
危害乳源木莲叶片的枝膜叶蜂属(膜翅目:叶蜂科)一新种*
2022-03-22 01:34牛耕耘何学友曾丽琼魏美才
林业科学 2022年1期
牛耕耘 何学友 曾丽琼 魏美才
(1.江西师范大学生命科学学院 南昌 330022;2.福建省林业科学研究院 国家林业和草原局南方山地用材林培育重点实验室 福州 350012)
木兰科(Magnoliaceae)是一类古老的植物类群,属于被子植物的基干类群之一。该科已知约15属、150余现生种,主要分布于东南亚,中北美也有分布(吴征镒等,2003),是我国南方重要资源植物。近年来,陆续发现并报道一些叶蜂类昆虫危害木兰科树种,膜翅目(Hymenoptera)叶蜂科(Tenthredinidae)的枝膜叶蜂属(Cladiucha)昆虫专性取食木兰科植物叶片,造成严重危害。
乳源木莲(Manglietiayuyuanensis)为木兰科木莲属常绿乔木,自然分布于中国大陆东南部,包括安徽南部、浙江、江西南部、福建、湖南南部、广东北部等地,生长于海拔700~1 200 m的阔叶林中,既是优良的庭园观赏和四旁绿化树种,也是优良的建筑和家具用材(李因刚等,2008)。近年来,福建永安等地报道乳源木莲被一种未知的叶蜂严重危害,该种叶蜂具有聚集性,种群数量较大(邓林富,2017)。
目前已知的“广腰亚目(Symphyta)”昆虫有9 000余种,其中取食木兰科植物的种类非常少,已经确认的只有2科7种:茎蜂科(Cephidae)的木兰异节茎蜂HeterojanusmagnoliaeWei &Xiao,2011幼虫蛀食紫玉兰(Magnolialiliiflora)的嫩茎(Weietal.,2011);叶蜂科2属6种聚集取食木兰科植物叶片,肿跗巨基叶蜂MegabelesescrassitarsisTakeuchi,1952 取食 皱叶木兰(Magnoliakobus)和台湾含笑(Micheliacompressa)叶片(Okutani,1970),马褂木巨基叶蜂MegabelesesliriodendrovoraxXiao,1993 取食马褂木(Liriodendronchinense)的叶片(萧刚柔,1993),玉兰巨基叶蜂MegabelesesmagnoliaeWei,2010 取食玉兰(M.denudata)和紫玉兰叶片(Wei,2010),厚朴枝膜叶蜂C.magnoliaeXiao,1994 取食厚朴(M.officinalis)叶片,海南枝膜叶蜂C.manglietiaeXiao (1994)取食海南木莲(M.hainanensis)叶片(Xiao,1994;萧刚柔,1998),大鞘枝膜叶蜂C.megathecaWei,2010 取食厚朴叶片(魏美才,2010)。
枝膜叶蜂属CladiuchaKonow,1902是叶蜂科内一个非常独特的属。本属在外部形态构造上与叶蜂科平背叶蜂亚科(Allantinae)很近似,但其两性触角构型与松叶蜂科(Diprionidae)非常近似,与叶蜂科一般类群差别甚大。Benson(1938)认为本属是平背叶蜂亚科的一个独特成员,并为其建立单独的Cladiuchini族。Abe等(1991)和Taeger等(2010)均将本属归入平背叶蜂亚科,但未设立族级单元。Wei(1997)首次发现本属和巨基叶蜂属MegabelesesTakeuchi,1952(其触角构造正常)雌虫的锯背片具独特的悬膜叶,膜翅目内只有本属、巨基叶蜂属、柄基叶蜂属ConobelesesWei,1997和横脊叶蜂属TripidobelesesWei,1997共4属具此构造,在整个昆虫纲的其他类群中均未见报道。基于独特的触角和锯腹片形态构造以及聚集取食木兰科植物叶片的独特食性,Wei(1997)保留了枝膜叶蜂族,并为枝膜叶蜂族和巨基叶蜂族建立了一个新的亚科,即巨基叶蜂亚科(Megabelesinae)。Wei等(1998)将巨基叶蜂亚科放在平背叶蜂亚科和叶蜂亚科(Tenthredininae)之间。最新的基于线粒体基因组构建的系统发育研究支持巨基叶蜂亚科的单系性,并排除了巨基叶蜂亚科作为平背叶蜂亚科或叶蜂亚科内群的可能性(Niuetal.,2021)。
包括本研究记述的新种,枝膜叶蜂属全世界已经记述6种:越南枝膜叶蜂CladiuchainsolitaKonow(越南,老挝)(Konow,1902;Smith,2017),海南枝膜叶蜂(中国海南),厚朴枝膜叶蜂(中国湖南、湖北、重庆和贵州)(Xiao,1994;Wei,2010),大鞘枝膜叶蜂(中国重庆、广西和贵州)(Wei,2010),刻胸枝膜叶蜂C.punctataWei(中国云南)(Niuetal.,2020),黄氏枝膜叶蜂C.huangbkiNiu &Wei sp.nov.(中国福建)。
1 材料与方法
1.1 材料
新种模式标本采集自福建永安,保存于中国南昌亚洲叶蜂博物馆(ASCN)。
1.2 术语和缩写
构造描述术语沿用Ross(1945)和Niu等(2010)。本研究中使用的缩写有:OOL,单复眼距,指复眼内缘与邻近单眼外缘的最短距离;POL,指一对侧单眼内缘的最短间距;OCL,指侧单眼后缘至后头边缘的最短距离。
1.3 图片拍摄和处理
采用NikonD700数码相机拍摄,使用Helicon Focus(HeliconSoft®)软件进行图片合成,获得多焦点合成的成虫图像,在Photoshop® CS6软件上进行调光、去杂污点等简单处理,获得最终图像。雄虫和雌虫外生殖器图片分别使用Moticam® 5000系统和 Motic® BA400系统获得原始图像,然后在Photoshop® CS6软件上进行调光、去杂污点等简单处理,获得最终图像。采用Canon-EOS70 D相机配18~200 mm常规镜头拍摄成虫和幼虫集群的生物学照片;卵列、土中预蛹、蛹和茧的近景照片采用Canon-EOS200 D相机配EFS18~55 mm近摄镜头,加近摄接圈拍摄;卵特写照片采用KEYENCE(基恩士)-VHX-5000相机拍摄。
1.4 线粒体全基因组测序和分析
本研究共使用线粒体基因组数据7条,样品编号、GenBank登录号及采集信息等见表1。样品残体保存于中国南昌亚洲叶蜂博物馆。

表1 标本采集信息及线粒体基因组登录号Tab.1 Collecting data and GenBank accession number of mt genome sequences of sample species
使用MitoZ软件(Mengetal.,2019)对线粒体全基因组进行组装,必要时使用GENEIOUS R11软件 (http:∥www.geneious.com)(Kearseetal.,2012)对局部进行核查并重新组装。参考MITOS平台 (Berntetal.,2013)对tRNA进行注释,并根据近缘种比较研究结果对蛋白质编码基因进行注释。
1.5 种级系统树构建
选取平背叶蜂亚科2种为外群,使用IQtree软件(Trifinopoulosetal.,2016)对巨基叶蜂属2个已知种和枝膜叶蜂属3种构建最大似然树。使用translateX软件(Abascaletal.,2010)分别对13个蛋白质基因进行比对,并使用PartitionFinder软件(Lanfearetal.,2012)对比对后的串联序列进行分区。
1.6 进化速率评估
使用PAML4.9软件(Yang,2007)对各分支上所有单个数据集的非同义替换和同义替换比值(dN/dS)进行评估计算。参数如下:seqtype=1(表示序列类型是密码子);model=2(表示模型是Two-ratio model);icode=4(表示所选用密码子表是无脊椎动物mtDNA密码表),其他参数采用软件默认设置。使用EasyCodeML(Gaoetal.,2019)以上述最大似然数作为参考拓扑结构。使用CodonW计算(Enc)评估密码子使用偏倚(Nachmanetal.,1996)。
2 结果与分析
2.1 黄氏枝膜叶蜂Cladiucha huangbki Niu &Wei sp.nov.(图1—3)
雌:体长12~15 mm(图1a)。体黑色,具显著蓝紫色金属光泽;上唇白色;唇基大部蓝黑色,中部常具黄白色斑,大小有变化;腹部第1背板气门后侧小型圆斑和第7腹板中部小斑浅黄褐色;足蓝黑色,后足基节外侧长条斑、后足第1转节大部、中后足股节中基部背侧条斑、前足股节前侧大部、前中足胫跗节、后足胫节亚基部约1/3浅黄褐色,尾须褐色。翅近透明,端半部微弱烟褐色,翅痣和翅脉黑褐色。体毛银色,触角毛黑色。

图1 黄氏枝膜叶蜂新种、成虫Fig.1 Cladiucha huangbki Niu &Wei sp.nov.a.雌成虫,正模 Female adult,holotype;b.雄成虫,副模 Male adult,paratype.
体型稍粗壮。除触角黯淡无光泽外,虫体各部均具强光泽。唇基刻点粗糙,较密集(图2b);额区和单眼区刻点粗糙密集,上眶和单眼后区刻点稀疏(图2a);前胸背板刻点稍稀疏;中胸背板前叶和侧叶顶部刻点稍密集,其余部分刻点细小稀疏;小盾片中部刻点稀疏,两侧刻点稍密集;附片刻点粗糙密集,盾侧凹,底部具微细刻纹;后胸淡膜区附近刻点粗糙;中胸前侧片上部无明显刻点,下部刻点稍明显(图2e);中胸后侧片无刻点,底部具少许刻纹;后胸前侧片腹侧无刻点,背侧具浅弱刻点;后侧片上沿刻点粗密;腹部第1背板光滑,第2背板两侧和其余背板大部具浅大刻点;后足基节腹侧刻点显著。
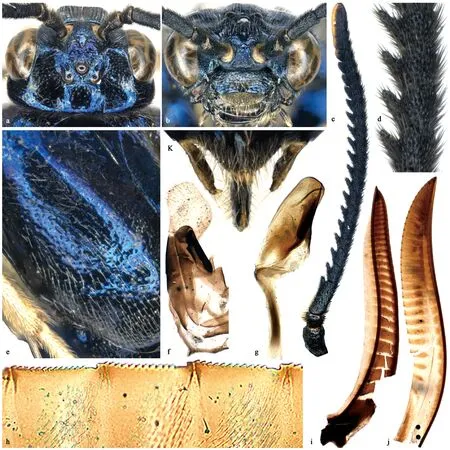
图2 黄氏枝膜叶蜂新种Fig.2 Cladiucha huangbki Niu &Wei,sp.nov.a.头部,背面观Head,dorsal view;b.头部,前面观Head,front view;c.触角Antenna;d.触角中部鞭分节Middle antennomeres;e.中胸前侧片Mesepisternum;f.生殖铗Gonoforceps;g.阳茎瓣 Penis valve;h.锯腹片中部锯刃 Middle serrulae of lancet;i.锯背片Lance;j.锯腹片Lancet;k.尾须和锯鞘背面观 Cerci and apical sheath,dorsal view;a,b,c,d,e,h,i,j,k.雌虫,正模 Female,holotype;f,g.雄虫,副模Male,paratype.
颚眼距0.4倍于侧单眼直径(图2b);触角窝上沿低弱隆起,中窝宽浅,无深窝;额脊低钝,弱隆起,中单眼前无显著隆起的纵脊;单眼中沟深,单眼后沟细,稍浅,POL∶OOL∶OCL=10∶19∶19;单眼后区稍隆起,侧面观稍低于单眼顶面,宽1.5倍于长;侧沟细弱,前部几乎消失,向后明显分歧;背面观后头长0.3倍于复眼长,两侧缘近似平行(图2a);后眶窄,后眶沟明显,后颊脊短而明显。触角22+3节,稍长于头胸部之和,明显长于头部宽的2倍,第3节微长于4、5节之和,明显短于4~6节之和,第3~17节腹侧各具1明显齿突(图2c),中部鞭分节腹侧齿突和该节侧缘突出部之间凹缺窄深(图2d),鞭节末端3节腹侧愈合。小盾片平坦,前缘短钝弧形凸出,后缘具低钝横脊,附片无中纵脊;后胸淡膜区间距约2倍于淡膜区长径。前足胫节内端距稍长于基跗节1/2长,端部分叉十分短小;后足基跗节约等于其后4个跗分节之和,基部窄细,中端部明显膨大,长宽比等于5.6∶1;爪内齿显著长于外齿。前翅R+M脉段约等于R脉长,2A+3A脉端部较弱,但通常可分辨,基臀室接近封闭;后翅Rs室封闭。尾须粗壮,长宽比约等于4,端部明显收窄(图2k);锯鞘宽大,明显长于头部宽,微短于后足股节;锯背片悬膜中部明显宽于两端,最宽处约0.7倍于该处锯背片宽(图2i);锯腹片29环节(图2j);中部锯刃平直,各具11个亚基齿,锯腹片侧面下部光裸区域较大,高三角形(图2h)。
雄:体长10~13 mm(图1b)。体色和构造类似雌虫,不同之处有:后足股节几乎全部蓝黑色,后足胫节基部白色部分短小,抱器白色;颚眼距0.3倍于侧单眼直径;侧板上部刻点稍明显;触角29+1节,第3~28节各具1对长齿突,第29节具1个齿突,端节斧状、扁平,端部钝截型;下生殖板宽大于长,端部钝截型;抱器中部明显内凸,端部倾斜钝截型(图2f);阳茎瓣头叶腹侧基部明显鼓出,中端部收窄,端部圆钝(图2g)。
词源:本种以我国著名昆虫学家黄邦侃先生姓氏命名。
变异:正模右侧后翅Rs室均不完全封闭,4雄性副模左右后翅Rs室均不完全封闭。此项变异在本属内属于正常变化。
正模:♀,福建永安燕东街道麻岭村,2019-IV-12—15,何学友。副模:2♀10,采集数据同正模。
鉴别特征:本种与大鞘枝膜叶蜂近似,但唇基白斑较小或几乎缺如;触角窝上沿微弱隆起,中窝浅平,底部无深凹,额脊低钝;单眼后区侧沟向后明显分歧;雌虫触角22+3节;小盾片平坦,中部具明显刻点;锯腹片中部锯刃几乎平直等。上述特征与后者不同。此外,2种的寄主植物也不同。
分布:福建(永安)。广东北部、浙江、江西和湖南是本种的潜在分布区。
寄主:木兰科乳源木莲(图3a)。
卵:香蕉型,尾端稍粗(图3e,f),长约2 mm,宽约0.5 mm。初卵乳白色,孵化前渐变深色。卵双列产于叶片主脉两侧叶肉内,外观卵痕明显(图3c)。
幼虫:初孵幼虫半透明,长约3 mm,宽约1 mm。老熟幼虫(图3d)粗壮,体长14.5~16.7 mm,宽3.7~4.1 mm;体黄褐色,头部、臀板大部和胸足大部黑色,酮体背侧中部暗褐色至黑褐色,边界不清晰;腹足7+1对,第6、7对腹足逐渐缩小。幼虫聚集习性显著(图3b)。
预蛹:老熟幼虫在树冠下入土做土室,在土室内越夏、越冬(图3g),土室长15~18 mm,宽6~9 mm。预蛹(图3h)体稍短缩,长13.5~16.1 mm,宽3.5~4.0 mm;腹足明显萎缩。
蛹:初蛹胸腹部淡黄色,头部和附肢乳白色,半透明(图3i),长12.9~15.9 mm,宽3.3~4.0 mm。中后期蛹色渐深,羽化前变黑色。

图3 黄氏枝膜叶蜂寄主、卵、幼虫、预蛹、蛹Fig.3 Host plant,egg,larva,prepupa and pupa of Cladiucha huangbki Niu &Wei,sp.nov.a.寄主Host plant;b.聚集危害的幼虫Larvae;c.产卵痕Spawning marks;d.幼虫Larva;e.卵列Egg row;f.卵Egg;g.幼虫入土Larva digging into soil;h.预蛹Prepupa;i.蛹Pupa.
2.2 几种枝膜叶蜂的线粒体序列比较和系统发育分析
对13个蛋白质编码基因的核苷酸序列进行P距离统计,结果如图4。在本种和刻胸枝膜叶蜂之间,有4个蛋白质编码基因(atp8、nad1、cox2、nad4L)的P距离均为0,但nad2基因的P距离为4.86%,其余9个基因的P距离介于0.56%(nad3)与0.18%(cob)之间。cox1条形码基因片段的P距离为0.30%,略低于cox1基因(1 536 bp)的P距离(0.39%)。该差异情况在膜翅目广腰蜂类中为首次报道。但昆虫近缘种间cox1基因的种间差距极小并不少见。Schmidt等(2017)统计了广腰蜂类1 126个种类共计5 360个样本的cox1条形码在种内和种间差异上的表现。其中叶蜂科涉及到6亚科853种共计4 467号样本,除Heterathrinae亚科外的5个亚科都存在cox1种间差距为0的现象,但未提到其他基因的差异情况。在枝膜叶蜂属中,cox1显然不适合单独作为条形码使用;而nad2是13个蛋白质编码基因中最小种间P距离唯一超过1%的基因,但它是否适合入选DNA条形码序列,仍需更多种类的检验。

图4 枝膜叶蜂三种之间P距离(%)Fig.4 P distance (%)of three Cladiucha species
使用线粒体基因组中13个蛋白质编码基因构建最大似然树,结果高度支持枝膜叶蜂属和巨基叶蜂属各自的单系性以及2属的姊妹群关系。
如图5所示,枝膜叶蜂属内无论是同义替换(dS)还是非同义替换(dN)均很小,这与其种间遗传距离较小是一致的。但是,枝膜叶蜂属内物种序列高度的相似性,是因为该属分化程度不高,还是因多重碱基替换或回复替换掩盖了其差异性,则需要对本属基因组进行更深入研究。

图5 巨基叶蜂亚科线粒体基因组蛋白质编码基因最大似然树及其进化速率Fig.5 Phylogenetic tree constructed with ML approach using 13PCGs sequence of Megabelesesinae and outgroups and the evaluation of substitution rates of the taxondS.同义替换Synonymous substitution;dN.非同义替换Nonsynonymous substitution;Enc.有效密码子数Effective number of codon.
7个物种的选择压力ω(ω=dN/dS)均小于1,表示这7个物种均处于净化选择之下。枝膜叶蜂属ω整体较大,巨基叶蜂属2个种的ω整体次之,黑唇十脉叶蜂和红头后室叶蜂最小,净化选择压力表现为枝膜叶蜂属<巨基叶蜂属<红头后室叶蜂<黑唇十脉叶蜂。其中刻胸枝膜叶蜂的净化选择压力最小(ω=0.223 5),黑唇十脉叶蜂的净化选择压力最大(ω=0.018 1)。有效密码子数(Enc)越小,密码子偏好性越强,进化速率普遍越小。ω与密码子的偏好性普遍成负相关性,密码子使用的偏好性越强,基因的负选择压力普遍越大。
2.3 枝膜叶蜂属分种检索表
1 唇基蓝黑色;后翅无封闭的中室(刻胸枝膜叶蜂的右侧翅具封闭M室);雌虫触角19~22+3节,明显短于头宽2倍,如果约等长于头宽2倍,则颚眼距0.6倍于侧单眼直径
2
唇基至少部分白色;后翅通常均具1个封闭的中室M(少量个体有变化);雌虫触角明显长于头宽2倍,如果短于头宽2倍,则触角仅16~17+3节,如果约等长于头宽2倍,则颚眼距0.2倍于侧单眼直径
4
2 中胸前侧片上半部无显著刻点;触角短于头宽2倍;尾须圆柱形,长宽比约等于4,端部稍尖;颚眼距0.2~0.3倍于单眼直径;小盾片无垂直后坡,刻点稀疏;锯背片悬膜腹缘平直;左侧锯背片末端背侧具6~7枚大齿;锯腹片中部锯刃显著突出,具6~8枚亚基齿
3
中胸前侧片上半部具粗大刻点;触角等长于头宽2倍;尾须粗壮,长宽比约等于2.8,端部具明显指状突;颚眼距0.6倍于单眼直径;小盾片后部刻点密集,具较短但显著的垂直后坡;锯背片悬膜腹缘明显波状弯曲,左侧锯背片末端背侧具12~13枚小齿;锯腹片中部锯刃稍突出,具9~10枚亚基齿。云南
刻胸枝膜叶蜂C.punctataWei
3 前胸背板全部蓝黑色;雌虫触角中部鞭分节侧齿狭长,几乎等长于鞭分节基部宽;后足基跗节明显膨大,长宽比小于5;前翅基臀室端部通常封闭。体长雌虫17 mm,雄虫10~11 mm。越南,老挝
越南枝膜叶蜂C.insolitaKonow
前胸背板后下侧具黄色斑纹;雌虫触角中部鞭分节端部腹侧齿突粗短三角形,长约等于鞭分节基部宽的1/2;后足基跗节细长,完全不膨大,长宽比大于6;前翅基臀室端部开放。体长雌虫14 mm,雄虫10 mm。海南
海南枝膜叶蜂C.manglietiaeXiao
4 雌虫触角16~17+3节,显著短于2倍头宽;尾须长宽比等于2;锯鞘明显短于头宽;锯腹片27环节,中部锯刃具6~7个亚基齿。重庆、湖南
厚朴枝膜叶蜂C.magnoliaeXiao
雌虫触角19~22+3节,等于或长于头宽的2倍;尾须长宽比等于3~4;锯鞘明显长于头部宽,显著长于后足基跗节;锯腹片31~32环节,中部锯刃具12~13个亚基齿
5
5 雌虫触角长约2倍于头宽;颚眼距0.25倍于侧单眼直径;中窝深,上部具1小型深坑;单眼后区侧沟向后近平行;唇基白色,仅边缘黑色;雌虫触角19~20+3节;锯腹片中部锯刃弱度倾斜。体长雌虫15~16 mm,雄虫未知。重庆,广西
大鞘枝膜叶蜂C.megathecaWei
雌虫触角明显长于头宽2倍;颚眼距0.4倍于侧单眼直径;中窝浅平,无深坑;单眼后区侧沟向后明显分歧;唇基蓝黑色,中部偶具小白斑;雌虫触角22+3节;锯腹片锯刃平直;体长雌虫12~13 mm,雄虫10~11 mm。福建
黄氏枝膜叶蜂C.huangbkiNiu &Wei,sp.nov.
猜你喜欢
植物保护(2021年4期)2021-11-12
华人时刊(2020年19期)2021-01-14
奥秘(2019年8期)2019-08-28
小猕猴学习画刊(2019年4期)2019-05-08
扬子江(2019年1期)2019-03-08
小猕猴学习画刊(2017年3期)2017-07-19
小猕猴智力画刊(2016年6期)2016-05-14
作文评点报·作文素材小学版(2016年9期)2016-03-18
百科知识(2015年7期)2015-04-08
江苏农业科学(2014年3期)2014-07-16
