
油莎豆块茎油脂积累相关基因CeWRI1的克隆与功能分析
2022-06-02 17:43徐硕邹智肖艳华张丽孔华郭静远郭安平
热带农业科学 2022年5期
徐硕 邹智 肖艳华 张丽 孔华 郭静远 郭安平


1. 海南大学热带作物学院,海南海口 570228;2. 中国热带农业科学院三亚研究院,海南三亚 572024;3. 海南省南繁生物安全与分子育种重点实验室/中国热带农业科学院热带生物技术研究所,海南海口 571101;4. 中南民族大学生命科学学院/武陵山区特色资源植物种质保护与利用湖北省重点实验室,湖北武汉 430074
摘 要:起源于非洲的油莎豆是迄今唯一已知在块茎中高水平积累油脂的新型草本油料作物。基于我国当前食用植物油和生物柴油原料供给紧张的局面,挖掘参与油莎豆块茎油脂积累的关键基因具有重要的理论意义和应用价值。WRI1(WRINKLED1)隶属于AP2/ERF转录因子家族,是一类被证实在油料种子发育过程中控制碳源由糖向油分配的关键基因。本研究采用RT-PCR技术从油莎豆的块茎中分离到一个WRI1同源基因(CeWRI1),该基因的编码区为1116 bp,预测编码371 AA,其理论分子量为41.58 kDa,等电点为5.76,总平均疏水指数为–0.750,不稳定系数为60.75,细胞核定位,这与其转录调控功能是一致的。与拟南芥WRI1类似,序列分析显示CeWRI1含有2个保守的AP2结构域(PF00847)、1个VYL基序和1个14-3-3/BPM结合基序;相比AP2结构域和N端,其C端序列的变异較大,但包含与蛋白降解相关的PEST基序。qRT-PCR分析显示,CeWRI1在叶片、叶鞘、根、匍匐茎和块茎等主要组织中都有表达,且其在起始期、膨大初期、膨大中期、膨大晚期和成熟期等不同发育时期块茎中呈现先降后升的J型表达趋势,表达丰度最高的为成熟期,最低是膨大中期,这与油脂的积累模式大体一致。在烟草中的功能分析显示,CeWRI1的异源瞬时过表达可显著提高叶片的油脂(甘油三酯)含量,这进一步证实该基因具有油脂调控功能。上述结果表明CeWRI1是调控油莎豆块茎高水平积累油脂的关键基因之一,这不仅为进一步揭示油莎豆块茎油脂积累的调控机制奠定了坚实的基础,也为后期的品种改良提供了宝贵的基因资源。
关键词:甘油三酯;油脂积累;WRINKLED1;qRT-PCR;瞬时表达中图分类号:S565.9 文献标识码:A
Cloning and Functional Characterization of CeWRI1, a Gene Involved in Oil Accumulation from Tigernut (Cyperus esculentus L.) Tubers
Abstract:Tigernut (Cyperus esculentusL.), which originates from Africa, is an herbaceous oil crop uniquely accumulating high level of oil in underground tubers. In view of the shortage of edible vegetable oil and biodiesel in China currently, it is of great significance to explore key genes involved in tuber oil accumulation.WRINKLED1(WRI1), which encodes a transcription factor belonging to the AP2/ERF family, has been proven to be a key factor controlling carbon distribution from sugar toward oil during seed development of oil crops. In this study, a gene namedCeWRI1that is homologous toArabidopsis thalianaWRI1(AtWRI1) was successfully isolated from tigernut tubers by using the reverse transcription polymerase chain reaction (RT-PCR) technique. The gene, of an open reading frame 1116 bp, was predicted to encode 371 amino acids with the theoretical molecular weight (MW) of 41.58 kDa, the isoelectric point (pI) of 5.76, the grand average of hydropathicity (GRAVY) of –0.750, and the instability index (II) of 60.75, implying its hydrophilic and instable features. Subcellular localization analysis suggests that the CeWRI1 protein is located in the nucleus, corresponding to its transcriptional regulatory function as a transcription factor. As observed in AtWRI1, sequence analyses show that CeWRI1 harbors several conserved structural features, i.e. two AP2 domains (PF00847), one VYL motif, and one 14-3-3/BPM-binding motif. Compared with the N terminus and AP2 domains, the C-terminal sequences of CeWRI1 are relatively variable, though a PEST motif associated with protein degradation was found. Quantitative real-time PCR (qRT-PCR) analysis reveals thatCeWRI1is expressed in all tested tissues, i.e. leaf blade, sheath, root, rhizome, and tuber; during various stages of developmental tuber, i.e. initial, early swelling, middle swelling, late swelling, and mature stages, a typical J-shape expression pattern was observed, peaking at the mature stage and minimizing at the middle swelling stage, which is generally consistent with the accumulation pattern of oil. Transient over-expression ofCeWRI1 in tobacco (Nicotiana tabacum) leaves significantly increased the triacylglycerol (TAG) content, supporting its role in oil accumulation. Taken together, our data suggests thatCeWRI1is most likely to be one of the key genes controlling high oil accumulation in tigernut tubers, which would not only lay a solid foundation for further uncovering the regulatory mechanism of tuber oil accumulation, but also provide a valuable resource for genetic improvement in tigernut and species beyond.6F173DBC-5E0E-4456-8EDA-520B1FEBBD37
Keywords: triacylglycerol; oil accumulation;WRINKLED1; qRT-PCR; transient expression
DOI: 10.3969/j.issn.1000-2561.2022.05.006
油莎豆(Cyperus esculentusL.),别名虎坚果,是一种隶属于禾本目莎草科的多年生草本[1-2]。生产上,油莎豆作为1年生作物栽培,以收获地下块茎为主要经济目标。油莎豆块茎的顶端含有芽点,可以像红薯、马铃薯等块根块茎类作物一样用作种豆进行无性繁殖;不同的是,油莎豆的块茎除富含淀粉(25%~45%)和可溶性糖(15%~20%)外,还积累24%~35%的油脂(甘油三酯, TAG),其中,油酸和亚油酸等不饱和脂肪酸的含量高达85%,属于保健食用油[3-6]。与大豆、油菜等传统油料作物相比,油莎豆具有适应性广、发展潜力大、每亩产油量高等特点,便于在不挤占现有耕地的情况下利用沙化边际土地增加我国的食用油供给,满足国家的战略需求[7]。然而,当前关于油莎豆的研究主要集中于栽培和产品开发[8-9],而对其遗传特性尤其是块茎油脂积累的分子机制知之甚少。
WRI1(WRINKLED1)隶属于AP2/ERF转录因子家族,是一类被证实在油料种子发育过程中调控油脂积累的关键基因[10]。WRI1基因最先在模式植物拟南芥中被鉴定,其突变体(wri1-1)的种子表现为种皮表面皱缩,种子含油量降低80%,究其原因是该突变体无法有效地将碳水化合物转化为脂肪酸合成所需的前体物质[11-12]。深入研究发现,WRI1主要通过结合顺式作用元件AW-box([CnTnG] (n)7[CG])进而调控下游基因的表达,这些基因主要参与糖酵解以及质体中脂肪酸的生物合成,如质体丙酮酸激酶-α和β亚基、丙酮酸脱氢酶E1-α亚基、生物素载体蛋白、酰基载体蛋白、β-酮脂酰-ACP合酶、烯酰-ACP还原酶1等[13-14]。至今,WRI1同源基因已在油菜、大豆、玉米、蓖麻、麻疯树、油棕等植物中得到克隆,其过表达可明显提高转基因植株的油脂含量[15-20]。2015年,GRIMBERG等[21]从油莎豆的块茎中分离到一个WRI1同源基因(CeWRI1tp),其瞬时过表达可显著提高烟草叶片的含油量,这表明该基因具有油脂调控功能。本研究组基于团队前期构建的油莎豆基因组草图发现该物种仅编码1个WRI1基因,将其命名为CeWRI1。CeWRI1的编码区与CeWRI1tp仅存在96.4%的序列一致性,推测其因品种差异所致,虽然如此,CeWRI1是否具有油脂调控功能还有待实验证实。本文重点报道CeWRI1基因及其编码蛋白的序列特征、理化特性、表达模式及其在烟草中的油脂调控功能,以期为下一步的品种改良奠定基础。
1 材料与方法
1.1 材料
1.1.1 植物材料 本氏烟草由本实验保存,种子萌发后移栽于25℃光照培养箱。供试油莎豆品系为‘热研1号,种豆催芽后播种于中国热带农业科学院文昌试验基地,组织及不同发育时期块茎等样品的采集详见邹智等[5]的方法。
1.1.2 菌株及载体 大肠杆菌DH5α和根癌农杆菌GV3101由本实验室保存;植物表达载体pNC-Cam1304-SubN由中国热带农业科学院热带生物技术研究所言普副研究员惠赠。
1.1.3 主要试剂 各类限制性内切酶、试剂盒详见肖艳华等[2]的文獻,试剂均为国产分析纯。引物合成和常规DNA测序委托北京六合华大基因科技有限公司完成。
1.2方法
1.2.1 总RNA的提取及cDNA的合成 样品总RNA的提取参照天根植物总RNA提取试剂盒说明书,经纯度、浓度和完整性检测合格后用Takara PrimeScript? RT reagent Kit with gDNA Eraser反转录试剂盒合成cDNA第一链,并将其置于–20℃保存备用。
1.2.2 基因克隆与载体构建 根据前期的文献报道从拟南芥和玉米的基因组(https://phytozome- next.jgi.doe.gov/pz/portal.html)中提取AtWRI1(AT3G54320)、AtWRI2(AT2G41710)、AtWRI3(AT1G16060)、AtWRI4(AT1G79700)、ZmWRI1a(GRMZM2G124524)、ZmWRI1b(GRMZM2G 174834)、ZmWRI2(GRMZM2G013657)和ZmWRI3(GRMZM2G131266)的基因序列。根据从油莎豆基因组中获得的CeWRI1全长转录本序列,设计如表1所示的基因特异性引物,以上述反转录的块茎cDNA作为模板,参照邹智等[22]的方法进行PCR扩增,产物经电泳检测后切胶回收目的条带,并利用同源重组的方法[2]将其克隆到pNC-Cam1304-SubN;构建的重组质粒pCam1304-6F173DBC-5E0E-4456-8EDA-520B1FEBBD37
CeWRI1采用冻融法[22]转入DH5α感受态细胞中,经菌落PCR验证后选取阳性克隆进行序列测定。
1.2.3 序列分析 采用在线软件ProtParam(https://web.expasy.org/protparam/)、SMART(http://smart.embl-heidelberg.de/)和WoLF PSORT(https://www.genscript.com/wolf-psort.html)分别分析蛋白的理化特性、保守结构域和亚细胞定位;用MEGA6.0软件进行多序列比对(MUSCLE)及进化树的构建(邻接法、bootstrap=1000)。
1.2.4 基因表达分析 参照邹智等[22]的方法进行荧光定量分析,引物信息详见表1,其中18SrRNA[2]和NtRPL23[21]分别为油莎豆和烟草的内参照基因,每个样品3次生物学重复,基因相对表达值和显著性差异分析分别用2–ΔΔCT法和SPSS软件进行。
1.2.5 烟草瞬时转化与鉴定 采用冻融法[22]将pCam1304-CeWRI1轉入GV3101感受态细胞中;参照GRIMBERG等[21]的方法,采用微量注射器将农杆菌工程菌导入6周龄烟草叶片;分别采集转化后1、3、5 d的叶片,其中一部分用于甘油三脂含量的测定,另一部分用于总RNA的提取和表达分析。
2 结果与分析
2.1基因克隆
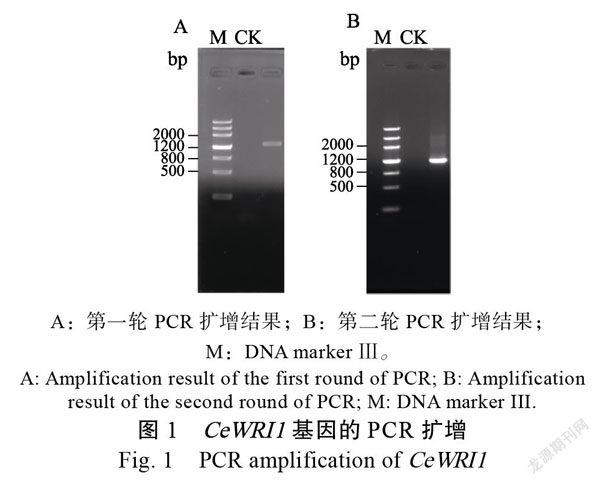
如图1所示,以CeWRI1-F/R作为引物的首轮PCR成功获得1条约1300 bp的特异条带(图1A),而以第一轮PCR产物作为模板、以CeWRI1-HF/R作为引物的第二轮PCR扩增到1条约1100 bp的目的条带(图1B);片段切胶回收后构建植物表达载体pCam1304-CeWRI1。
2.2CeWRI1生物信息学分析
测序及序列分析表明,CeWRI1的编码区(CDS)为1116 bp,略高于CeWRI1tp的1110 bp;序列比对显示其由6个碱基(AGAATG)的重复序列插入所致;此外,两序列还存在34个单核苷酸多态性(SNP)(图2)。CeWRI1预测编码371 AA,其理论分子量为41.58 kDa、等电点(pI)为5.76、总平均疏水指数(GRAVY)为–0.750、不稳定系数(II)为60.75、脂肪族指数(AI)为61.11,与CeWRI1tp相当(表2)。SMART分析和序列比对显示CeWRI1含有2个保守的AP2结构域(PF00847)、1个VYL基序和1个14-3-3/BPM结合基序;相比AP2结构域和N端,蛋白的C端序列变异较大,但与AtWRI1一样包含PEST基序(图3)。WoLF PSORT分析显示,CeWRI1定位在细胞核(表2)。
2.3CeWRI1进化分析
为进一步揭示CeWRI1的进化特征,研究将其与已报道的拟南芥和玉米同源蛋白构建了进化树。如图4所示,这些蛋白明显聚为三组,其中,CeWRI1与CeWRI1tp、AtWRI1、ZmWRI1a和ZmWRI1b聚在第一组,其序列一致性分别为96.0%、43.1%、46.1%和46.6%,暗示它们具有类似的生物学功能;在玉米中,第一组出现了扩张;第二组包括AtWRI3、AtWRI4和ZmWRI3,其在拟南芥中出现了扩张;第三组包括AtWRI2和ZmWRI2,其与前两组的亲缘关系相对较远。
2.4 CeWRI1基因表达分析
为揭示CeWRI1的表达特性,本研究首先分析了基因在叶片、叶鞘、根、匍匐茎和块茎(起始期)等主要组织中的表达模式。如图5A所示,CeWRI1在所有组织中均有微弱表达,其中在根和叶片中的表达丰度最高,其次是块茎和匍匐茎,均显著高于叶鞘。进一步对起始期(S1)、膨大初期(S2)、膨大中期(S3)、膨大晚期(S4)及成熟期(S5)等不同发育时期块茎的表达模式分析显示,基因呈现先降后升的J型表达模式,丰度最高的为成熟期,最低是膨大中期(图5B)。
2.5CeWRI1的功能分析
为证实CeWRI1在油脂积累方面的功能,本研究对烟草叶片进行了瞬时转化。qRT-PCR分析显示,转空载的对照3个时间点均未检测到CeWRI1的转录本,而实验组在转化后1 d即检测到转录,随后稳步上升,分别增加了6.6和8.4倍(图6A),这与TAG的积累模式(图6B)相似;相比基因表达,TAG的积累水平存在一定的滞后性,3 d时仅增加1.2倍,5 d时增加到2.2倍(图6B)。
3 讨论
6F173DBC-5E0E-4456-8EDA-520B1FEBBD37我国是油脂消费大国,其中,超七成的食用植物油依赖进口,严重超过国际安全警戒线,而这种供需矛盾随着社会经济的发展变得愈加突出[7]。在当今错综复杂的国际形势下,如何提升我国食用油脂的自给能力是油料科研工作者面临的一项重要课题。传统食用植物油主要来源于大豆、油菜、玉米、花生等油料作物的种子,存在产量偏低且易受气候条件影响等问题。相比而言,用块根、块茎等营养组织生产油脂更具优势及较好的互补性,然而自然情况下这些组织通常不积累油脂[23],而目前有关油脂代谢与调控的知识主要来自以拟南芥为代表的种子类植物[10]。油莎豆是迄今唯一已知在块茎中高水平积累油脂的新型油料作物[3],深入揭示其油脂积累机制具有重要理论意义和应用价值。
鉴于WRI1基因在种子油脂调控方面的关键作用[15-20],本研究从油莎豆的块茎中分离到其同源基因CeWRI1。CeWRI1与前人报道的CeWRI1tp存在较大的序列差异,其中包括六碱基的插入和数十个SNP,这不仅额外增加了2个氨基酸,同时也造成13个氨基酸的变异。虽然如此,CeWRI1和CeWRI1tp都定位在细胞核,具有相似的理化特性,并且都含有AP2结构域、VYL基序和14-3-3/ BPM结合基序等保守结构。在拟南芥中,位于第一个AP2结构域的VYL基序为AtWRI1的油脂调控功能所必须[17],14-3-3/BPM结合基序是磷酸化修饰位点,其与蛋白的稳定性密切相关[10, 24]。与AtWRI1相比,CeWRI1和CeWRI1tp蛋白的C端序列变异很大,即便如此,它们都含有类似的PEST基序,该基序被证明与蛋白的降解有关[25]。进化分析进一步证实了CeWRI1与AtWRI1的直系同源关系;相对而言,CeWRI1与第二组ZmWRI3的亲缘关系较近,而与第三组ZmWRI2较远。在玉米中,存在2个WRI1基因,即ZmWRI1a和ZmWRI1b,共线性分析显示它们来源于禾本科特异的全基因组重复,虽然它们都可以互补拟南芥wri1-4突变体,但其表达出现了明显的分化,ZmWRI1a已进化成调控油脂代谢的主效基因[16]。与CeWRI1tp[21]一样,CeWRI1在烟草中瞬时过表达可显著提高叶片的油脂含量,且该基因在块茎发育过程中的表达模式与油脂积累模式[3]大体一致,这表明CeWRI1是调控块茎高水平积累油脂的关键基因之一,虽然如此,其具体的调控机制还有待进一步研究。此外,CeWRI1也在块茎以外的其他组织(如叶片、叶鞘、根和匍匐茎)中表达,且在根和叶片中的表达水平甚至高于刚起始的块茎,暗示该基因可能参与块茎以外组织的油脂积累或者参与其他代谢通路。
综上,本研究完成了油莎豆CeWRI1基因的克隆及序列特征、进化关系、表达模式(包括主要组织和典型发育时期的块茎)和油脂调控功能的分析。这些结果不仅为进一步揭示CeWRI1调控油莎豆块茎高水平积累油脂的分子机制奠定了坚实的基础,也为今后的品种改良提供了宝贵的基因资源。
参考文献
- DE CASTRO O, GARGIULO R, DEL GUACCHIO E, CAPUTO P, DE LUCA P. A molecular survey concerning the origin of Cyperus esculentus (Cyperaceae, Poales): two sides of the same coin (weed vs. crop)[J]. Annals of Botany, 2015, 115(5): 733-745.
- 肖艳华, 邹 智, 赵永国, 郭安平, 张 丽. 油莎豆乙酰乳酸合酶基因CeALS的克隆与分析[J]. 生物技术通报, 2022, 38(4): 12-21.XIAO Y H, ZOU Z, ZHAO Y G, GUO A P, ZHANG L. Molecular cloning and characterization of an acetolactate synthase gene (CeALS) from tigernut (Cyperus esculentus L.)[J]. Biotechnology Bulletin, 2022, 38(4): 12-21. (in Chinese)
- TURESSON H, MARTTILA S, GUSTAVSSON K E, HOFVANDER P, OLSSON M E, BULOW L, STYMNE S, CARLSSON A S. Characterization of oil and starch accumulation in tubers of Cyperus esculentus var. sativus (Cyperaceae): A novel model system to study oil reserves in nonseed tissues[J]. American Journal of Botany, 2010, 97(11): 1884-1893.
- CODINA-TORRELLA I, GUAMIS B, TRUJILLO A J. Characterization and comparison of tiger nuts (Cyperus esculentus L.) from different geographical origin[J]. Industrial Crops and Products, 2015, 65: 406-414.
- 邹 智, 赵永国, 张 丽, 孔 华, 郭运玲, 郭安平. 基于单分子实时测序的油莎豆全长转录组分析[J]. 中国油料作物学报, 2021, 42(2): 229-235.ZOU Z, ZHAO Y G, ZHANG L, KONG H, GUO Y L, GUO A P. Single-molecule real-time (SMRT)-based full-length transcriptome analysis of tigernut (Cyperus esculentus L.)[J]. Chinese Journal of Oil Crop Sciences, 2021, 43(2): 229-235. (in Chinese)
- JING S, OUYANG W, REN Z, XIANG H, MA Z. The in vitro and in vivo antioxidant properties of Cyperus esculentus oil from Xinjiang, China[J]. Journal of the Science of Food and Agriculture, 2013, 93(6): 1505-1509.
- 张学昆. 我国油莎豆产业研发进展报告[J]. 中国农村科技, 2019(287): 67-69.ZHANG X K. Research progress of the tigernut industry in China[J]. China Rural Science Technology, 2019(287): 67-69. (in Chinese)
- EZEH O, GORDON M H, NIRANJAN K. Enhancing the recovery of tiger nut (Cyperus esculentus) oil by mechanical pressing: Moisture content, particle size, high pressure and enzymatic pre-treatment effects[J]. Food Chemistry, 2016, 194: 354-361.
- HU B, LI Y, SONG J, LI H, ZHOU Q, LI C, ZHANG Z, LIU Y, LIU A, ZHANG Q, LIU S, LUO Q. Oil extraction from tiger nut (Cyperus esculentus L.) using the combination of microwave-ultrasonic assisted aqueous enzymatic method-design, optimization and quality evaluation[J]. Journal of Chromatography A, 2020, 1627: 461380.
- KONG Q, YANG Y, GUO L, YUAN L, MA W. Molecular basis of plant oil biosynthesis: insights gained from studying the WRINKLED1 transcription factor[J]. Frontiers in Plant Science, 2020, 11: 24.
- FOCKS N, BENNING C. wrinkled1: A novel, low-seed-oil mutant of Arabidopsis with a deficiency in the seed-specific regulation of carbohydrate metabolism[J]. Plant Physiology, 1998, 118(1): 91-101.
- CERNAC A, BENNING C. WRINKLED1 encodes an AP2/EREB domain protein involved in the control of storage compound biosynthesis in Arabidopsis[J]. Plant Journal, 2004, 40(4): 575-585.
- BAUD S, WUILLEME S, TO A, ROCHAT C, LEPINIEC L. Role of WRINKLED1 in the transcriptional regulation of glycolytic and fatty acid biosynthetic genes in Arabidopsis[J]. Plant Journal, 2009, 60(6): 933-947.
- MAEO K, TOKUDA T, AYAME A, MITSUI N, KAWAI T, TSUKAGOSHI H, ISHIGURO S, NAKAMURA K. An AP2-type transcription factor, WRINKLED1, of Arabidopsis thaliana binds to the AW-box sequence conserved among proximal upstream regions of genes involved in fatty acid synthesis[J]. Plant Journal, 2009, 60(3): 476-487.
- LIU J, HUA W, ZHAN G, WEI F, WANG X, LIU G, WANG H. Increasing seed mass and oil content in transgenic Arabidopsis by the overexpression of wri1-like gene from Brassica napus[J]. Plant Physiology and Biochemistry, 2010, 48(1): 9-15.
- POUVREAU B, BAUD S, VERNOUD V, MORIN V, PY C, GENDROT G, PICHON J P, ROUSTER J, PAUL W, ROGOWSKY P M. Duplicate maize wrinkled1 transcription factors activate target genes involved in seed oil biosynthesis[J]. Plant Physiology, 2011, 156(2): 674-686.
- MA W, KONG Q, ARONDEL V, KILARU A, BATES P D, THROWER NA, BENNING C, OHLROGGE JB. Wrinkled1, a ubiquitous regulator in oil accumulating tissues from Arabidopsis embryos to oil palm mesocarp[J]. PLoS One, 2013, 8(7): e68887.
- JI X J, MAO X, HAO Q T, LIU B L, XUE J A, LI R Z. Splice variants of the castor WRI1 gene upregulate fatty acid and oil biosynthesis when expressed in tobacco leaves[J]. International Journal of Molecular Sciences, 2018, 19(1): 146.
- YE J, WANG C, SUN Y, QU J, MAO H, CHUA N H. Overexpression of a transcription factor increases lipid content in a woody perennial Jatropha curcas[J]. Frontiers in Plant Science, 2018, 9: 1479.
- GUO W, CHEN L, CHEN H, YANG H, YOU Q, BAO A, CHEN S, HAO Q, HUANG Y, QIU D, SHAN Z, YANG Z, YUAN S, ZHANG C, ZHANG X, JIAO Y, TRAN LP, ZHOU X, CAO D. Overexpression of GmWRI1b in soybean stably improves plant architecture and associated yield parameters, and increases total seed oil production under field conditions[J]. Plant Biotechnology Journal, 2020, 18(8): 1639-1641.
- GRIMBERG ?, CARLSSON A S, MARTTILA S, BHALERAO R, HOFVANDER P. Transcriptional transitions in Nicotiana benthamiana leaves upon induction of oil synthesis by WRINKLED1 homologs from diverse species and tissues[J]. BMC Plant Biology, 2015, 15: 192.
- 鄒 智, 郭运玲, 孔 华. 橡胶树叶片衰老相关基因HbLEA3的克隆与表达分析[J]. 西南林业大学学报(自然科学), 2021, 41(4): 42-48.ZOU Z, GUO Y L, KONG H. Cloning and expression analysis of HbLEA3, a gene associated with leaf senescence in rubber tree (Hevea brasiliensis Muell. Arg.)[J]. Journal of Southwest Forestry University (Natural Science), 2021, 41(4): 42-48. (in Chinese)
- XU C, SHANKLIN J. Triacylglycerol metabolism, function, and accumulation in plant vegetative tissues[J]. Annual Review of Plant Biology, 2016, 67: 179-206.
- ZHAI Z, LIU H, SHANKLIN J. Phosphorylation of WRINKLED1 by KIN10 results in its proteasomal degradation, providing a link between energy homeostasis and lipid biosynthesis[J]. The Plant Cell, 2017, 29(4): 871-889.
- MA W, KONG Q, GRIX M, MANTYLA J J, YANG Y, BENNING C, OHLROGGE J B. Deletion of a C-terminal intrinsically disordered region of WRINKLED1 affects its stability and enhances oil accumulation in Arabidopsis[J]. Plant Journal, 2015, 83(5): 864-874.
猜你喜欢
疯狂英语·读写版(2020年3期)2020-09-22
农村农业农民·B版(2018年3期)2018-04-09
作文周刊·高二版(2018年38期)2018-02-18
新农村(2017年14期)2017-09-30
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
智能计算机与应用(2016年4期)2016-09-26
天津农业科学(2009年1期)2009-12-28
数理化学习·高一二版(2009年3期)2009-04-30
数理化学习·高三版(2009年3期)2009-04-30
