
新疆野苹果多倍体诱导及叶片特征分析
2022-01-12 08:35张丞慧李凯铭赵琪于洁贡嘎益西胡宝全
天津农学院学报 2021年4期
张丞慧,李凯铭,赵琪,于洁,贡嘎益西,胡宝全
新疆野苹果多倍体诱导及叶片特征分析
张丞慧,李凯铭,赵琪,于洁,贡嘎益西,胡宝全通信作者
(天津农学院 园艺园林学院,天津 300392)
新疆野苹果是苹果重要的野生资源和砧木,挖掘其利用方法有利于苹果育种工作的进一步开展。本研究采用新疆野苹果实生幼苗为材料,通过不同浓度秋水仙素的浸渍处理诱导四倍体,并观察其叶片特征结构的变化。结果表明,0.2%的秋水仙素处理幼苗1 d能取得较好的诱导效果,新疆野苹果四倍体相对于二倍体,其叶片加厚,气孔密度降低,叶绿素含量增加,叶片解剖结构发生变化并导致叶片紧实度增加、疏松度下降,表现出高抗逆性的叶片特征。研究结果可为新疆野苹果种质资源的开发提供新途径,并为苹果新型优质砧木的培育和苹果的倍性育种工作提供新的参考和理论支持。
新疆野苹果;四倍体;秋水仙素;叶片;解剖结构
新疆野苹果是蔷薇科苹果属植物,原产于中亚和我国的新疆地区,属于第三纪孑遗物种,已经被列为国家濒危二级保护植物,是珍贵的苹果野生种质资源,且具有与栽培品种亲和力强、种质资源丰富、耐寒和耐盐碱等特点[1],因此在陕 西、山东、河北、新疆等地被广泛用作栽培苹果的砧木[2]。
多倍体植物广泛存在于被子植物中,几乎所有的被子植物在进化中都经历过古多倍体事件,多倍体化也是新物种形成的重要途径。而果树多倍体化后一般具有营养生长旺盛、果实大和抗逆性强等特点[3],因此多倍体育种是苹果种质创新的重要手段之一。目前栽培苹果中,大部分品种为二倍体,但部分品种出现三倍体或四倍体,如‘乔纳金’‘北海道’‘金冠’和‘嘎拉’等[4]。苹果多倍体可通过自然突变或人工诱导两条途径产生,其中自然突变包括体细胞突变和减数分裂不分离的生殖细胞突变。通过对大量苹果实生苗的染色体鉴定发现,二倍体自然授粉可产生三倍体,占比0.3%,也可产生四倍体,约占0.1%[5],但自然突变几率低,获得多倍体筛选工作量大。因此人工诱变是获得苹果多倍体的重要途径,其中使用最广泛的是秋水仙素诱导。如0.5%的秋水仙素浸渍‘金冠’的成熟胚2 d后,可诱导苹果四倍体,诱导率可达10.6%,‘寒富’和‘烟嘎’苹果的试管苗叶片在含有秋水仙素的液体再生培养基中培养,也可诱导四倍体,诱变率在5.3%~22.2%[6]。含有25 mg/L秋水仙素的不定芽再生培养基上,诱导‘皇家嘎拉’苹果外植体,可获得四倍体,诱导率最高为36.7%[7]。
苹果多倍体的诱导虽然取得一些进展,但苹果砧木的多倍体化研究进展较慢。而目前对新疆野苹果的研究集中于对新疆野苹果的繁育特性、抗逆性生理机制和野苹果林的保护等方面,对于新疆野苹果种质资源的创新工作进展较为缓慢,目前尚未发现有学者对新疆野苹果进行多倍体化的种质资源创新。本研究利用不同浓度的秋水仙素对二倍体新疆野苹果诱导加倍,获得四倍体新疆野苹果植株,筛选最适的诱导条件,并对四倍体新疆野苹果的叶片结构特征进行探究,为新疆野苹果的种质创新提供新的途径和理论基础。
1 材料与方法
1.1 试验材料
试验以展开子叶的新疆野苹果幼苗为材料,种子来源于经鉴定为二倍体的5年生新疆野苹果树。将新疆野苹果种子室温浸泡过夜,然后将剥去种皮的种子置于培养皿中,覆盖润湿的纸巾,于22 ℃温箱中催芽,待种子展开子叶未抽真叶时,种植于基质(草碳∶蛭石∶珍珠岩=1∶1∶1)中培养。
1.2 试验方法
1.2.1 四倍体新疆野苹果诱导
以水为对照,室温条件下将含有不同浓度秋水仙素溶液(0.20%、0.10%、0.05%、0.01%)的棉花,浸渍新疆野苹果幼苗茎尖生长点24 h,蒸馏水清洗后正常培养,每个处理梯度25株,3次重复。
1.2.2 染色体鉴定
饱和对二氯苯溶液中,室温浸泡茎尖3 h,蒸馏水清洗后,卡诺固定液固定12 h,取出于1 mol/L盐酸中60 ℃解离10 min,蒸馏水清洗3次后,卡宝品红染色1 h。于载玻片上经盖玻片压片后,显微镜下镜检,选取5个视野对染色体计数,并判定多倍体。
1.2.3 气孔及保卫细胞测量
撕取新疆野苹果叶片表皮于载玻片上,Leica DM4000B 显微镜观察,LAS4.2软件测量气孔及保卫细胞长宽,每个植株选取5片叶片,选取5个视野,统计每个视野下气孔数,并测量其中5个气孔及保卫细胞的长宽,计算气孔密度,水处理植株作为对照,数据使用SPSS 19.0,采用方差分析进行显著性检验。计算处理后的存活率和诱导率,并使用SPSS 19.0进行单因素方差分析和多重比较。
气孔密度(个/mm2)=气孔个数/视野面积 (1)
1.2.4 叶片解剖结构观察
每株新疆野苹果取健康叶片,以主脉为对称轴,切成宽度为5 mm的小段,于FAA固定液中处理24 h后,梯度乙醇脱水,二甲苯透明,石蜡包埋,于Leica RM 2235切片机上切片,番红固绿染色,中性树胶封片,Leica DM4000B显微镜观察,LAS 4.2软件测量叶片上下表皮、栅栏组织、海绵组织、叶片和叶肉厚度,计算紧实度、疏松度、上下表皮相对厚度,每株叶片样本测量10次,数据使用SPSS 19.0,采用方差分析进行显著性检验。
紧密度/%=栅栏组织厚度/叶片厚度×100 (2)
疏松度/%=海绵组织厚度/叶片厚度×100 (3)
上表皮相对厚度/%=上表皮厚度/叶片厚度×100 (4)
下表皮相对厚度/%=下表皮厚度/叶片厚度×100 (5)
1.2.5 叶片叶绿素含量测定
各取植株叶片1 g,于研钵中加入少许石英砂和5 mL 95%乙醇,充分研磨,直到残渣变白,液体和残渣转移至15 mL离心管,少量95%乙醇冲洗,定容至10 mL,置于4 ℃冰箱中,静置12 h后,以2 000 r/min离心10 min,95%乙醇为空白对照,在波长665、649 nm下,测定吸光度,并按照ARNON的修正公式计算叶绿素含量[8],数据使用SPSS 19.0,采用方差分析进行显著性检验。
2 结果与分析
2.1 新疆野苹果诱导结果
不同浓度秋水仙素处理新疆野苹果幼苗结果见表1。0.05%秋水仙素处理存活率与対照组差异不显著,0.01%、0.10%和0.20%处理存活率显著低于对照组。存活率的变化与秋水仙素浓度的提高没有线性对应关系,但高于0.05%秋水仙素处理对新疆野苹果幼苗的存活有一定的抑制作用。
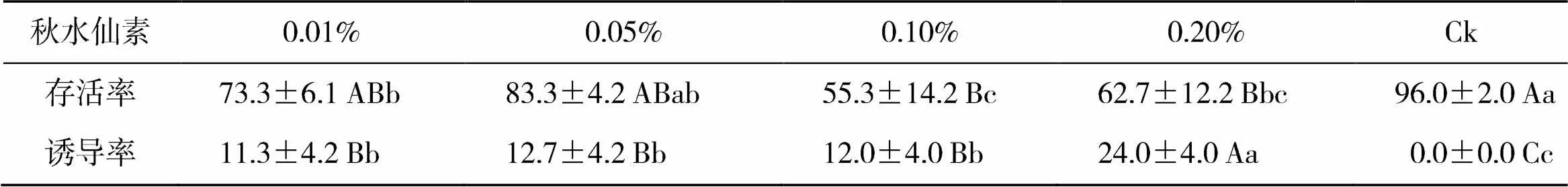
表1 秋水仙素处理结果 %
注:大写字母表示<0.01水平上不同处理间差异显著,小写字母表示<0.05水平上不同处理间差异显著
取变异植株和二倍体的茎尖制备染色体,统计细胞分裂中期细胞(图1),发现对照株染色体数为2=34,变异株的染色体数为4=68。依据染色体计数结果判定多倍体诱导结果,结合表1可知,0.20%秋水仙素诱导效果最好,而秋水仙素浓度0.01%到0.10%,多倍体诱导率变化不大。因此0.2.0%的处理浓度适合浸渍法诱导新疆野苹果四倍体。
注:A为二倍体;B为变异植株
2.2 新疆野苹果二倍体和四倍体叶片气孔及保卫细胞比较
撕取四倍体和对照的叶片表皮,测量气孔和保卫细胞的长宽如表2所示,四倍体的气孔和保卫细胞大小均极显著高于二倍体(<0.001),同时根据视野面积计算气孔密度,四倍速体气孔极显著少于二倍体。这表明变异植株叶片气孔及保卫细胞显著增大,同时气孔数量大量减少。对叶片气孔密度进行统计,发现四倍体气孔密度显著低于二倍体。

表2 变异植株与正常二倍体气孔及保卫细胞大小比较分析
注:***表示<0.001
2.3 新疆野苹果二倍体和四倍体叶片叶绿素含量比较
测定新疆野苹果二倍体和四倍体的叶片叶绿素发现,四倍体叶绿素a的含量极显著高于二倍体,计算叶绿素a/b比值和总叶绿素含量,四倍体显著高于二倍体,叶绿素b含量变化不显著(表3),这说明四倍体光合作用可能强于二倍体,营养代谢加强。
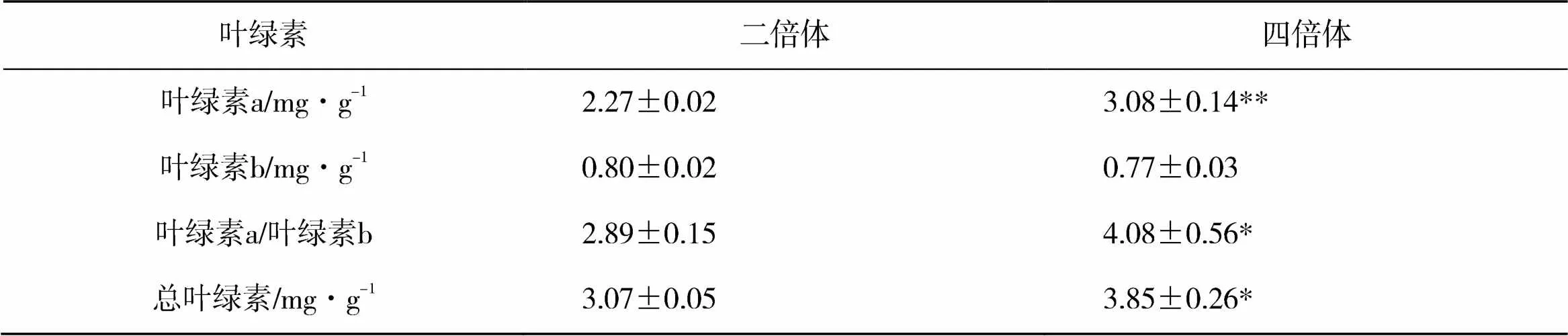
表3 新疆野苹果二倍体和四倍体叶绿素含量比较
注:*和**分别表示<0.05和<0.01
2.4 新疆野苹果二倍体和四倍体叶片解剖结构
分析新疆野苹果二倍体和四倍体叶片解剖结构发现(图2,表4),四倍体叶片厚度、叶肉厚度、上表皮和栅栏组织极显著厚于二倍体,海绵组织和下表皮显著厚于二倍体,四倍体叶片紧实度显著高于二倍体,二倍体叶片疏松度显著高于四倍体,上下表皮相对厚度变化不显著。结果说明新疆野苹果染色体加倍后,叶片结构中各部分组织不同程度的加厚,其中栅栏组织和上表皮加厚最显著,加厚导致四倍体叶片紧实度增加,疏松度降低。
注:A为二倍体;B为四倍体
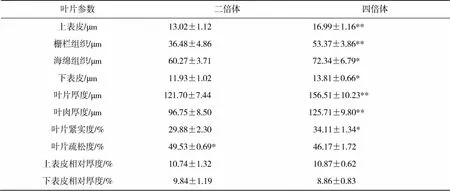
表4 新疆野苹果二倍体和四倍体叶片解剖结构比较
注:*和**分别表示<0.05和<0.01
3 讨论
苹果广泛栽种于世界各地,其适应性强,果实营养价值高。而多倍体器官巨大化的特点,使倍性育种成为果树育种中一种重要的种质创新手段。虽然苹果多倍体在苹果自然繁殖群体中可通过筛选获得,但效率和成本较高,因此人工诱导是普遍采用的倍性育种手段。其中秋水仙素是诱导多倍体效果最好的化学诱变剂,被广泛应用于苹果的多倍体诱导。但秋水仙素作为有丝分裂阻断剂,本身对植物细胞具有毒害作用,会导致发育停滞和器官畸形等问题[9]。因此选取合适的秋水仙素浓度处理植物材料,是倍性育种工作中需要摸索的试验条件。通常秋水仙素的浓度越高,对植物组织生长的抑制作用越明显。本试验在对新疆野苹果的诱导过程中,观测到了较高浓度的秋水仙素对幼苗生长有一定的抑制作用现象,但抑制作用与秋水仙素浓度的增加没有成比例增加,这可能与幼苗的个体差异有关。
秋水仙素诱导多倍体,因处理材料的不同,而采用不同浓度的秋水仙素,以往使用不定芽等组培组织作为处理材料,由于处理时间长,因此采用的秋水仙素的浓度较低,为15~120 mg/L,诱导率在10%~30%[6-7]。但培养基诱导法,对不同的苹果品种材料需要重新优化组培条件,获得组培苗还需诱导生根移植大田,步骤繁琐耗时长。本试验采用剥掉苹果种子种皮的方法快速促进苹果种子萌发,获得幼苗后浸渍处理,筛选获得多倍体,相对于培养基法较为简便,也能取得较好的诱导效果,其诱导率可达24.0%。这表明浸渍法诱导多倍体可能优于培养基法。李晓艳等对越橘采用培养基法和茎段浸渍法的试验对比结果就表明,长时间采用含低浓度秋水仙素培养基诱导的方法,其诱导效果不如短时间的浸渍法[10]。这说明秋水仙素诱导多倍体不宜处理时间太长,否则秋水仙素会抑制材料的生长。
多倍体植物一般会表现出叶片增厚变大的趋势[11],通过对新疆野苹果二倍体和四倍体的叶片解剖结构分析,四倍体叶片厚度显著高于二倍体。这与以往对多倍体苹果的观察结果一致,如马跃等发现人工诱导的‘寒富’苹果同源四倍体叶片明显加厚[12],李林光对‘乔纳金’等苹果多倍体的观察也发现多倍体叶片明显增厚[6]。这表明多倍体化影响新疆野苹果的叶片厚度发生显著变化。进一步比较分析叶片解剖结构各组织的变化发现,其中栅栏组织变厚的趋势更加显著,这种改变导致新疆野苹果四倍体的叶片紧实度显著上升而疏松度显著下降。同样的结果也出现在对‘寒富’多倍体的检测分析中,同源四倍体‘寒富’苹果的栅海比比二倍体增加了16.1%[12]。
植物叶片对环境的变化最为敏感,而植物叶片结构的变化则会改变植物对环境的适应性[13]。新疆野苹果四倍体叶片各部分增厚的同时,栅栏组织特异性的增厚导致四倍体叶片变得更加紧实。这种紧实的叶片结构被认为能赋予叶片更强的抗逆性[14]。以往研究表明,苹果属植物多倍体化后栅栏组织变厚其抗逆性提高。如‘寒富’苹果四倍体的栅海比增加,其抗盐胁迫能力提高[12,15]。苹果属其他植物多倍体的研究也表明多倍体化确实能提高其抗逆性。如珠美海棠同源四倍体对干旱、盐和缺铁胁迫有较高的抗性[16],湖北海棠四倍体具有较强的抗落叶病能力,抵御寒冷、盐和干旱胁迫的能力增强[17]。新疆野苹果四倍体栅栏组织增厚表明其抗逆性也很可能得到增强,对此有待于进一步试验验证。
在对新疆野苹果四倍体的气孔观测和叶绿素含量分析中发现,相对于二倍体,新疆野苹果四倍体叶片增厚的同时,气孔密度显著降低,同时叶绿素含量增加,这与马跃等的研究一致[12]。气孔密度降低,表明蒸腾作用可能减弱,其抗旱性可能提高,而叶绿素含量的增加则表明四倍体的光合作用可能增强,表明其积累生物量的速度加快。但试验中,获得的多倍体仅为地上部分加倍成功,获得完整四倍体植株还需经过扦插繁殖,同时由于检测深度的原因,获得的部分四倍体可能存在嵌合体的问题,还需要进一步培养后进行深入检测确定。
4 结论
本试验采用不同浓度秋水仙素浸渍新疆野苹果幼苗诱导获得四倍体,并分析四倍体的叶片解剖结构、气孔特征和叶绿素含量的变化,发现0.2%的秋水仙素处理幼苗1 d,诱导效果最佳,四倍体叶片气孔密度显著降低,叶片加厚,紧实度显著上升,疏松度下降,且叶绿素含量显著提高。该结果可为新疆野苹果的倍性育种工作提供参考和理论支持。
[1] 梅闯,闫鹏,艾沙江·买买提,等. 新疆野苹果()受苹果小吉丁虫危害程度与树皮厚度、径阶的关系[J]. 中国农业科技导报,2016,18(4):24-30.
[2] 郑点,吴玉霞,覃伟铭,等. 新疆野苹果作为苹果砧木利用的研究进展[J]. 中国野生植物资源,2019,38(2):56-59,65.
[3] 景士西. 园艺植物育种学总论[M]. 2版. 北京:中国农业出版社,2007.
[4] 马跃,宣景宏,张志宏. 苹果多倍体育种研究进展[J]. 北方果树,2015(6):1-3.
[5] 周广芳,王成强. 果树多倍体育种途径与成就[J]. 落叶果树,1997(S1):50-52,60.
[6] 李林光. 苹果多倍体种质创新及鉴定评价研究[D]. 沈阳:沈阳农业大学,2008.
[7] 刘庆忠,赵红军,刘鹏,等. 秋水仙素处理离体叶片获得皇家嘎拉苹果四倍体植株[J]. 果树学报,2001(1):7-10.
[8] 张宪政. 植物叶绿素含量测定方法比较研究[J]. 沈阳农学院学报,1985,26(4):82-84.
[9] 康萨如拉,牛建明,张庆,等. 短花针茅叶片解剖结构及与气候因子的关系[J]草业学报,2013,22(1):77-86.
[10] 李晓艳,张志东,李亚东,等. 秋水仙素诱导离体培养越橘多倍体研究[J]. 东北农业大学学报,2010,41(1):38-42.
[11] 晏春耕. 植物多倍体及其应用[J]. 生物学通报,2007(4):14-18.
[12] 马跃,张蕾,李元源,等. ‘寒富’苹果二倍体及其同源四倍体叶片超微结构和叶绿素荧光参数特征[J]. 西北植物学报,2012,32(3):477-483.
[13] 陈雪峰,景晨娟,赵习平,等. 植物叶片组织结构在抗逆研究中的应用进展[J]. 河北农业科学,2018,22(3):50-53.
[14] 马英姿,梁文斌,陈建华. 经济植物的抗寒性研究进展[J]经济林研究,2005(4):89-94.
[15] 薛浩,张锋,张志宏,等. ‘寒富’苹果与其同源四倍体耐盐差异研究[J]. 园艺学报,2015,42(5):826-832
[16] 贾少桦. 珠美海棠四倍体离体诱导、鉴定及其抗逆性研究[D]. 保定:河北农业大学,2009.
[17] 李晓林,成明昊. 四倍体湖北海棠特性研究[J]. 西南农业大学学报,1999,21(6):529-531.
Induction of polyploid(Ledeb.)Roem. and characteristic analysis of its leaves
Zhang Chenghui, Li Kaiming, Zhao Qi, Yu Jie, Gongga Yixi, Hu BaoquanCorresponding Author
(College of Horticulture and Landscape, Tianjin Agricultural University, Tianjin 300392, China)
is a rich natural resource and rootstock of apple tree. Mining its utilization method is beneficial to the further development of apple breeding. In this study, tetraploidwas induced by soaking stem apex of seedlings with different concentrations of colchicine, and changes in the characteristic structure of the leaves in tetraploidwas analyzed. The results showed that 0.2% colchicine treatment for 1 day achieved better induction effect. The leaves thickness and chlorophyll content of tetraploid increased, but stomatal density decreased, compared with diploid. Changes in anatomical structure of tetraploid leaves significantly resulted the increase of the cell tense ratio and the decrease of spongy ratio, which were was the leaf characteristics with high resistance. The results can provide a new way for the development ofgermplasm resource, new reference and theoretical support for the cultivation of new high-quality apple rootstock and polyploid breeding work of apple.
(Ledeb.)Roem.; tetraploid; colchicine; leaves; anatomical structure
1008-5394(2021)04-0006-05
10.19640/j.cnki.jtau.2021.04.002
Q944.52;S722.3
A
2020-07-08
天津市大学生创新创业训练计划项目(201810061251)
张丞慧(1998—),男,本科在读,主要从事林木倍性育种方面的研究。E-mail:18149296066@126.com。
胡宝全(1982—),男,助理研究员,博士,主要从事林木遗传育种方面的研究。E-mail:baoquan-hu@126.com。
责任编辑:杨霞
猜你喜欢
西北农业学报(2022年5期)2022-06-07
热带作物学报(2022年5期)2022-06-01
科学养鱼(2021年12期)2022-01-14
山西农业科学(2021年3期)2021-12-06
东北林业大学学报(2021年12期)2021-02-10
诊断学(理论与实践)(2020年6期)2020-12-09
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
江苏农业科学(2016年9期)2016-11-28
江苏农业科学(2014年9期)2014-11-15
