
乳极性脂质组成及其功能性研究进展
2022-04-15 11:14李墨翰宋婉莹张秀敏陈佳丽热罕古丽阿卜杜拉刘爱成于海坤曹雪妍岳喜庆
乳业科学与技术 2022年2期
李墨翰,宋婉莹,张秀敏,陈佳丽,热罕古丽·阿卜杜拉,刘爱成,于海坤,张 娟,曹雪妍,*,岳喜庆,*
(1.沈阳农业大学食品学院,辽宁 沈阳 110866;2.北京食品科学研究院,北京 100068)
乳是一种含有脂质、蛋白质、碳水化合物和矿物质等成分的复杂液体,其中脂质虽然在含量上仅占乳总成分的2%~5%,但在能量供应上却占哺乳动物新生儿每日摄入的50%以上[1]。乳脂由乳腺上皮细胞分泌,其中大部分以甘油酯(非极性脂质)的形式存在于乳脂肪球内部(约98%),其余乳脂为极性脂质(约2%),则主要分布在乳脂肪球膜表面[2-3]。
甘油酯由乳腺细胞的粗面内质网合成,随后以乳糜微粒的形式积累于细胞质,并通过相互融合增大体积形成乳脂肪球,最后通过细胞质被运输到细胞两极,并被分泌到乳汁中[4-5]。乳脂肪球从乳腺细胞释放到乳的过程中,通过细胞膜时被3 层磷脂层包裹,这种3 层磷脂分子层统称为乳脂肪球膜[6-7]。乳脂肪球的这一特殊分泌过程能够将哺乳动物母体自身细胞表层的膜结合上蛋白、糖类和脂质,并最终转移到乳中,这对于哺乳动物新生儿的生长发育具有重要作用[8]。因此,本文综述不同乳源中乳极性脂质种类及数量的差异,同时探讨乳极性脂质的生理功能,以期为功能性乳脂产品的研发提供参考。
1 乳极性脂质的种类
如图1所示,根据脂质基本骨架的差异,可将脂质分成8 类,包括聚酮(polyketides,PK)、糖脂(saccharolipids,SL)、异戊烯醇脂(prenol lipids,PR)、甾醇脂(sterol lipids,ST)、鞘脂(sphingolipids,SP)、甘油磷脂(glycerophospholipids,GP)、甘油酯(glycerolipids,GL)、脂肪酸(fatty acids,FA)[9-11]。
乳中极性脂质含量较低,但其对于促进哺乳动物神经、大脑、视觉、肠道等的发育,以及保持乳脂肪球的结构具有不可或缺的作用,目前鉴定到的乳中的极性脂质主要有甘油磷脂、鞘脂和糖脂等[12-14]。
1.1 甘油磷脂
甘油磷脂由甘油、脂肪酸、磷酸及含氮化合物等组成,因其结合的磷酸具有可离解的磷酸基团,所以也是极性脂质的一种。甘油的1位和2位羟基各结合1 分子脂肪酸,3位羟基结合1 分子磷酸,该磷酸的羟基可与不同的取代基团连接,进而形成不同的甘油磷脂;如甘油磷脂酰胆碱(glycerophosphatidylcholine,PC)、甘油磷脂酰乙醇胺(glycerophosphatidylethanolamine,PE)、甘油磷脂酰甘油、甘油磷脂酸、甘油磷脂酰肌醇(glycerophosphatidylinositol,PI)、甘油磷脂酰丝氨酸(glycerophosphatidylserine,PS)、心磷脂、溶血磷脂酰胆碱、溶血磷脂酰乙醇胺等[13]。甘油磷脂含有2 条疏水的脂酰基长链,又含有极性较强、亲水的磷酸及取代基团,因此其具有疏水性和亲水性。甘油磷脂在水溶液中,其亲水部分趋向于水相,而疏水部分则互相聚集,进而形成稳定的微团或双分子层,其形成的其他常见结构还有单分子层、微囊等[15]。
1.2 鞘脂
鞘脂是一类含有鞘氨醇骨架的两性脂,一端连接1 个长链脂肪酸,另一端为1 个极性醇。鞘脂包括鞘磷脂(sphingomyelin,SM)、神经节苷脂、脑苷脂等,一般存在于植物和动物膜内,尤其是在中枢神经系统的组织内含量丰富[16]。鞘磷脂是最常见的鞘脂,其极性头是磷脂酰胆碱或磷脂酰乙醇胺,因含磷,鞘磷脂有时也被归入磷脂;脑苷脂不含磷,常呈中性,其极性头含糖;神经节苷脂是最复杂的鞘脂,含有由几个糖基组成的极性头端,其糖基至少包括1 个唾液酸。脑苷脂和神经节苷脂这样的含糖脂质也可归于糖脂一类,称鞘糖脂[17]。
1.3 糖脂
糖脂是指含有糖基配体的脂类化合物,糖脂是一类两亲性分子,在生物体内广泛存在。乳中的脂质主要是鞘糖脂和甘油糖脂。如1.2节所述,鞘糖脂亦可被归类为鞘脂类。鞘脂类分子由以下3 种结构组成:第1种结构是鞘氨醇(一种长链、带有氨基的二醇,链长约18 个碳原子);第2种结构是长链脂肪酸(链长约18~26 个碳原子),以酰胺键与鞘氨醇相结合成神经酰胺;第3种结构是极性基团的头部,通常连接在鞘氨醇第1个碳原子的羟基上。因鞘脂极性基团的差异,可将其分为不同类型。极性基团中含有磷酸则称为鞘磷脂,含有糖基则称为鞘糖脂。鞘糖脂分子中的糖基数目不等,其中含1 个糖基的鞘糖脂统称为脑苷脂[17]。
2 不同乳源乳极性脂质的差异
表1总结了包括人乳、水牛乳、牦牛乳、奶牛乳、山羊乳、绵羊乳、驴乳、骆驼乳在内的不同乳源的极性脂质含量,可以看出不同乳源极性脂质含量差异很大。尽管物种不同,但大部分乳源极性脂质中相对含量最高的均为PE,其中PE在驴乳极性脂质中相对含量最高,为60.2%,其次是绵羊乳、骆驼乳、牦牛乳、奶牛乳、水牛乳和人乳。SM在大多数乳源极性脂质中的相对含量仅次于PE,在人乳和牦牛乳中相对含量最高,而驴乳中SM相对含量最低,为8.8%。此外,不同乳源中PC相对含量也均较高,如在山羊乳中相对含量为28.5%,仅次于PE,在奶牛乳和绵羊乳中相对含量更高,分别为29.3%~33.2%和24.50%~30.50%。不同乳源的极性脂质相对含量差异显著,如水牛乳的PI相对含量在几种乳中最高,而PS最低;驴乳中PE相对含量远远超过其他几种乳,而SM、PC、PI相对含量却偏低。
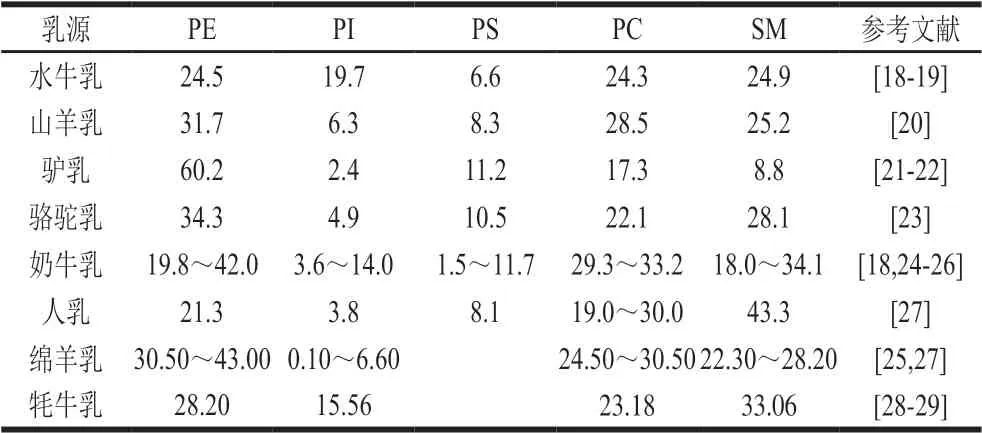
表 1 不同哺乳动物乳极性脂质亚类占总极性脂质的相对含量Table 1 Proportion of polar lipid subclasses relative to total polar lipids in different mammalian milks%
3 乳极性脂质的生理作用
3.1 抑制中性脂肪吸收
膳食中的PC通过相互作用结合成微团,降低脂肪酶和酯酶与甘油三酯(triacylglycerol,TG)的结合率,同时使蛋白质进入水相,进而抑制肠道对脂质的吸收[30]。Patton等[31]通过体外研究发现,PC分子在TG的乳液界面和本体水相之间分布,其在磷脂酶A2作用下水解为溶血卵磷脂和脂肪酸,显著减少了脂肪酶和酯酶与TG的结合,使蛋白质进入水相,进而达到抑制中性脂肪吸收的效果。Homan等[32]发现,PC在含有混合胶束的牛磺胆酸盐中明显抑制了Caco-2细胞对胆固醇的摄取,进而抑制中性脂肪酸的吸收。Rodgers等[33]的体内研究、Rampone等[34]的体外(大鼠空肠模型)研究及Greenhalgh[35]、Beil[36]等的人体实验也阐述了PC的类似作用。此外,研究表明,膳食中的PE也能够通过影响血清脂质和载脂蛋白的分布变化,进而抑制中性脂肪的吸收[37]。Imaizumi等[38]通过大鼠实验发现,膳食中连续2 周补充2%的PE可以降低血清中胆固醇的含量,同时发现胆固醇含量与肝脏PE含量呈负相关。另外,研究表明,膳食中的鞘脂能够通过疏水作用减少肠道对胆固醇、甘油酯和脂肪酸的吸收,进而抑制肠道的脂质吸收[39]。Noh等[37]通过对成年雄性大鼠进行实验,发现饲料中添加牛乳鞘脂能够显著降低大鼠对α-生育酚的吸收,从而达到抑制中性脂肪吸收的作用。同时,膳食中的鞘脂消化产物,如神经酰胺和鞘氨酸苷,也能抑制胆固醇和脂肪酸的吸收[40]。
3.2 调节肠道微生物群落组成
人类胃肠道中含有万亿个与人类生理活动、肠道功能密切相关的细菌,最为丰富的细菌分别为拟杆菌门、厚壁菌门和放线菌门[41-42]。其中,厚壁菌门与拟杆菌门相对丰度的比值与肥胖及高脂饮食相关[43]。乳极性脂质因具有一定的抗菌活性,因此可能具有调节肠道菌群的作用[44]。Fischer等[45-46]通过细胞实验及体外实验发现,鞘氨醇和脂肪酸能够通过破坏病原体的超微结构而产生抗菌活性,并且可能具有预防或治疗感染的潜力。Nejrup等[47]通过对健康婴儿粪便进行体外发酵,发现添加10%鞘氨醇的长链非酯化脂肪酸可以增加粪便中双歧杆菌的相对丰度,而单独添加长链非酯化脂肪酸则对双歧杆菌的相对丰度没有影响,说明鞘氨醇具有一定的调节肠道菌群的作用。Norris等[48]发现,膳食中添加0.25%的牛乳鞘磷脂能够增加高脂饮食小鼠粪便中双歧杆菌、厚壁菌门和放线菌门的相对丰度,从而达到调节肠道菌群的效果。随后,Millar等[49]进一步对敲除低密度脂蛋白受体的高脂饮食小鼠中添加2%的牛乳磷脂,发现双歧杆菌和拟杆菌门的相对丰度升高,同时厚壁菌门与拟杆菌门的相对丰度比值降低,说明牛乳磷脂具有调节肠道微生物、抑制肥胖的作用。大部分动物实验结果均表明,乳极性脂质对肠道微生物的群落结构有一定影响。然而,在不同的研究中其微生物群落分布存在差异,而这种差异是否与乳极性脂质的组成形式及添加量相关,仍有待进一步研究。
3.3 预防心血管疾病
心血管疾病仍然是影响全球死亡率的最主要因素之一[50],目前通过膳食调节血脂水平仍是预防心血管疾病的主要方式[51]。研究表明,乳极性脂质能够减少肠道中胆固醇、TG和脂肪酸的吸收,降低血清中的脂质水平,保护心血管健康,同时具有抑制肝肿大、防止肝脂肪变性的作用[52]。Wat等[53]也通过动物实验证明了牛乳极性脂质对于心血管疾病的预防作用,他们发现在高脂饮食小鼠饲料中添加富含牛乳磷脂的乳提取物后,小鼠的肝质量、肝脏总脂质含量、肝脏TG含量、总胆固醇含量及血脂水平均明显降低,说明其具有抑制高脂饮食小鼠肝肿大、防止肝脂肪变性及降低血清脂质水平的作用。Vors等[54]随后进行了人体实验,通过对58 名绝经后妇女进行为期4 周的双盲随机对照实验发现,牛乳磷脂组显著降低了血浆胆固醇含量与高密度脂蛋白和胆固醇的比率,同时也降低了载脂蛋白ApoB与ApoA-1的比率,进而表明牛乳磷脂能够改善心脏代谢健康,特别是通过减少肠道胆固醇的吸收来改善心脏代谢情况。总的来说,乳极性脂质对血脂和炎症的有益作用已经在动物及人体实验中被广泛证实,也是目前通过膳食预防心血管疾病的重要选择之一。
3.4 预防非酒精性脂肪肝
非酒精性脂肪肝是全世界慢性肝病最常见的病因之一,在我国具有很高的发病率[55]。研究表明,膳食中摄入鞘磷脂有助于抑制动脉粥样硬化的发展,同时通过减少中性脂质斑块体积来防止肝脏脂质的积累[56]。Yamauchi等[57]建立高脂饮食诱导的肥胖小鼠模型,并进行4 周的对照实验发现,膳食中添加牛乳鞘磷脂能够使肝脏中总脂肪酸含量减少,从而达到预防非酒精性脂肪肝的目的。Zhou等[58]也取得了相似的结果,他们发现膳食中添加牛乳极性脂质能够显著降低血浆中脂肪酸含量。此外,Millar等[49]进一步建立高脂、高胆固醇饮食的低密度脂蛋白受体敲除小鼠模型,并进行为期14 周的对照实验发现,牛乳极性脂质剂量依赖性抑制胸主动脉的动脉粥样硬化发展,特别是膳食中添加2%牛乳磷脂组显著降低了中性脂质斑块大小,进而抑制慢性肝病的发生与发展。然而,关于乳极性脂质对于人群实验是否有效,以及其是否呈剂量依赖性仍有待进一步研究。
3.5 促进认知功能与神经系统发育
包括甘油磷脂在内的一些极性脂质不仅是构成大脑和视网膜细胞的重要物质,还参与神经信号转导等过程,对大脑和神经发育至关重要[59]。Oshida等[60]通过动物实验发现,饲料中增加0.81%的牛乳鞘磷脂能够显著增加小鼠的脑髓鞘干质量;Gurnida等[61]对2~8 周龄的婴儿补充富含牛乳神经节苷脂的婴儿配方乳粉至24 周龄,发现与对照组相比,实验组婴儿的血清中神经节苷脂GM3、GD3和总神经节苷脂水平升高,证明其可能有益于神经系统发育;Tanaka等[62]对28 例早产儿进行18 个月的随机双盲实验发现,鞘磷脂强化乳组(同时添加20 g鞘磷脂和100 g甘油磷脂)婴儿的BSID-II行为评分、费根测试分数和18 个月时持续注意力测试分数等均更高。上述研究证明了乳极性脂质对于认知功能和神经系统发育的促进作用,然而牛乳与母乳对于婴幼儿神经系统发育的作用是否存在异同仍有待阐明。
3.6 抗炎作用
研究表明,乳极性脂质中的鞘磷脂及其水解产物(鞘氨醇和神经酰胺)能够改善肠屏障紧密连接蛋白的表达,进而提高肠道屏障的完整性,从而减轻高脂饮食诱导的肠道炎症,发挥抗炎作用[63]。Mazzei[64]、Norris[65]等均发现,膳食中补充牛乳鞘磷脂及其水解产物能够减少高脂饮食C57BL/6J小鼠中血清炎症标志物的含量,进而减少小鼠血清炎症细胞因子、趋化因子和炎症标记物RNA的表达,从而抑制结肠炎的发生。乳极性脂质中的多种成分对小鼠有明显抗炎作用[66-68],但在人类临床实验中的结果尚存在争议,具体作用机理及临床效果有待进一步研究。
4 结 语
乳中极性脂质虽然在乳脂中占比较少,但却对哺乳动物的生长发育有着不可或缺的作用。近年来,随着脂质组学及分子生物学技术的不断发展进步,乳中极性脂质的表征与功能性研究取得了大量成果,包括在脂质分子水平上鉴定和定量了数百种人类和其他哺乳动物的乳脂,为婴幼儿配方乳粉的研发提供了详实的理论依据与丰富的数据支持。然而,人乳和其他乳源的极性脂质在分子水平上的差异,以及这些差异对生理功能的影响,仍有待进一步研究与探索。另外,有关乳极性脂质潜在生理功能的机制研究,以及相关功能性产品的研发,仍需要进一步探索。
猜你喜欢
现代临床医学(2022年4期)2022-09-29
作物学报(2022年10期)2022-07-21
食品安全导刊(2021年21期)2021-08-30
昆明医科大学学报(2021年4期)2021-07-23
农产品加工(2021年8期)2021-05-20
老年医学研究(2021年6期)2021-03-09
健康大视野(2020年22期)2020-11-30
食品界(2019年2期)2019-03-10
大陆桥视野·下(2017年8期)2017-09-19
共产党员(辽宁)(2015年4期)2015-06-07
