
闽楠中WRKY家族成员鉴定及缺磷胁迫下表达分析*
2022-05-11 11:59周生财吴梦洁童再康张俊红程龙军
林业科学 2022年2期
张 苗 周生财 吴梦洁 童再康 韩 潇 张俊红 程龙军
(1.浙江农林大学林业与生物技术学院 省部共建亚热带森林培育国家重点实验室 杭州 311300; 2.丽水职业技术学院 丽水 323000)
磷元素(P)是植物生长发育所必需的大量元素之一,在生命大分子构成、能量转移、代谢调控和蛋白质活化等生命活动中发挥重要作用(Raghothama, 1999)。植物主要吸收土壤中的可溶性无机磷,但土壤中磷主要以非生物活性的有机磷存在,限制了植物对土壤中磷的利用。而过量施用磷肥,不但增加农作物生产成本,还会对环境造成污染(Daveetal., 2010; Vanceetal., 2003)。因此,在研究植物吸收、利用磷机制的基础上,通过现代分子育种技术,提高植物自身吸收、利用磷的效率对农作物和林木种植经济可持续发展具有重要作用。
土壤中可利用磷较少,植物在长期进化基础上形成了一系列分子水平上的适应机制(Chiouetal., 2011; Chienetal., 2018),明确植物在磷吸收、利用方面的分子机制是改善植物磷营养状况的基础。目前,在分子水平上,已有很多植物磷吸收、转运和信号转导以及植物在低磷条件下采用的分子调控策略的研究(Zhangetal., 2014; Wangetal., 2017; Haetal., 2014)。在植物适应低磷胁迫的分子调控机制中,转录因子发挥了非常重要的作用。其中,MYB类转录因子PHR1被认为是迄今为止发现的调控植物磷饥饿响应的最重要的转录调控因子,它通过调控PS1基因,影响很多磷胁迫响应相关基因的表达(Pantetal., 2015; Nilssonetal., 2007); 低磷环境下,bHLH32可以作为一个负调控因子参与调控花色素苷的合成及根毛形成过程(Chenetal., 2007); 转录因子ZAT6则参与磷相关的根系发育以及磷在植物体内的分布平衡(Devaiahetal., 2007)。
除此之外,WRKY转录因子也在植物磷饥饿胁迫中发挥重要作用。WRKY转录因子因其蛋白序列中含有1~2个DNA结合结构域WRKY(WRKYGQK)而得名。它通过识别基因启动子区域的W-box元件(C/TTGACT/C)调控相关基因表达(Eulgemetal., 2000)。除WRKY结构域外,WRKY蛋白中一般还含有1~2个其特有的锌指结构域,参与DNA序列的结合。根据其序列特征,锌指结构域可分为2种类型: C-X4-5-C-X22-23-H-X-H(C2H2型)或C-X7-C-X23-H-X-C(C2HC型)。按照WRKY蛋白所含WRKY结构域的数目及锌指结构类型将它们分为3个亚类。第Ⅰ、Ⅱ亚类所含锌指结构均为C2H2型,WRKY结构域在第Ⅰ亚类中为2个而在第Ⅱ亚类中为1个; 第Ⅲ亚类含有1个WRKY结构域,但其锌指结构类型为C2HC型(Eulgemetal., 2000)。拟南芥(Arabidopsisthaliana)的AtWRKY6、AtWRKY42、AtWRKY45、AtWRKY75和水稻(Oryzasativa)的OsWRKY74等都被证明在植物低磷胁迫下参与磷的转运、分配和信号转导,对提高植物对低磷环境的适应性发挥重要功能(Devaiahetal., 2007; Chenetal., 2009; Suetal., 2015; Daietal., 2016)。麻风树(Jatrophacurcas)的58个WRKY基因中有16个在低磷胁迫下表达发生明显变化(Xiongetal., 2013)。大豆(Glycinemax)中GmWRKY45超表达也能提高植株对低磷胁迫的适应性(Lietal., 2019)。在低磷条件下,对植物WRKY转录因子相关功能和调控机制的深入研究和分析,不但能丰富对植物磷吸收和利用分子机制的了解,还有利于挖掘植物中参与磷吸收、利用相关的WRKY基因成员并加以利用。
目前对植物中WRKY基因低磷条件下响应机制的研究还主要在几种模式植物中,其他植物中报道很少。我国南方农林用地主要以可吸收磷含量低的酸性红壤土为主,土壤中低生物活性的磷会严重限制农林植物的经济效益(于姣妲等, 2017)。针对我国南方较为广泛种植的农林植物,研究WRKY基因家族成员与低磷胁迫响应之间的关系,对利用分子育种策略改善它们的种植效益是非常有意义的探索。
楠木(Phoebe)主要在浙江、福建、江西等地分布,是中国特有的珍贵树种,材质优良,气味芬芳,既可作用材树种,生产名贵的家具,又可作观赏用的行道树。但由于其过度采伐逐渐成为濒危保护树种(Chenetal., 2020)。近年来,楠木人工林的培育开始兴起,在资源保存和利用上发挥了重要作用,但在栽培和育种上仍有许多问题需要解决。尤其是楠木不耐贫瘠,在南方低磷立地条件下种植,生长会受到严重影响(贺维, 2015)。因此,研究楠木磷吸收利用的相关分子机制,推进磷高效利用的楠木品种选育工作,对楠木产业的发展具有重要意义。
本研究以我国南方分布范围比较广的闽楠(Phoebebournei)为研究材料,在全基因组范围内鉴定了68个WRKY转录因子,分析了它们的基因结构和蛋白编码特征。基于转录组测序,对正常供磷、缺磷处理下的闽楠幼苗进行了WRKY基因家族成员的表达分析,初步鉴定了楠木中磷胁迫响应的WRKY基因,为进一步研究这些基因在磷吸收、利用分子调控途径中发挥的功能提供参考。
1 材料与方法
1.1 试验材料
闽楠半同胞家系种子采集于浙江庆元楠木种子园,沙藏后第2年春播种,6个月后,选取长势一致的植株作为试验材料。缺磷试验在植物培养箱(Snijders MC1000, 荷兰) 中进行,植株培养条件为: 白天温度25 ℃,夜间温度22 ℃; 相对湿度75%; 日照16 h/黑暗8 h; 光照强度150 μmol·m-2s-1。
1.2 PbWRKY家族基因成员鉴定
利用在线网站(http:∥pfam.xfam.org)下载WRKY转录因子保守结构域的隐马尔可夫模型(pfam03106),通过HMMER(ver 3.1)软件中hmmsearch模块在闽楠基因组蛋白序列文件中进行检索,E值设定为< e-20,去掉重复序列。对获得的PbWRKY基因进行染色体定位分析,并对PbWRKY基因进行编号、命名。
1.3 PbWRKY家族基因及其编码蛋白特性
对68个PbWRKY及拟南芥AtWRKY6、AtWRKY18、AtWRKY40、AtWRKY42、AtWRKY45、AtWRKY75和水稻OsWRKY74的蛋白序列用MEGA7.0构建1 000个重复的无根邻接(NJ)系统发育树。用在线软件GSDS2.0(http:∥gsds.cbi.pku.edu.cn)分析PbWRKY基因的外显子、内含子的数目和位置。利用Protaram(https:∥web.expasy.org/protparam/)在线网站对PbWRKY家族成员编码蛋白进行氨基酸数目、分子质量、等电点等理化特性预测。在https:∥www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi上的Batch CD-Search程序搜索PbWRKY蛋白序列中的WRKY结构域和存在的其他结构域。利用TBtools(ver.1.075)(Chenetal., 2020)作图展示。氨基酸序列通过ClustalX(ver 1.81)进行多序列比对,确定WRKY结构域和锌指结构域的位置。
1.4 PbWRKY家族基因组织特异性表达分析
以3年生闽楠半同胞家系植株为材料,选取根、韧皮部和形成层、木质部和叶片为材料提取RNA,每个样本重复3次,质检后的RNA用于RNA-seq文库构建,而后采用Illumina NovaseqTM6000 进行测序,转录组数据质量特征见表1。转录组测序由杭州联川生物技术股份有限公司完成。
1.5 缺磷处理闽楠幼苗转录组分析
试验材料分为对照组和缺磷处理组,每组各15株,每5株作为一个生物学重复。栽培方式为水培,对照组(Control,CK)采用1/4霍格兰营养液,缺磷处理组(deficiency of phosphorus, Treatment)采用去除磷元素的1/4霍格兰营养液,每3天更换1次营养液。根据前期预试验,采取缺磷处理60天作为取样时间点,分别取对照组与处理组各5株苗的根和成熟叶片,迅速液氮冷冻,作为1个重复用作RNA提取。试验重复3次。转录组测序方法同1.4,转录组数据质量特征见表1。
1.6 缺磷处理闽楠幼苗有效磷含量测定
分别取对照组与处理组各5株苗的根和成熟叶片样品各1 g,作为1个重复,将样品放入冻干机抽离水分,充分研磨后,测定有效磷含量。分别称取样品0.2 g于10 mL离心管中,加蒸馏水浸提,少量多次定容至 25 mL容量瓶中,并用0.22 μm水系滤膜过滤,溶液待测。利用间断化学分析仪BluVisionTM(Skalar,荷兰)测定磷含量。试验重复3次。
磷含量(mg·g-1)=显色液浓度(mg·L-1)×显色体积(mL)/干质量(g)×1 000。
1.7 RNA提取、cDNA逆转录和实时定量qPCR
将液氨中冷冻的样品混合均匀,充分研磨。利用RNA植物多糖多酚提取试剂盒(TIANGEN,北京)提取组织总RNA。并用反转录试剂盒PrimeScriptTMRT(Takara,北京)进行cDNA合成。实时定量qPCR试验中,用Primer3(v.0.4.0)设计引物(表2),引物合成由有康生物科技有限公司(杭州)完成。利用SYBR Premix Ex TaqTM II(Vazyme,南京),在CFX-96-well Real-Time System(Bio-Rad,美国)进行qPCR。反应体系为: SYBR Premix Ex TaqTM II 5 μL,上下游引物各0.2 μL,稀释5倍后的cDNA 0.4 μL,超纯水4.2 μL,总体积10 μL。反应程序为: 95 ℃预变性1 min; 95 ℃变性10 s,57 ℃退火持续10 s,72 ℃延伸20 s,45个循环。内参基因为PbEF1α,按照 2-ΔΔCt法计算目的基因的相对表达量。
1.8 数据处理
运用Microsoft Excel对试验数据进行整理; 试验数据的统计分析在SPSS(ver 26.0)软件上进行,并用GraphPad Prism(ver 8.0)作图。
2 结果与分析
2.1 PbWRKY基因和蛋白序列分析
通过hmmsearch搜索,最终得到68个闽楠WRKY基因。它们分别分布在12条染色体上和1条Scaffold上,呈不均匀分布。3号染色体上数量最多,有15个; 其次为5号染色体和9号染色体,分别有9个和8个; 1、2、7、8号染色体上各有5个,6和12号染色体上各有4个,4和10号染色体上分别有3个,11号染色体和Scaffold356上各有1个。根据其在染色体上的定位分别将基因命名为PbWRKY1-68(表3)。所有的PbWRKY基因均含有内含子,最少1个,PbWRKY64内含子数目最多,为28个(图1)。基因长度在1 018 bp(PbWRKY38)~39 719 bp(PbWRKY64)之间。编码氨基酸为169(PbWRKY11)~986(PbWRKY64)个。等电点在5.02(PbWRKY53)~9.81(PbWRKY39)之间。分子质量为19.09(PbWRKY11)~111.56 kDa(PbWRKY64)(表3)。

表1 组织表达与缺磷处理的转录组数据质量特征

表2 PbWRKY基因qPCR引物

表3 闽楠WRKY基因及编码蛋白特性
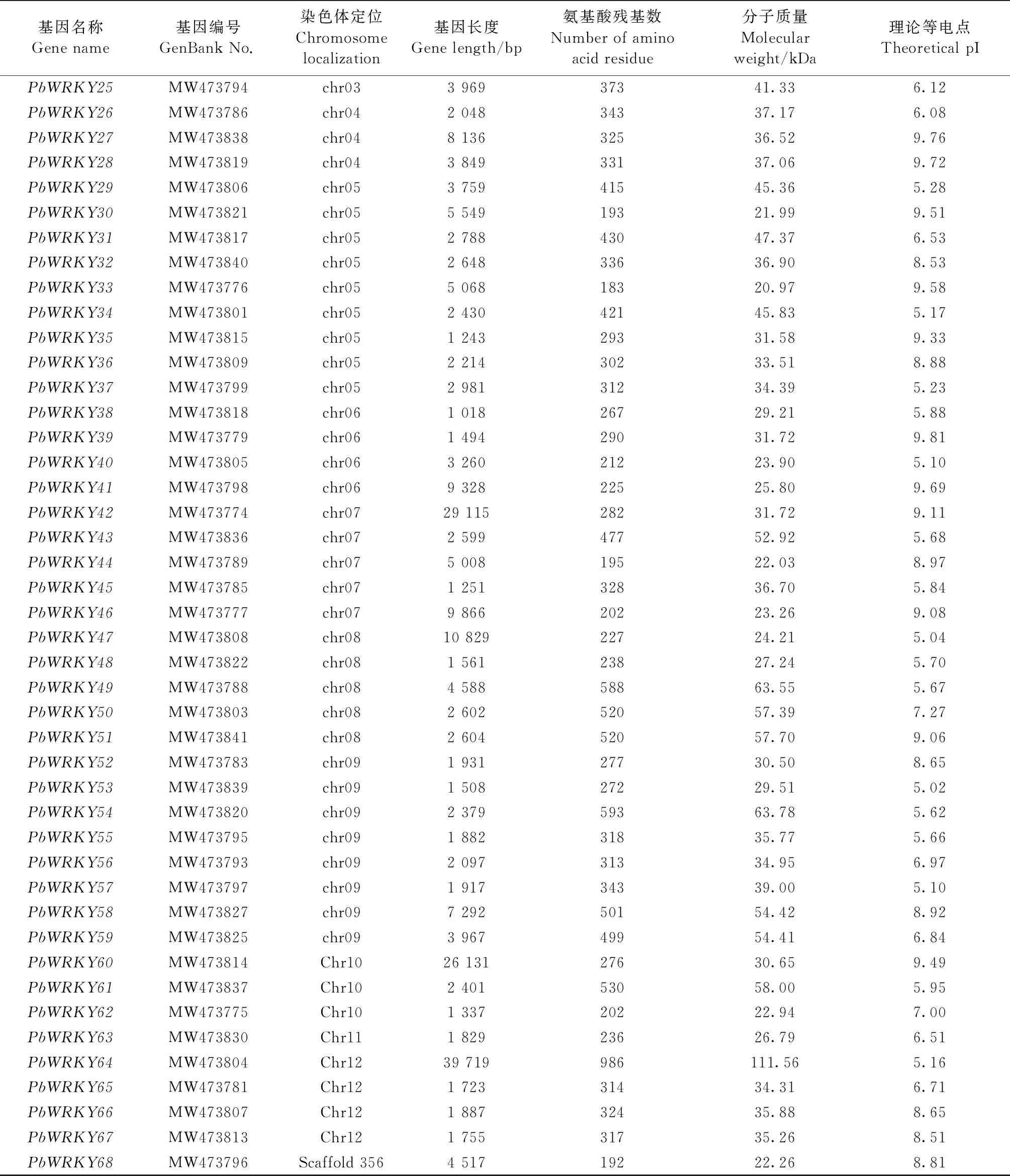
续表 Continued

图1 闽楠WRKY基因家族系统进化树、基因结构和结构域
所有PbWRKY蛋白序列均含有1或2个WRKY结构域,其核心蛋白序列为WRKYGQK; 同时含有1~2个WRKY蛋白所特有的锌指结构域,不同的WRKY基因编码的蛋白序列中含有的锌指结构域可能不同,有的为C2H2型,有的则为C2HC型(Eulgemetal., 2000)(图2)。基于蛋白序列进化树的构建及WRKY和锌指结构域的特征,68个PbWRKY可以分为I、II、III 3个亚类。其中,I亚类中含有14个PbWRKY,它们含有2个WRKY结构域和2个C2H2型锌指结构域; 第1和2条染色体上的10个PbWRKY (PbWRKY1-10)全部为第I亚类,第I亚类的另外4个成员则集中分布在第3条染色体上,暗示第I亚类是在这3条染色体上进行复制扩增的。II、III亚类中都仅含1个WRKY结构域,分别各有47个和7个PbWRKY。但II亚类中的锌指结构域与I亚类同为C2H2型,而III亚类中的锌指结构域则为C2HC型。第Ⅱ亚类中根据进化关系的不同,又可分为Ⅱa、Ⅱb、Ⅱc、Ⅱd、Ⅱe 5个子组,Ⅱa和Ⅱc分别含有6个和14个PbWRKY,其他子组都含有9个。Ⅱa和Ⅱb子组的PbWRKY蛋白序列中,除了WRKY结构域之外,还有其他结构域,如PbWRKY64的sec6结构域(pfam06046)、PbWRKY67的碱性亮氨酸拉链结构域(bZIP superfamily,cl21462)、PbWRKY58和PbWRKY59的亮氨酸拉链结构域(pfam15294)以及Ⅱd子组蛋白的锌离子结合结构域(pfam10553)等(Droge-Laseretal., 2018; Babuetal., 2006; Yazdanietal., 2020)(图1)。
2.2 PbWRKY基因组织表达特异性分析
闽楠根、韧皮部和形成层、木质部和叶片的转录组分析结果(图3)表明,68个PbWRKY基因在不同组织中的表达特征可以分为5类: 韧皮部和形成层中相对表达量较低(5个); 木质部中表达量高而叶片中表达量较低(19个); 韧皮部和形成层中表达量较高而叶片中表达量较低(10个); 根中表达量相对较高(18个); 叶片中表达量高而根中表达量低(16个)。不同PbWRKY基因组织表达特异性也有很大区别,表明它们的功能也有很大不同。

图2 闽楠WRKY蛋白序列中WRKY结构域和锌指结构类型

图3 闽楠WRKY基因不同组织部位表达热图
2.3 闽楠缺磷处理后有效磷含量的变化
由于不同植物响应低磷胁迫的情况不同,根据前期预试验,对6个月正常培养的闽楠幼苗进行60天缺磷处理。60天后,缺磷处理的植株侧根数量相对于对照明显增多,较发达,但地上部表型与对照相比没有明显差异。缺磷处理的植株叶和根中有效磷含量均显著下降(图4)。正常培养的闽楠叶片有效磷含量平均为1.235 1 mg·g-1,缺磷处理叶片有效磷平均为0.249 0 mg·g-1,降幅为79.8%; 正常培养的闽楠根中有效磷含量平均为4.349 1 mg·g-1,缺磷处理根中有效磷含量平均为0.334 9 mg·g-1,降幅为92.3%。以上结果说明此时缺磷处理的闽楠植株已经处于明显的磷饥饿状态。

图4 正常培养和缺磷处理闽楠60天后叶片(A)和根(B)中有效磷含量变化
2.4 缺磷胁迫下PbWRKY基因表达分析
缺磷处理的闽楠幼苗叶、根的转录组分析结果表明: 与对照相比,缺磷处理植株叶和根组织中表达差异达到2倍以上的PbWRKY基因共有21个,占PbWRKY基因总数目的30.9%(表4)。所有差异表达基因在缺磷处理的叶片中均被诱导,PbWRKY66诱导最强,诱导倍数达到了18.30倍。而根中表达差异达到2倍以上的基因共有5个,分别为PbWRKY52、PbWRKY55、PbWRKY56、PbWRKY65和PbWRKY66。在根中,它们在缺磷条件下表达均受到抑制。因此,这5个基因在缺磷处理60天的闽楠植株中有一个共同的表达特点,即在叶片中受缺磷诱导,而在根中受缺磷抑制。抑制最强的也是PbWRKY66,表达量仅为对照的0.091倍。
2.5 缺磷胁迫下PbWRKY基因表达的qPCR验证
为了验证转录组的可靠性,将21个差异表达的PbWRKY基因进行了qPCR验证(图5)。结果表明,这些基因的qPCR结果除了在差异表达倍数上有所浮动外,表达模式与转录组测序结果相吻合。说明转录组测序中基因表达的结果具有可靠性。在这21个基因中,有与参与拟南芥磷胁迫相关的WRKY基因相似程度很高的PbWRKY基因(图6)。如与AtWRKY6、AtWRKY42同属于Ⅱb子组的PbWRKY50、PbWRKY51、PbWRKY61等,它们都在叶片中受到强烈诱导;PbWRKY20则与AtWRKY45和AtWRKY75同属于Ⅱc子组,该基因也表现为缺磷下叶片中的诱导表达; 而与AtWRKY40和AtWRKY18氨基酸序列相似程度较高的有PbWRKY52、PbWRKY65和PbWRKY66,这3个基因的表达模式均属于缺磷胁迫下叶片中被诱导而根中被抑制。其他差异PbWRKY基因则表现为在叶片中被强烈诱导。

表4 闽楠幼苗缺磷处理60天后PbWRKY差异表达基因

图5 qPCR分析差异基因表达量

图6 PbWRKY和参与缺磷响应的AtWRKY、OsWRKY蛋白序列系统进化树
3 讨论
目前发现,大多数高等植物基因组中,WRKY基因成员数目众多。如拟南芥有72个(Eulgemetal., 2000),水稻有102个(Rossetal., 2007),毛果杨(Populustrichocarpa)有122个(Heetal., 2012),大豆中甚至达到了176个(Songetal., 2016)。本研究在闽楠中共鉴定出68个WRKY基因。这些基因编码蛋白的WRKY结构域保守性很强,仅PbWRKY62的WRKY结构域第6位氨基酸由“Q”变为“K”,这种变异会导致其结合的顺式作用元件由W-box变为WK-box(TTTTCCAC),进而引起调控基因上的差异(Verketal., 2008)。基于WRKY结构域的数量和锌指结构特点,PbWRKY蛋白也分为I、II、III 3个亚类。一般认为亚类I是进化上较为古老的一类,含有2个WRKY结构域(Zhangetal., 2005)。而数量最多的亚类II根据氨基酸序列的相似性及进化关系,又可被分为5个不同的子组,各子组间属于非单源性进化关系(Rushtonetal., 2010)。但从PbWRKY的进化关系上看,分类情况并非如此简单,如PbWRKY37尽管含有1个WRKY结构域和1个C2H2型锌指结构,但进化分析表明它不属于任何一个已知亚类II的子组。这种情况在其他植物中也有发现,如拟南芥中的AtWRKY38和AtWRKY52(Eulgemetal., 2000)。麻风树中的WRKY家族成员也有类似情况(Xiongetal., 2013)。表明PbWRKY家族成员编码蛋白序列比基于结构域的分类具有更大多样性,暗示基因功能分化也更为复杂。
PbWRKY基因组织表达特异性模式呈多样性,在一定程度上也表明了不同PbWRKY基因成员可能发挥不同功能。而且,进化上亲缘关系比较近的PbWRKY基因,组织特异性表达模式也类似。如同属于亚类I的PbWRKY2、PbWRKY3、PbWRKY7、PbWRKY18和PbWRKY19,它们都属于韧皮部和形成层表达受抑制的类型;PbWRKY20、PbWRKY32、PbWRKY33、PbWRKY41、PbWRKY46和PbWRKY68均为IIc子组中成员,它们在闽楠木质部表达量最高。由此可见,进化上亲缘关系较近的PbWRKY成员,其功能上可能具有一定的相关性。
WRKY基因在低磷胁迫中发挥着非常重要的作用。拟南芥中AtWRKY6和AtWRKY42作为负调控因子调控磷转运体PHO1基因的表达,低磷条件下被抑制表达(Chenetal., 2009; Suetal., 2015);AtWRKY45则可以通过结合在另一个磷转运体PHT1基因的启动子区域激活它的表达(Wangetal., 2014)。另外,AtWRKY18和AtWRKY40在低磷处理早期的根部也受到强烈诱导(Linetal., 2011)。水稻中超表达OsWRKY74的植株,在低磷条件下,相对于对照仍能显著提高根和地上部分的生长量(Daietal., 2016)。小麦(Triticumaestivum)中与AtWRKY75同源的TaWRKY72b-1在烟草(Nicotiana)中超表达同样能提高低磷下植株中的干质量、单株磷积累量和磷利用效率(苗鸿鹰等, 2009)。这些结果都表明,WRKY基因是参与植物磷吸收、转运途径中分子调控的一类重要转录因子。本研究中缺磷60天的闽楠植株叶片和根中,约有1/3的PbWRKY基因的表达发生变化。与拟南芥负调控因子AtWRKY6和AtWRKY42亲缘关系最近的PbWRKY50、PbWRKY51和PbWRKY61却都在叶片中受到诱导,在根中PbWRKY50、PbWRKY61的表达也有一定程度的上调(未超过2倍),这也表明这3个基因与AtWRKY6的功能有所不同。而在缺磷条件下差异表达的基因中,PbWRKY52、PbWRKY55、PbWRKY56、PbWRKY65和PbWRKY66这5个基因在根中被抑制表达的倍数超过2倍。低磷条件下,闽楠植株必然会通过基因调控作用提高根部吸收、转运磷的效率,因此,这5个基因极有可能是磷吸收转运的负调控因子,与AtWRKY6和AtWRKY42功能类似。它们在低磷条件下,通过对磷吸收转运相关基因如PHO1等的抑制,提高植物吸收、利用磷的能力; 但它们同时在叶片中又被诱导表达,诱导倍数都在5倍以上,暗示它们除了参与根部磷的吸收和转运,还可能参与了低磷条件下叶片中磷元素的再分配,其具体功能值得深入研究。另外,这5个基因中PbWRKY52、PbWRKY65和PbWRKY66在亲缘关系上与拟南芥的AtWRKY18和AtWRKY40最近,它们均属于IIa子组(图6)。但AtWRKY18和AtWRKY40分别在低磷处理1 h和2 h后,诱导水平达到最高,24 h时相对于对照已没有变化(Linetal., 2011); 而闽楠中这3个基因在低磷60天时表现出受抑制的现象,是否表明其在磷饥饿前期和后期发挥的功能不同,或者木本植物中这3个基因与拟南芥中的同源基因具有不同的功能,这些都需要进行进一步研究加以证实。PbWRKY55和PbWRKY56则与OsWRKY74一样都属于第III亚类(图6)。低磷条件下,OsWRKY74在植株叶片中被稳定诱导,而在根中低磷处理早期有比较明显的诱导,它可能参与了水稻的磷元素的吸收与平衡作用,但具体机制仍不清晰(Daietal., 2016)。因此PbWRKY55和PbWRKY56低磷条件下特殊表达模式产生的原因及基因的功能也值得深入探究。与磷吸收转运正调控因子AtWRKY45和AtWRKY75亲缘关系较近的差异表达基因为PbWRKY20,但它同样表现出叶片中被诱导、根中被抑制(抑制表达未超过2倍)的表达模式,表明它也可能在植株地上部分和根中都参与了磷的吸收和转运。
另外,其他具有显著性表达差异的15个基因都仅在叶片中被诱导表达,很明显,它们可能主要参与低磷条件下叶片磷元素的运输及再分配过程。低磷条件下,植物地上部分的光合作用,生长以及碳的代谢、分配等过程都会受到严重影响,这些过程都与磷元素的再分配有关(Hideakietal., 1991)。WRKY基因极有可能参与其中,但到目前为止,涉及低磷条件下植株地上部分磷元素的运输和再分配机制的研究很少(Wangetal., 2017)。在本研究基础上,进一步解析PbWRKY基因在植株地上部分参与的低磷响应分子机制,对楠木的耐低磷分子辅助育种工作,以及丰富木本植物适应磷胁迫的分子调控机制也有重要意义。
磷响应相关的基因在磷处理的不同时间下,表达的模式会有很大的不同,60天缺磷处理尽管使植株处于了明显磷饥饿状态下,但单一处理时间点的选择在一定程度上限制了低磷条件下WRKY基因表达分析的全面性,可能会使一些早期参与低磷胁迫响应的基因未能被发现。今后研究应该更加系统地分析低磷胁迫下WRKY基因的功能和调控机制。
4 结论
本研究在闽楠全基因组范围内,筛选鉴定出68个PbWRKY基因,命名为PbWRKY1-68,在所有染色体上都有分布。根据蛋白序列所含WRKY结构域的个数和锌指结构类型,将其分为I、II、III 3个亚类,亚类II 又分为IIa、IIb、IIc、IId、IIe 5个子组,PbWRKY37在亚类II独立分组。低磷处理60天,共有 21个PbWRKY基因在闽楠叶片和根中发生显著性差异表达,所有21个基因均在叶片中被诱导上调,其中有5个PbWRKY基因在根中被抑制,表明这21个差异基因可能参与闽楠叶和根中的磷元素吸收、转运和再分配过程。
猜你喜欢
中国交通信息化(2022年8期)2022-10-28
中国种业(2022年9期)2022-10-13
湖北农业科学(2022年11期)2022-07-18
体育科技文献通报(2022年3期)2022-05-23
特种经济动植物(2022年2期)2022-02-15
中学生数理化·高一版(2021年12期)2021-09-05
农民致富之友(2019年35期)2019-01-13
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
家教世界·创新阅读(2016年12期)2017-01-09
