
稳态剪切流中杆菌在粗糙表面吸附行为的有限元分析
2022-07-06 00:09丁孟辉陈巧悦刘丹枫丁明明
伊犁师范大学学报(自然科学版) 2022年1期
丁孟辉,陈巧悦,刘丹枫,丁明明,2,3*
(1.伊犁师范大学物理科学与技术学院,新疆凝聚态相变与微结构实验室,新疆伊宁 835000;2.中国科学院长春应用化学研究所高分子物理与化学国家重点实验室,吉林长春 130022;3.广东工业大学轻工化工学院,广东广州 510006)
0 引言
杆菌的吸附行为在自然界中起着至关重要的作用[1],如人类肠胃炎[2]、乳酸菌发酵[3]或微生物降解[4]等过程都与杆菌的吸附有关.在生物学和生物物理学等方面,研究杆菌在基材上的动态行为是非常重要的[5],不仅能够作为生物吸附过程的理论依据,还能够进一步促进相关生物技术的发展,如活细胞与人工材料之间界面的开发和改进[6],具有代表性的应用有生物传感器的开发和经过验证的植入材料的生产[7].
与囊泡、胶囊、细胞等模型类似,杆菌或球菌也可以被认为是一种由粘弹性膜包裹流体组成的“软颗粒”[8].目前,这种“软颗粒”和表面吸附的动态行为是分子和细胞生物动力学领域的热门研究[9],主要关注“软颗粒”膜特性和流体分布对“软颗粒”吸附和形变的影响[10],其中“软颗粒”与表面之间的吸附作用是吸附动力学的核心内容[11].1992年,Hammer和Apte首次提出了吸附动力学[12],用于模拟粘合键的吸附和解吸附过程,而粘合键的分子键用线性弹簧来代替,“软颗粒”被视为刚性粒子而忽略其形变.此后,这种方法被广泛应用于模拟“软颗粒”在剪切流作用下的吸附动力学行为[13].研究结果表明,吸附动力学主要有3种:吸附、滑移和解吸附[14].粘合键随着弹性的加强会导致吸附稳定性变弱,归因于刚性连接能够在较小扰度下承受较大作用力,最终引起恒定载荷下的快速解离.当发生滑移或吸附时,前沿新粘合键的吸附率大于后沿旧粘合键的解吸附率[15].
此外,“软颗粒”周围剪切流的作用力与粘合键吸附力之间的平衡以及力的扭矩,都对剪切流下的吸附动力学有着重要影响[16].当剪切速率低时,“软颗粒”变形较小,底部变平进而接触表面,使其稳定吸附;也就是说,小形变促进了“软颗粒”的稳定吸附.然而,当剪切速率增大时,“软颗粒”的动力学行为变得极为复杂,高形变使其底部凹陷,粘合键消失.这种特殊形状的出现加快了从稳定吸附到滑移、滑移到解吸附的过渡,意味着大形变阻碍了“软颗粒”稳定吸附[17].总体来说,增加“软颗粒”与表面之间的吸附强度会引发“软颗粒”吸附运动[18],增加流体的剪切速率会减弱“软颗粒”与表面之间的吸附强度,进而促进“软颗粒”的解吸附运动[19].
事实上,表面光滑是相对的,表面微小的凸起会影响流体分布,进而改变“软颗粒”的运动和形变.目前,很多表面修饰以及功能化[20],经常会改变表面的粗糙度,而这种微小的改变对于像细菌这种尺寸的“软颗粒”而言,很可能会改变其运动机理.到目前为止,对杆菌等“软颗粒”运动和变形的影响仍不够明确.换言之,在微米尺度下,仍缺乏杆菌在粗糙表面上受到剪切流引起的动态行为的理解.
为了系统分析稳态剪切流中杆菌在粗糙表面的动力学行为,本文使用了基于流固耦合的有限元方法,从动态角度呈现杆菌在不同粗糙表面的动力学行为,进而厘清吸附、滑移和解吸附3种运动状态与剪切率和粗糙表面特性的关系.本文第一节是流固耦合模型的具体描述;第二节是结果与讨论部分,重点阐明稳态剪切流中杆菌在粗糙表面上运动行为,并发现杆菌有3种运动模式,即吸附、滑移和解吸附,且这3种运动模式主要依赖于剪切速率和粗糙表面障碍物高度,在低剪切速率下杆菌与粗糙表面稳定吸附,在剪切速率适中时杆菌在粗糙表面滑移,而高剪切速率时杆菌与粗糙表面解吸附.
1 计算部分
1.1 模型与方法
基于我们前期关于囊泡和胶囊在剪切流中的模型[21-23],如图1a.本文在微米尺度上建立了长度L=300 μm,高度H=50 μm的二维微通道,下壁设置有正弦函数构建的高度为h和宽度为d的粗糙表面[24],且将下壁面设为固定面无速度移动,上壁面设置以ν速度向右移动,为光滑表面.由于上壁面速度大于下壁面速度产生速度差,因此在上下壁面中间的流体能够形成稳态剪切流.最后将杆菌(半长轴a=2.4 μm,半短轴b=0.6 μm,如嗜酸乳杆菌[25])放入微通道中,初始高度为y.虽然在实际中往往不均为粗糙表面,但本文理想化的粗糙面将有助于详细描述杆菌在粗糙表面的运动形态,进而形成统一且规律性的认识,从而帮助我们理解相关生理过程.
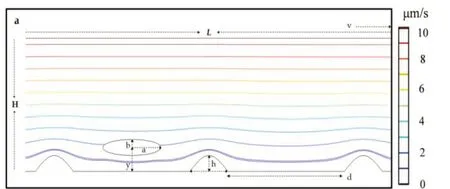
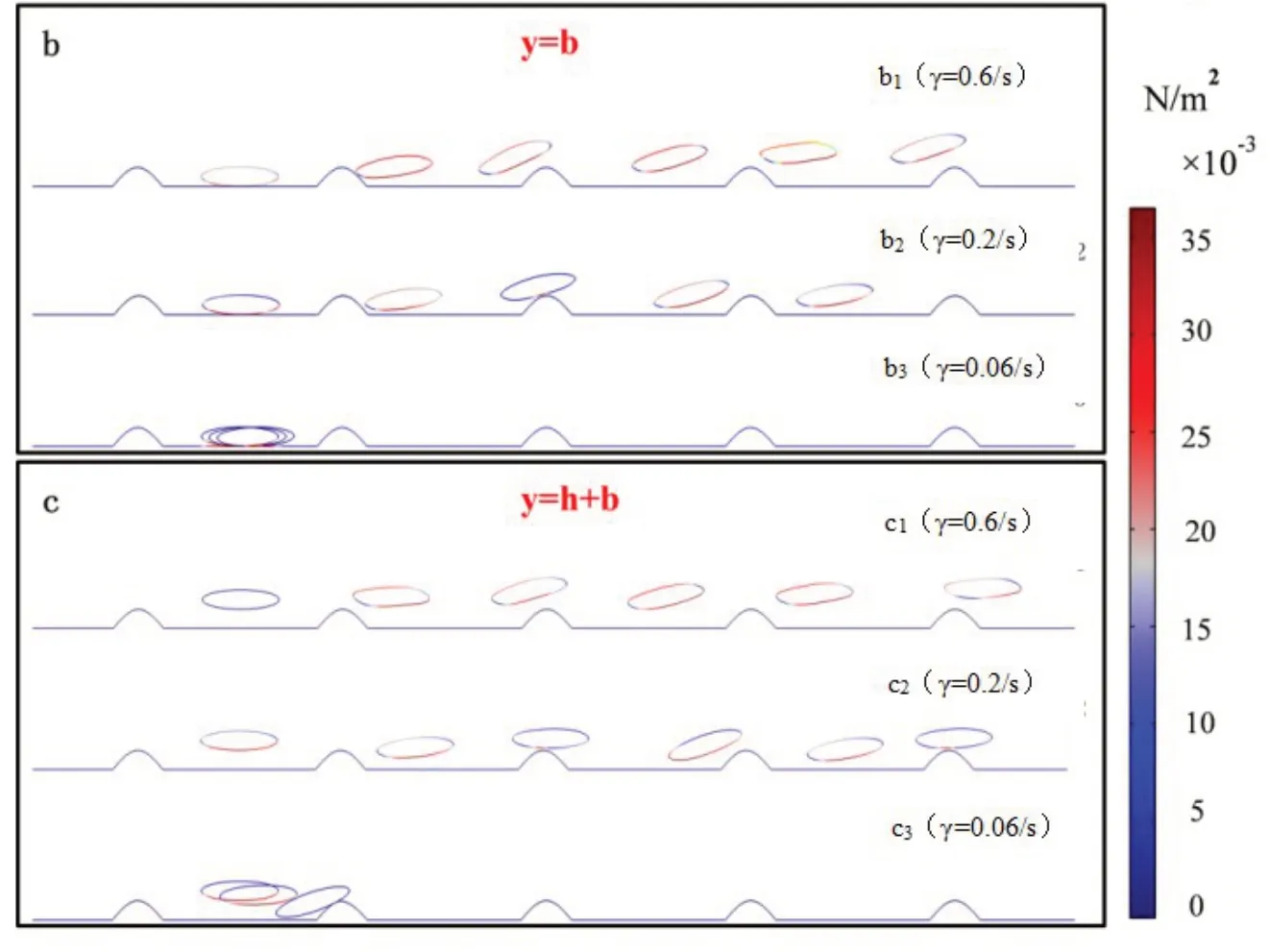
图1 (a)稳态剪切流中杆菌在粗糙表面的示意图;(b)杆菌底部与粗糙表面谷底接触(y=b)时的3种运动状态,其中y表示杆菌质心距粗糙表面谷底的距离,b表示杆菌短轴长度;(c)杆菌底部与粗糙表面障碍物高度相同(y = h+ b)时的3种运动状态,其中h表示粗糙面障碍物高度
与囊泡、胶囊、细胞等模型类似,本文假定杆菌是由粘弹性膜包裹流体组成的“软颗粒”,在流体力的作用下,杆菌的迁移和变形可描述为

其中,ρsolid是膜密度,Usolid是位移矢量,σ是应力张量,FV是体积力.利用Kelvin-Voigt模型描述膜的粘弹性行为[26],其中应力和弹性应变率的关系为

其中,η=0.022 Pa·s是膜粘度,ε是膜应变张量,由下式给出:

其中,τ是松弛时间,G是剪切模量,被定义为

其中,E=300 Pa为杨氏模量,ν=0.45是泊松比[27,28].此外,本文主要考察杆菌在重力下在粗糙表面的动力学行为,重力可表示为

其中,Fg是重力,g是重力加速度.
杆菌的运动和变形是由杆菌外流体的拖拽和膜的粘弹性以及杆菌内流体共同决定.杆菌内外的流体假定为不可压缩牛顿流体,具有相同的密度ρfluid=1 000 kg/m3和粘度μ=0.005 Pa·s,流体动力学方程如下:
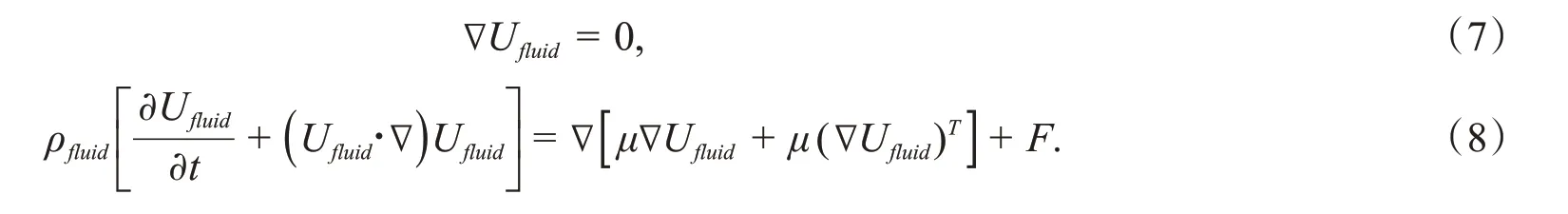
其中,F为外力,包括影响流体的其他体积力,∇对应于(∂/∂x,∂/∂y)T,Ufluid是流体的速度矢量.
流体和膜的界面处载荷和位移都相同,进而实现流固耦合,可表示为

其中,νw是膜速度,n是流膜界面的单位法向量,Γ是流体施加在膜边界上的合力,它是对流体的反作用力的负值.此外,由于本文研究中的雷诺数Re≪1,这表明流体是层流,不易形成湍流或不规则流场[29].
考虑杆菌的迁移,其水平速度可表示为

其中,usolid x和νx分别是杆菌的水平位移和速度.
本文使用COMSOL Multiphysics 5.3a模拟杆菌在粗糙表面的动力学行为,其中膜和流体流动采用了基于流固耦合(FSI)的有限元方法,并利用了非结构化网络.求解FSI 相关方程用的是任意拉格朗日欧拉(ALE)方法,即基于ALE 方法中的几何移动边界和网格滑移,计算杆菌膜的可变形网格的新坐标.除此之外,为了避免出现低质量网格影响计算精度[30,31],本文采用了自动重新划分网格.为了防止杆菌膜与粗糙表面接触引起计算结果的不收敛,本文在下壁面引入排斥区,由厚度为0.1 μm的线性弹簧组成[32,33].当膜接触到排斥区时,会被弹簧沿着边界的法线方向推开.最后,使用瞬态研究和PARDISO 求解器计算相关控制方程.
2 结果与讨论
如图1所示,我们首先考察了杆菌在不同剪切速率时,其吸附与解吸附的动力学行为.图1b为杆菌底部与粗糙表面的底部相切时的迁移规律动态演化图,即初始高度y=b.从图1b的b1中可以看出,在剪切速率较大时,杆菌表现出解吸附行为,在流体的作用下杆菌的形状发生微小改变,由椭圆形缓慢变为不规则类椭圆形,沿着粗糙表面的起伏发生滑移运动,最后逐渐脱离粗糙表面.从图1b的b2动态演化图可以看出,在特定剪切速率时杆菌表现出滑移运动状态,在流体作用下杆菌始终沿着粗糙表面的起伏发生滑移运动.从图1b的b3中可以看出,在剪切速率较小时,杆菌被困在障碍物形成的底部,形成稳定的吸附运动状态.
图1c为杆菌底部与粗糙表面的峰顶相切时的迁移规律动态演化图,即初始高度y= h+b.从图1c的c1、c2、c3可以看出,同样出现了解吸附、滑移以及吸附3种运动形态,主要差别为形成稳定吸附时(图1c的c3),不再是停在粗糙表面底部,而是形成被障碍物阻挡的形态.理论上,若杆菌在流体的驱动下受到的升力大于其自身的重力,就会逐渐脱离粗糙表面,如图1b的b1和图1c的c1所示;若升力与重力恰好处于动态平衡,就会出现类似的滑移现象,如图1b的b2和图1c的c2所示;若升力不足以克服重力的影响,就会出现逐渐靠近表面进而形成稳定吸附.实际上,在相关实验中,也观察到了类似现象,Mehdi Molaei等人通过观察大肠杆菌在各种流动剪切下的表面附近的轨迹,发现近表面流动剪切促进杆菌重新定向并增强表面分散性[34];Justin P.Jahnke等人发现大肠杆菌细胞在剪切速率低时采用弯曲几何形状,但在高剪切时采用扩展几何形状并与流动方向对齐[35];Thibaut S等人对不同的细菌进行剪切应力实验,证明剪切应力改变了细菌在基质表面的空间分布[36].因此,我们希望借助模拟,用一个具有代表性的理论模型,详细描述杆菌在粗糙表面的运动形态,进而形成统一且规律性的认识,从而帮助我们理解相关生理过程,以及如何避免杆菌在表面的沉积.
为了更细致分析与区分3种运动形态,我们计算了不同剪切速率下杆菌底部到粗糙表面底部的最小距离,如图2所示,其中ymin=1.2 μm,表示与粗糙表面障碍物顶部相切.当剪切速率γ=0.02/s 时,随着模拟的进行,杆菌底部到粗糙表面底部的最小距离迅速减小并直至趋于0,形成稳定吸附状态.从计算的ymin值可以看到,杆菌没有跨过障碍物,而是直接降到了粗糙表面底部.当剪切速率γ=0.1/s时,除初始位置外,杆菌在运动过程中,形成了2个峰值,峰值大小约为1.2 μm.这表明杆菌在形成吸附前,2次跨过了障碍物,然后才形成吸附形态.这也表明,形成吸附形态的过程与剪切速率密切相关.当剪切速率γ=0.2/s时,杆菌的运动形式表现出周期性变化,我们认为此时杆菌为滑移运动,也就是杆菌随着障碍物的起伏做有规律的上下迁移.当剪切速率γ=0.4/s,γ=0.6/s或γ=1/s时,随着模拟的进行,杆菌底部与粗糙表面底部的距离逐渐变大,成为解吸附运动.但我们发现,即使杆菌底部已经远离障碍物顶部,其在y方向仍然做着振荡运动,只是振荡幅度逐渐减小,如γ=1/s曲线所示.这主要是因为障碍物对其附近的流场有着重要影响,流场随障碍物的变化,也进一步影响了杆菌的运动方式,只是随着杆菌离障碍物越来越远,这个影响逐渐减弱.我们的计算结果表明,表面的粗糙程度对杆菌的迁移规律有着重要影响,当表面粗糙度难以参数化表征时,其周围的流场分布将会变得极其复杂,进而对杆菌形成很多不明确的影响.理想化的粗糙表面,对我们理解这个现象以及该现象背后的规律,有着重要帮助.
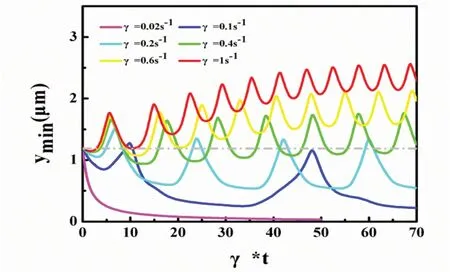
图2 不同剪切速率下杆菌底部到粗糙表面底部的最小距离
我们进一步研究不同初始角度对杆菌3种运动形态的影响,如图3所示,其中(a—d)图分别为初始角度θ=0°,θ=30°,θ=45°和θ=90°的情况,其中θ定义为杆菌长轴与剪切场方向的夹角,ymin=1.2 μm表示杆菌底部恰好与粗糙表面障碍物顶部相切.
综合图3我们可以发现,当剪切速率γ=0.06/s时,随着模拟的进行,杆菌底部到粗糙表面底部的最小距离缓慢减小并直至趋于0,形成稳定的吸附状态.从计算的ymin值可以看到,杆菌没有跨过障碍物,而是直接降到了粗糙表面底部.值得注意的是,随着初始角度的增加,ymin值下降速率加快,这主要归因于此时杆菌顶部与底部受到的剪切力差值较大.
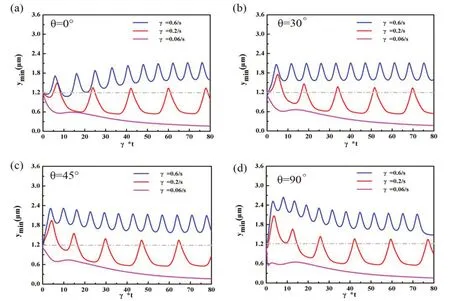
图3 不同初始角度对杆菌底部到粗糙表面底部的最小距离的影响,其中(a)表示初始角度θ=0°时,剪切速率为0.6/s、0.2/s和0.06/s时分别对应的解吸附、滑移和吸附运动;(b)表示初始角度θ=30°时,对解吸附、滑移和吸附运动的影响;(c)表示初始角度θ=45°时,对解吸附、滑移和吸附运动的影响;(d)表示初始角度θ=90°时,对解吸附、滑移和吸附运动的影响
剪切速率γ=0.2/s时,除初始位置外,杆菌在运动过程中,在初始角度θ为0°、30°、45°、90°时峰值的数量分别为5个、5个、5个、6个.从峰值大小分析可知,随着初始角度的增加,第一个周期性变化的峰值逐渐增大,峰值最后稳定在1.2 μm以上,且形成周期的时间变短,即杆菌随着障碍物的起伏做有规律的上下迁移.
当剪切速率γ=0.6/s时,随着初始角度的增加,杆菌底部与粗糙表面底部的距离逐渐增大,形成解吸附运动.除初始位置外,杆菌在运动过程中,在初始角度θ为0°、30°、45°、90°时峰值的数量分别为10个、11个、12个、12个.从峰值大小分析可知,随着初始角度的增加,第一个周期性变化的峰值逐渐增大,且形成周期的时间变短,即杆菌在y方向振荡的频率增加.总之,我们发现不同初始角度对杆菌在不同剪切速率下的运动形态影响较小.
为了进一步研究杆菌的运动形态,我们计算了剪切速率和粗糙表面高度的协同影响,如图4所示.其中(a)为杆菌底部与粗糙表面底部相切,即初始位置y=b的情况,(b)为杆菌底部与粗糙表面障碍物顶部相切,即初始位置y= h+b的情况.图4(a)中,当粗糙表面高度在0 <h<0.5b范围内时,随着剪切速率的逐渐增加,杆菌的运动形式从吸附直接转变为解吸附的状态,没有观察到滑移行为,这表明此时杆菌更容易实现解吸附;当粗糙表面高度在0.5b≤h≤3b范围内时,随着剪切速率的逐渐增加,杆菌运动形式从与粗糙表面稳定吸附先转变为表面滑移后转变成解吸附的状态,这表明此时杆菌受障碍物高度和剪切速率的协同影响显著;当粗糙表面高度h>3b的情况时,杆菌的运动模式只存在一种形式,即稳定吸附,此时障碍物的高度已经可以完全屏蔽剪切流场的影响.粗糙表面高度越高,意味着粗糙表面附近的流体速度越小,高剪切速率也无法改变这一趋势,使得杆菌在粗糙表面底部很难受到流体的升力.因此容易与壁面牢固粘附.
图4(b)中,当粗糙表面高度在0 <h<0.5b范围内时,随着剪切速率的逐渐增加,杆菌运动形式从吸附直接转变为解吸附的状态,与(a)图的结果一致,这表明当障碍物高度小于杆菌短轴距离的一半时,无论杆菌初始位置如何变化,其运动形态主要受剪切场的影响;当粗糙表面高度在0.5≤h≤3b范围内时,随着剪切速率的逐渐增加,杆菌运动形式从与粗糙表面稳定吸附先转变在表面滑移后转变成解吸附的状态,这与(a)图结果类似,也表明此时杆菌受障碍物高度与剪切速率的协同影响显著;当粗糙表面高度h>3b时,随着剪切速率的逐渐增加,杆菌运动从与壁面吸附直接转变为在壁面滑移的状态,此时与(a)图差异最大,也就是说,无论障碍物多高,只要杆菌初始位置不在障碍物形成的波谷里面,较大的剪切速率就可以避免杆菌的吸附.
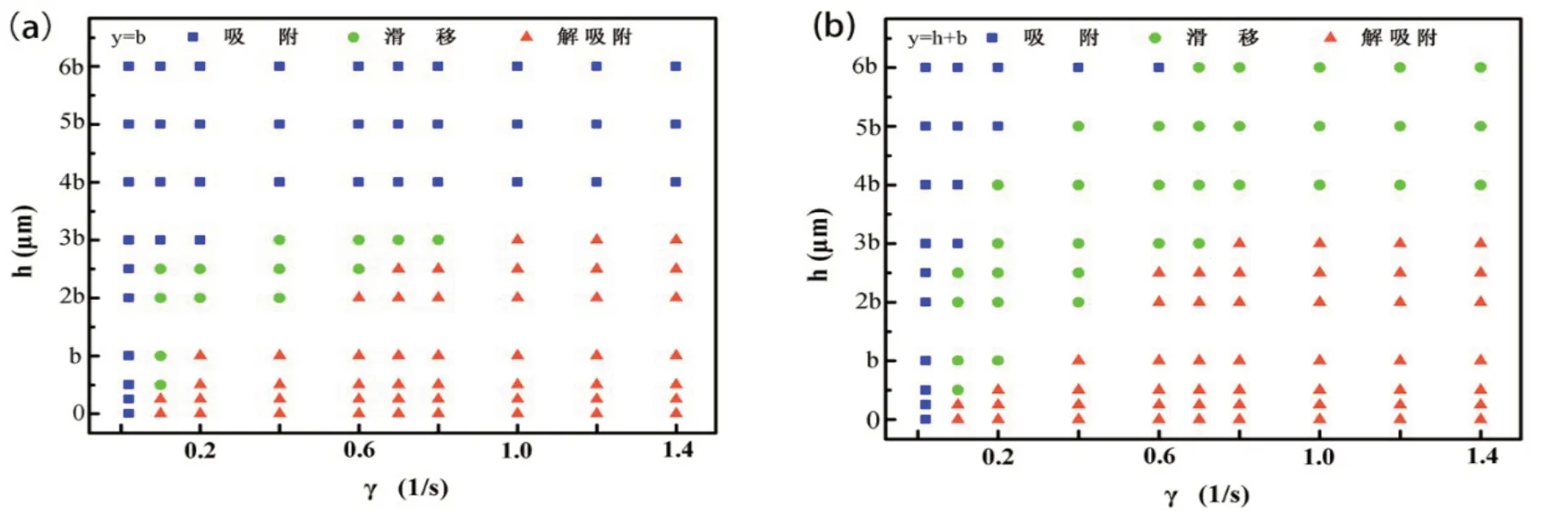
图4 不同初始位置条件下杆菌运动形态的动力学相图,其中(a)表示初始位置为y=b时,在不同粗糙表面高度下随着剪切速率的增加对应的不同运动状态;(b)表示初始位置为y=h+b时,在不同粗糙表面高度下随着剪切速率的增加对应的不同运动状态
我们的模拟结果表明,杆菌的初始位置对其运动方式有较大影响,当障碍物高度小于短轴距离的一半时,可以认为此时表面相对光滑,杆菌的运动形式主要受剪切速率的影响;当障碍物高度与短轴距离相当时,此时杆菌的运动形式最复杂,随着剪切速率的增大,先后出现吸附、滑移和解吸附行为,此时障碍物高度与剪切速率的协同影响最显著;当障碍物高度大于短轴距离的1.5倍时,此时障碍物的影响逐渐占据主导,不会出现解吸附行为.
我们定义临界滑移剪切速率为刚好可以使杆菌从吸附状态转变到滑移状态的最小剪切速率.从图5中可知,当0.5b≤h≤2.5b时,随着初始角度的增加,杆菌的临界滑移剪切速率不变,即γ=0.1/s.当3b≤h≤4b时,杆菌滑移剪切速率在γ=0.2/s 保持不变.当h=5b时,杆菌的滑移剪切速率不随初始角度的增加而改变,即γ=0.4/s.当h=6b时,初始角度在0°≤θ≤60°范围内时,杆菌的滑移剪切速率保持不变,即γ=0.7/s;初始角度在60°<θ≤90°范围内时,杆菌的滑移剪切速率逐渐增加至γ=0.9/s;初始角度在90°<θ≤120°范围内时,杆菌的滑移剪切速率逐渐减小至γ=0.6/s;初始角度在120°<θ≤135°范围内时,杆菌的滑移剪切速率逐渐增加至γ=0.7/s后保持不变.当h=8b时,初始角度在0°≤θ≤45°范围内时,杆菌的滑移剪切速率保持不变,即γ=0.8/s;初始角度在45°<θ≤90°范围时,杆菌的滑移剪切速率先逐渐增加至γ=1/s,后逐渐减小至γ=0.8/s;初始角度在90°<θ≤135°范围内时,杆菌的滑移剪切速率逐渐保持不变,即γ=0.8/s;初始角度在135°<θ≤150°范围时,杆菌的滑移剪切速率逐渐减小至γ=0.7/s.当h=10b时,杆菌的滑移剪切速率保持不变,即γ=1/s.我们的计算结果表明,只有高障碍物时,大的初始角度才会对杆菌的迁移规律有一定的影响.

图5 障碍物高度不同时临界滑移剪切速率相图,其中障碍物高度h从0.5b到10b,杆菌初始位置为y = h+ b
从图6可知,粗糙表面障碍物密度在0<d/b<8范围内时,随着剪切速率的逐渐增加,杆菌运动形式从与粗糙表面稳定吸附先转变在表面滑移后转变成解吸附状态;粗糙表面密度在d/b≥8时范围内时,随着剪切速率的逐渐增加,杆菌运动形式从与粗糙表面稳定吸附直接转变成滑移状态.显然,粗糙表面障碍物密度能够影响杆菌的运动方式.粗糙表面障碍物密度越大,意味着粗糙表面附近的流体阻力越大,杆菌的运动形式受影响越大.当粗糙表面障碍物密度较低时,此时流场在表面分布相对均匀,导致杆菌一直处于类滑移状态,这也是实验中最常见的运动形式[34-36].
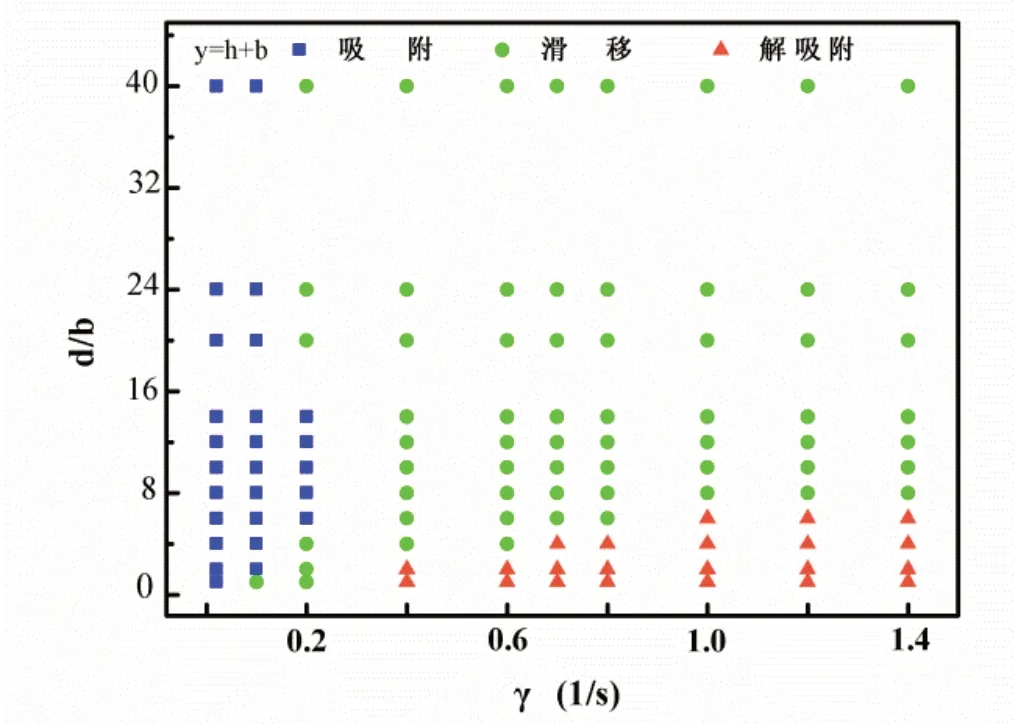
图6 考虑障碍物密度的杆菌运动形态的动力学相图,其中杆菌初始位置y = h+ b
3 结论
本文使用有限元的方法定性研究了稳态剪切流中杆菌在粗糙表面上的动力学行为,模拟发现障碍物高度和剪切速率对杆菌的运动形式有较大影响,而初始位置、初始角度以及障碍物密度等均有不同程度的影响.主要发现:当障碍物高度小于杆菌短轴距离的一半时,可以认为此时表面相对光滑,杆菌的运动形式主要受剪切速率的影响;当障碍物高度与杆菌短轴距离相当时,此时杆菌的运动形式最复杂,随着剪切速率的增大,先后出现吸附、滑移和解吸附行为,此时障碍物高度与剪切速率的协同影响最显著;当障碍物高度大于杆菌短轴距离的1.5倍时,此时障碍物的影响逐渐占据主导地位,不会出现解吸附行为.希望通过本次研究,进一步理解和认识杆菌在粗糙表面的动力学行为,为其在医疗抗菌、靶向药物研发、微生物环境修复和工业制造等领域中的应用提供可靠的支撑.
猜你喜欢
发明与创新(2022年28期)2022-11-22
力学与实践(2022年3期)2022-07-02
能源工程(2022年2期)2022-05-23
大众科学(2020年7期)2020-10-26
航空发动机(2020年3期)2020-07-24
动漫界·幼教365(中班)(2020年3期)2020-04-20
创新作文(1-2年级)(2019年4期)2019-10-15
好孩子画报(2019年10期)2019-01-10
筑路机械与施工机械化(2015年1期)2015-09-18
地震研究(2014年3期)2014-02-27
