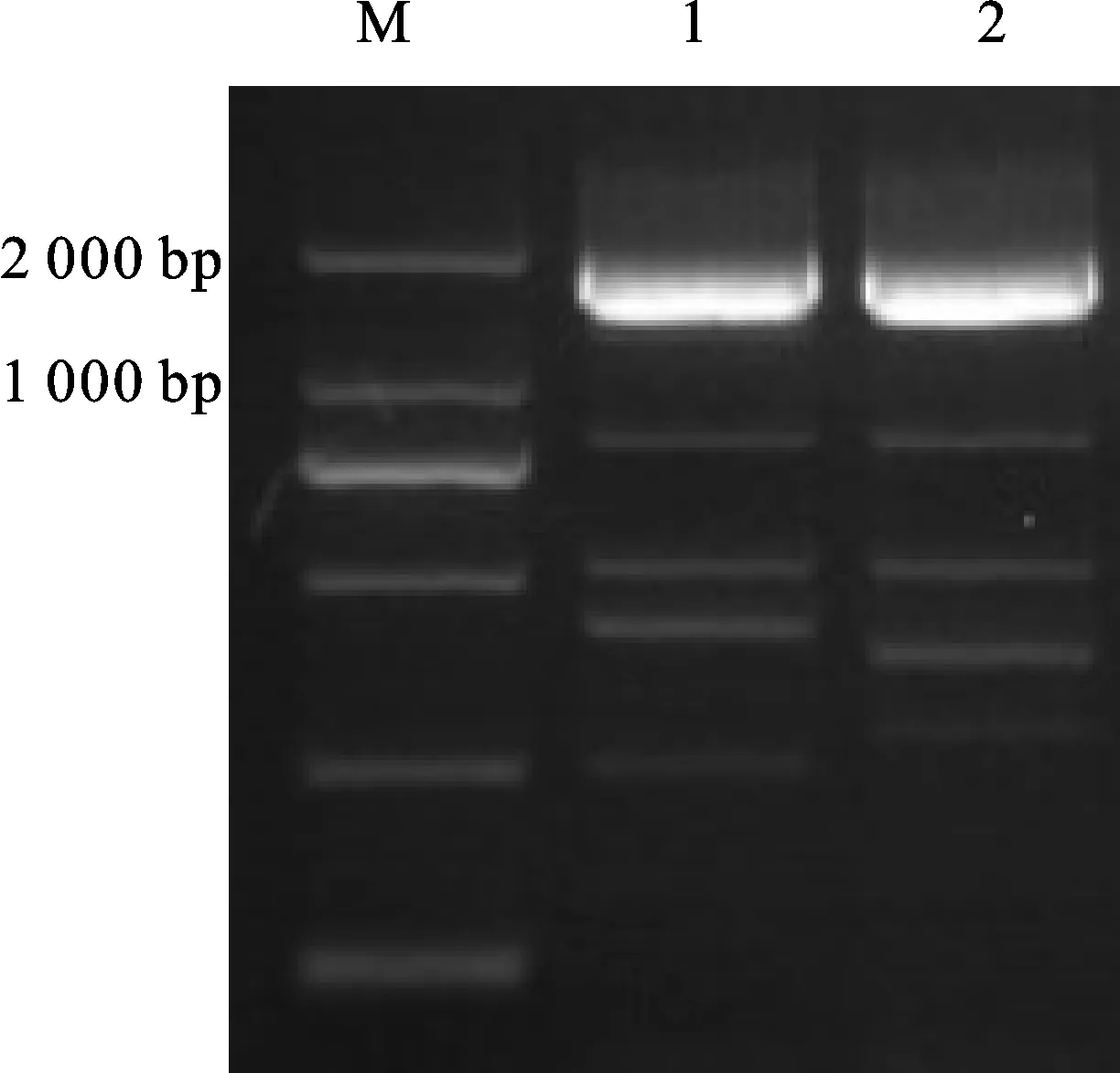
利用PlantTFDB、RGAP、TAIR等数据库检索GeBP家族转录因子,发现在已被鉴定的拟南芥和水稻中,绝大多数GeBP家族成员氨基酸序列的中央区域均含有DUF573结构域。将小麦西农N13039H的TaGeBPL转录因子与拟南芥和水稻中的GeBP转录因子保守结构域进行多序列比对分析,结果(图3)显示,TaGeBPL与AT2G20613.1、AT4G00130.1、AT5G41765.1、LOC_Os01g14720.1的保守域氨基酸序列长度有较大差异性,而与其他氨基酸的序列长度相近,结构域相似性也较高。

图3 TaGeBPL与拟南芥、水稻GeBP蛋白的氨基酸序列比对Fig.3 Sequence alignment of TaGeBPL and GeBP of Arabidopsis thaliana and Oryza sativa
为进一步了解小麦TaGeBPL的进化关系,从PlantTFDB数据库下载拟南芥和水稻中的GeBP转录因子氨基酸序列,通过MEGA 7.0软件构建系统发育树,结果(图4)表明,TaGeBPL与水稻LOC_Os02g18660.1、LOC_Os02g53150.1处于同一分支,亲缘关系较近,相似性较高。
图4 TaGeBPL与拟南芥、水稻GeBP蛋白系统发育分析Fig.4 Phylogeny analysis of TaGeBPL and GeBP of Arabidopsis thaliana and Oryza sativa
2.4 启动子区序列结构分析
利用Ensembl Plants数据库分析TaGeBPL基因起始密码子上游约2 000 bp的序列,结果(表2)发现,TaGeBPL的启动子区域含有与逆境、植物激素相关的顺式作用元件。如在起始密码子上游1 163~1 168 bp处含有MBS干旱应答元件,在上游402~407 bp处含有LTR低温应答元件,在上游1 864~1 868 bp处含有as-1植物激素应答元件,在上游158~162 bp处含有ABRE脱落酸应答元件。此外,启动子区域还含有多个光应答元件(I-box、G-box、Sp1),其位置和数量均不同。启动子分析结果表明,该基因可能受多种植物激素信号调控,参与生物和非生物胁迫途径。

表2 TaGeBPL基因启动子区顺式作用元件Table 2 Cis-acting elements of TaGeBPL gene in the promoter regions
2.5 小麦TaGeBPL基因的表达模式
研究TaGeBPL基因在小麦中的表达模式有利于挖掘其潜在功能,将白粉菌菌株E09接种于抗病近等基因系N9134R和感病近等基因系N9134S中,采用实时荧光定量PCR的方法,对接种白粉菌E09 0、12、24、36、48和72 h后TaGeBPL基因的相对表达量进行测定。结果(表3)表明,接种E09后TaGeBPL基因在N9134R和N9134S两个材料中的表达模式不同。TaGeBPL基因在N9134R中的表达量呈现“降-升-降”的变化趋势,接种12、24和72 h时TaGeBPL基因相对表达量显著下调,分别为0 h时的61%、68%和74%;接种36和48 h时TaGeBPL基因的相对表达量显著上调,分别为0 h的1.23和 1.19倍。而TaGeBPL基因在N9134S中的表达量呈现“升-降-升-降”的变化趋势,在12 h时TaGeBPL相对表达量显著上调,36 h时其表达量达到峰值,约为对照组的2倍,差异显著,72 h时恢复至未接种对照相同的水平。小麦TaGeBPL基因在N9134S中的表达量始终高于在N9134R中的表达量,推测TaGeBPL基因可能响应白粉菌的侵染,参与了病原菌响应的相关 途径。
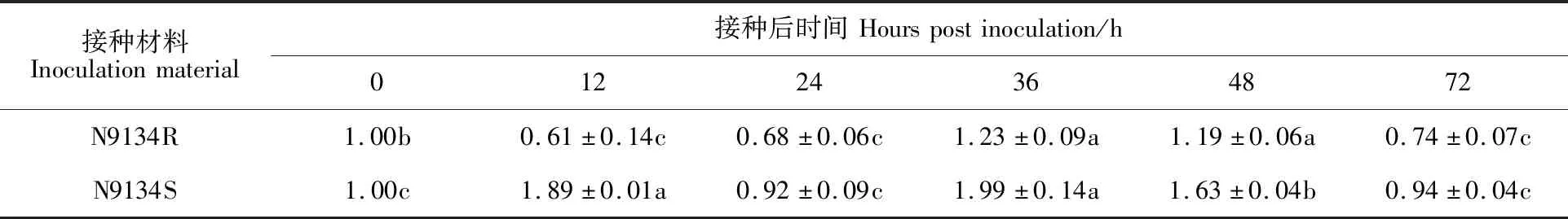
表3 TaGeBPL基因的qRT-PCR分析Table 3 qRT-PCR analysis of TaGeBPL gene
2.6 小麦TaGeBPL的亚细胞定位
NLS Mapper和Cell-PLoc预测结果显示,该蛋白具有3个核定位信号,定位在细胞核上。为验证TaGeBPL的亚细胞定位结果,利用本氏烟草瞬时表达系统,以PYJ::GFP空载体为对照组,以PYJ:TaGeBPL:GFP融合载体为试验组,注射生长良好的烟草植株,结果(图5)显示,在绿色荧光通道下,PYJ:TaGeBPL:GFP融合蛋白仅在细胞核内有绿色荧光信号,说明该蛋白定位于细胞核中。
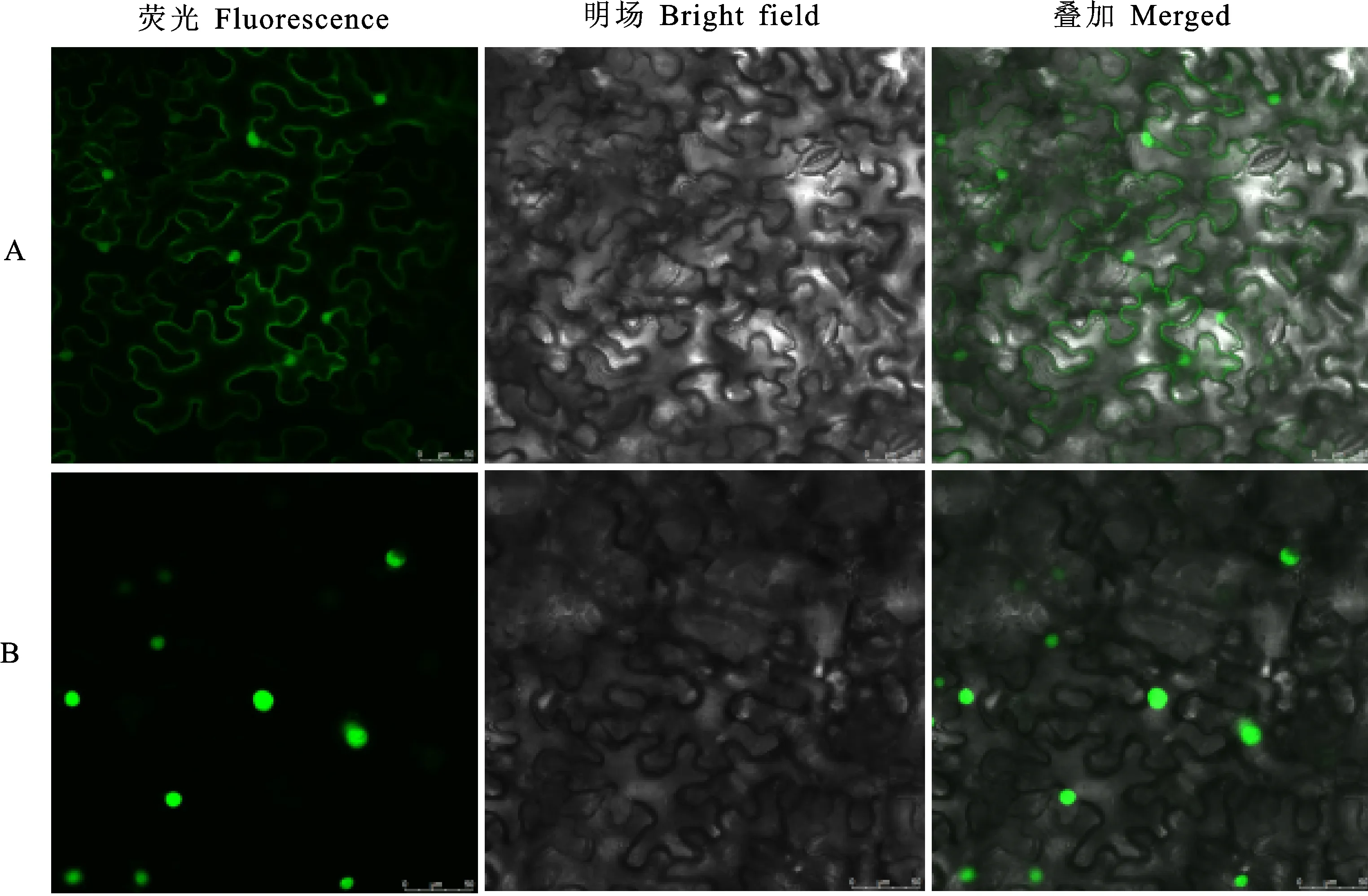
A:对照PYJ::GFP空载体定位;B:PYJ:TaGeBPL:GFP融合载体定位。A:Localization of control PYJ::GFP empty vector;B:Localization of PYJ:TaGeBPL:GFP fusion vector.图5 小麦TaGeBPL的亚细胞定位Fig.5 Subcellular localization of wheat TaGeBPL protein
2.7 TaGeBPL的转录激活活性
转录因子的转录激活活性对其发挥生物学功能具有重要作用,为检测TaGeBPL转录因子的转录激活活性,将试验组pGBKT7::TaGeBPL、对照组pGBKT7-T+pGBKT7-p53和pGBKT7分别转至酵母Y2HGold菌株中,发现它们均能在SD/-Trp营养缺陷培养基上正常生长(图6),说明外源载体pGBKT7::TaGeBPL成功转入酵母菌中。为检验pGBKT7::TaGeBPL融合蛋白对其下游报告基因的激活情况,将含有pGBKT7::TaGeBPL、pGBKT7-T+pGBKT7-p53、pGBKT7的酵母菌株分别涂布于SD/-Trp/X-α-Gal、SD/-Trp/-His/-Ade营养缺陷型固体培养基上,结果(图6)发现,含pGBKT7::TaGeBPL的Y2HGold菌株不能激活LacZ报告基因表达,使X-α-Gal发生显色反应,说明诱饵载体pGBKT7::TaGeBPL无法激活Y2HGold酵母菌株中指示基因的表达。在SD/-Trp/-His/-Ade营养缺陷培养基上,pGBKT7::TaGeBPL酵母菌株不能正常生长,证实了TaGeBPL在酵母细胞中无转录自激活活性。

100、10-1、10-2、10-3为4种不同稀释浓度酵母菌液;pGBKT7::TaGeBPL为试验组;pGBKT7-T+pGBKT7-p53为阳性对照组;pGBKT7为阴性对照组。100,10-1,10-2 and 10-3are four different dilution concentration of yeast solution;pGBKT7::TaGeBPL is experimental group,pGBKT7-T+pGBKT7-p53 is positive control group;pGBKT7 is negative control group.图6 小麦TaGeBPL转录激活活性分析Fig.6 Transactivation activity assay of TaGeBPL in wheat
3 讨 论
本研究基于前期转录组测序结果,利用RT-PCR技术克隆得到小麦TaGeBPL基因。该基因仅存在于染色体第3同源群,且与现阶段已知的小麦唯一GeBP基因(TraesCS1D02G132000)同源性低于30%。拟南芥中,GeBP转录因子家族成员的基因结构较简单,在染色体上随机分布,含有DUF573结构域[14],与前人研究相同,TaGeBPL蛋白也含有DUF573结构域,但其功能未知。对拟南芥、水稻和小麦GeBP基因编码蛋白的保守结构域进行比对,发现它们具有较高的相似性,TaGeBPL同水稻LOC_Os02g18660.1、LOC_Os02g53150.1亲缘关系较近,而LOC_Os02g18660.1和LOC_Os02g53150.1在植株顶端分生组织和种子中的表达量较高,在生殖生长阶段发挥重要功能[16],推测小麦中TaGeBPL蛋白可能具有相似功能。在利用生物信息学分析TaGeBPL的基础上,本研究进行了基因时空特异性表达检测,qRT-PCR结果表明,白粉菌接种后各个时间点该基因在抗感材料中的相对表达量不相同,且在各个时间点感病材料中TaGeBPL基因的表达量始终高于抗病材料中的表达量,推测小麦中TaGeBPL基因参与了病原菌响应途径,与GeBP基因在拟南芥中的研究结果[17]一致,这不仅为后期研究小麦GeBP基因作用与机理奠定基础,也为探究拟南芥等其他物种GeBP同源基因的功能提供参考。此外,拟南芥GeBP蛋白可在体内形成二聚体[17],可从与之互作的蛋白为出发点,间接地研究目标蛋白在植物体中的更多功能[28]。需要注意的是,拟南芥中二聚体的形成与亮氨酸拉链区域的最后四个氨基酸有密切的关系,而小麦TaGeBPL蛋白C端没有亮氨酸拉链特殊区域,可能不同物种或同一家族的不同成员之间有差异,总的来说,研究蛋白二聚体为分析GeBP转录因子家族提供了思路,对其功能挖掘也具有重要意义[29]。
功能分析是研究基因的一个方面,探究其作用机理可更加深刻地认识该基因。本研究利用cNLS Mapper发现,TaGeBPL转录因子位于细胞核中,具有两种类型的核定位信号,这一特征表明TaGeBPL转录因子不需要依靠其他具有NLS转录因子的介导,可以直接被受体蛋白识别进入细胞核内[30]。这与拟南芥GeBP/GPLs不同,拟南芥GPL3蛋白虽然定位于细胞核内,但并未发现其具有核定位信号[17],这表明不同物种GeBP进入细胞核的方式可能不同。亚细胞定位结果表明,TaGeBPL仅在细胞核中有绿色荧光信号,进一步验证了其在核中发挥作用。在拟南芥中GeBP也位于植物细胞核中,而水稻中GeBP家族成员不仅存在于细胞核中,还存在于细胞质、叶绿体类囊体薄膜上[16],原因可能是在不同物种GeBP参与了不同的生理过程。
植物中众多转录因子参与植物激素调控途径[31],GeBP转录因子也不例外。如在模式植物中,Chevalier等[17]发现,拟南芥GeBP/GPLs基因通过对A型ARR基因的调控,进而参与细胞分裂素反应,且对细胞分裂素的响应具有冗余作用[17];而石 蕾[16]研究表明,水稻LOC_Os06g10710在乙烯处理后表现出敏感响应,说明不同物种的GeBP转录因子家族成员可能参与不同的植物激素信号调节途径。在模式植物中,GeBP/GPLs对细胞分裂素的作用仅发生在特定的组织和器官中,而并非整个植株中都能发挥此作用[17]。此外,此方面的研究对象是具有亮氨酸拉链的GeBP家族成员,无亮氨酸拉链的其他成员是否参与植物激素调节是未知的。而在小麦中该蛋白不含有明显特征的亮氨酸拉链,所以TaGeBPL能否响应细胞分裂素调控途径还需要下一步的研究。值得注意的是,转录因子调控功能基因的表达通常是由相同或不同转录因子协同作用的结果,本研究对TaGeBPL的转录激活活性试验为后续互作蛋白的筛选研究奠定了基础。
猜你喜欢
今日农业(2022年16期)2022-11-09
湖北农业科学(2022年11期)2022-07-18
今日农业(2021年8期)2021-11-28
金桥(2021年10期)2021-11-05
今日农业(2021年13期)2021-08-14
实用肿瘤学杂志(2020年4期)2020-12-08
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
医学综述(2011年12期)2011-12-09
亚热带农业研究(2011年3期)2011-09-29
