
沟叶结缕草ZmCOR410基因抗逆功能鉴定
2022-09-14 04:22张晓寒陈彦良张珊珊韦善君
华北农学报 2022年4期
张晓寒,陈彦良,马 欣,张珊珊,韦善君
(中央民族大学 生命与环境科学学院,北京 100081)
结缕草属(Zoysia)植物是C4类暖季型禾本科草,约有10 个种,具有发达的匍匐茎和地下根茎,是建植草坪草的良好材料[1-2]。目前,在全球广泛种植的有日本结缕草(Z.japonicaSteud.)、沟叶结缕草(Zoysiamatrella)、细叶结缕草(Z.tenuifolia)、中华结缕草(Z.sinicaHance)以及它们之间的杂交种,它们形成的草坪耐高温[3]、干旱[4]、高盐[5]和重金属[6-7]胁迫,耐践踏[8],养护成本低。在我国结缕草地理分布广泛,种质资源丰富。其中,山东半岛和辽东半岛有大面积的野生结缕草资源,是我国结缕草种子生产的重要基地[9-10]。在我国草坪草业界,结缕草一直占据着重要的地位,首要原因为结缕草是我国唯一出口种子的草种,其次是我国适合种植结缕草的面积广泛。沟叶结缕草及其衍生种地上茎密集,叶较硬且直立,匍匐茎发达,形成的草坪色泽翠绿,质地厚且有弹性,在长江流域及以南区域的公园、广场草坪建植和公路两侧绿化中广泛采用[8,11]。日本结缕草和中华结缕草耐践踏能力强,在建植运动场草坪上有优势,在我国南北均有种植。然而,多年以来在我国广泛种植的结缕草品种还是以进口的为主[12]。优质品种是草坪业发展的核心,随着我国对草业发展的重视,近20 a来具有自主知识产权的品种不断推出,例如,2018年分别由华南农业大学和江苏省中国科学院植物研究所培育的广绿和苏植5号通过了全国草品种审定委员会审定。自有产权优质品种的推出将进一步促进我国结缕草产业的快速发展。
优质的草坪草不仅草坪质地优美,而且还需要有较强的适应逆境的能力,所以提高抗逆性一直是结缕草育种中的一项重要内容[13-15]。研究结缕草响应逆境胁迫的特征及抗逆的分子机理,对培育高抗逆品种有指导价值。日本结缕草基因组大小约334 Mb,预计有59 271个编码蛋白的基因[16]。在组学方面,Xuan 等[17]采用双向电泳技术研究了低温胁迫下日本结缕草和沟叶结缕草匍匐茎中的差异表达蛋白,Wei 等[18]和Long 等[19]先后报道了日本结缕草和沟叶结缕草在低温胁迫下的转录和生理调节,Wang 等[20]从转录组水平研究了大穗结缕草在盐迫下的调节,Wang 等[21]从生理和转录组水平对日本结缕草盐敏感和耐盐的2个株系进行了比较研究,这些结果为研究结缕草逆境响应基因的功能提供了丰富的信息。在基因功能研究方面,已经报道的逆境响应转录因子有DREB2[22]、DREB1[23-24]、锌指蛋白[25]、ICE[26-27]、NAC[28],功能基因有液泡膜质子-焦磷酸酶基因[29]、LEA基因[30-31]、GRP(Glycine-rich RNA-binding proteins)基因[32]、铜锌超氧化物歧化酶基因[33]及其分子伴侣基因[34]。由于结缕草的种质驯化历程短,诸多原始遗传性状得以保留,研究结缕草抗逆的分子机制还有利于挖掘新基因资源用于其他植物的育种。
COR(Cold regulated)基因是早期发现的一些在低温诱导下表达量显著上调的基因,例如拟南芥中的COR78/RD29A、COR47、COR15a和COR6.6,小麦中的脱水素(Dehydrin)和晚期胚胎发育蛋白。后来的研究发现,COR基因的表达通常也受干旱和ABA等因素诱导。COR蛋白的共性是富含亲水性氨基酸,能够维持生物大分子及膜系统的稳定性,具有增强细胞抗冻、抗脱水功能[35]。
本研究报道沟叶结缕草的一个低温胁迫响应基因ZmCOR410,分析了其表达产物的生物信息学特征,检测了其在低温胁迫下的表达情况,并在拟南芥植株和酵母细胞中评价了其抗逆功能,旨在为认识结缕草抗逆机制积累资料。
1 材料和方法
1.1 试验材料
供试材料为沟叶结缕草马尼拉,采自浙江大学华家池校区校园草坪。拟南芥为哥伦比亚生态型。植物表达载体pZP212和农杆菌Gv3101菌株为中央民族大学生命与环境科学学院生化与分子实验室保存材料,酵母表达载体pYES-Dest52和酵母G19菌株由中国科学院(江苏省)植物园刘建秀研究员惠赠。
1.2 ZmCOR410基因的克隆及生物学信息学分析
根据转录组数据设计引物P1(ATGGAGGACGAGAGGAACAAAGC)和P2(ATGGAGGACGAGAGGAACAAAGC)。以 4 ℃处理 72 h 的 cDNA 为模板,用P1和P2PCR扩增ZmCOR410基因的编码区(CDS)。PCR 扩增反应体系为(20 μL): 2×PrimeSTAR Max DNA Polymerase 10 μL,P1和P2各 0.25 μL(10 μmol/L), cDNA 模板 1 μL, ddH2O 8.5 μL。反应程序为 :(98 ℃ 10 s,58 ℃ 15 s,72 ℃ 10 s)× 32 循环。将PCR产物送到上海生工测序检测确定扩增产物的序列。利用GENSCAN(http://genes.mit.edu/GENSCAN.html)预测所编码的氨基酸序列;采用Protparam(http://web.expasy.org/protparam/)分析蛋白质的理化性质;通过TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)进行蛋白跨膜区分析;蛋白质二级结构通过SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测;系统发育树采用软件MEGA 7.0(NJ法,Bootstrap=1 000)构建。
1.3 ZmCOR410基因表达对低温胁迫的响应
沟叶结缕草种植于培养室内,培养基质为珍珠岩∶蛭石∶营养土=1∶1∶2,培养室温度为28 ℃/22 ℃(L/D),定期修剪。以修剪后6周龄植株为试材,转入培养箱中进行4 ℃低温处理,分别于2,6,12,24,72 h后取植株顶端第1~2片展开叶,同时取培养室内植株叶为对照,液氮中速冻后置于-80 ℃冰箱中保存备用。用植物总RNA提取试剂盒(TaKaRa)提取总RNA。选用经琼脂糖凝胶电泳和NanoDrop2000检验合格的RNA样品,以Oligo dT为引物,用反转录酶M-MLV反转录为cDNA,具体操作按说明书进行。以cDNA为模板,以actin1(NCBI accession No.:GU290545)为内标基因,采用qRT-PCR法检测各处理中ZmCOR410的表达量。内标基因引物为Pactin-F(GATATGCACTTCCCCATGCT)和Pactin-R(CGAGCTTCTCCTTGATGTCC),ZmCOR410基因的引物为5′—3′(GAAGCACGAGGTCAAGAAGG)和(TCTCGTCGATCACCTCCTCT)。反应总体系12 μL,其中cDNA 1 μL,SYBR Premix 6 μL,基因上、下游引物各0.24 μL。反应程序:95 ℃ 30 s;95 ℃ 5 s,56 ℃ 30 s,共40个循环。采用-ΔΔCt法计算基因的相对表达量。
1.4 拟南芥中评价ZmCOR410基因抗逆功能
将ZmCOR410CDS克隆到植物表达载体pZP212的XbaⅠ和SacⅠ之间,获得重组质粒pZP212-ZmCOR410。采用冻融法将重组质粒转化农杆菌Gv3101,通过浸花法转化拟南芥。拟南芥种子经次氯酸钠溶液(含2%有效氯)表面消毒后,播种到含有50 μg/mL硫酸卡那霉素的MS培养基上,筛选转基因植株,收集转基因植株T3种子用于抗逆评价试验。将野生型和转基因拟南芥植株种子播种在MS培养基平板上,10 d后将幼苗移栽至蛭石∶营养土=1∶1的基质中培养。冷冻处理是将移栽后28 d的苗子转到培养箱中,温度调到-6 ℃,2 h后洒冰水以诱导冰核形成,再继续冷冻处理12 h,然后将温度调到4 ℃化冻12 h。将植株转回到培养室中恢复生长,14 d后用叶片伤害指数来描述叶片的受损情况[36]。首先根据受损程度将叶片分为5级:基本无明显受损(0级)、表面有少量变黄(Ⅰ级)、将近1/2面积焦枯(Ⅱ级)、大部分焦枯(Ⅲ级)、全部焦枯死亡(Ⅳ级)。
①
高温处理是将移栽后两周龄的幼苗转入60 ℃培养箱中处理4 h,然后转入温室内恢复培养,14 d后观察记录植株存活率。干旱处理是幼苗移栽7 d后停止浇水,直到植株出现明显萎蔫症状,复水7 d后观察统计存活情况。以上胁迫处理设转基因植株和对照植株2个群体,样本量均为30株以上。采用χ2统计分析2个群体植株的存活率差异。
1.5 酵母细胞中评价ZmCOR410基因抗高温功能
采用Gateway重组技术将ZmCOR410的CDS克隆到酵母表达载体pYES-Dest52中,获得重组载体pYES-ZmCOR410。分别将重组载体和pYES-Dest52转入酵母G19菌株感受态细胞中,在亮氨酸缺陷型培养基(SD/Leu-)上筛选转化子。制备酵母细胞的无菌水悬浊液,参照张杰等[37]试验方法,在50 ℃下处理25,35,45 min,然后转到30 ℃复苏10 min。取100 μL菌悬液涂布接种到SD/Leu-培养基平板上,涂布未经高温处理的菌悬液为对照,30 ℃培养48 h后统计菌落数,计算存活率。每个处理均设置3个重复。
2 结果与分析
2.1 ZmCOR410基因的克隆
在前期的转录组测序结果中,有一个注释为COR410的拼接物,长1 717 bp,包含一个完整的阅读框。在预测的阅读框两端设计引物,以4 ℃低温处理72 h植株叶片的cDNA为模板,进行PCR扩增,结果获得了接近1 000 bp 的特异带(图1)。对扩增片段进行克隆和测序结果显示,该片段长927 bp,序列与预测的阅读框相符。
在NCBI网站上进行BlastX分析,结果表明,该基因氨基酸序列与小麦COR410基因的相似度高,因此命名为ZmCOR410。根据文献[16]下载沟叶结缕草基因组序列,用ZmCOR410的编码区(CDS)进行查找,获得其在基因组上的序列。将CDS、转录组拼接序列以及基因组序列进行比对可知,基因中含有一个长101 bp的内含子,翻译起始密码子往前1 700 bp序列中,正链含有2个ACCGAC序列和1个GCCGAC序列,反向互补链中有1个GCCGAC序列(图2),推测其转录受CBF/DREB1转录因子调节。

图1 ZmCOR410 编码区的PCR扩增Fig.1 The amplification of the coding sequence of ZmCOR410 by PCR
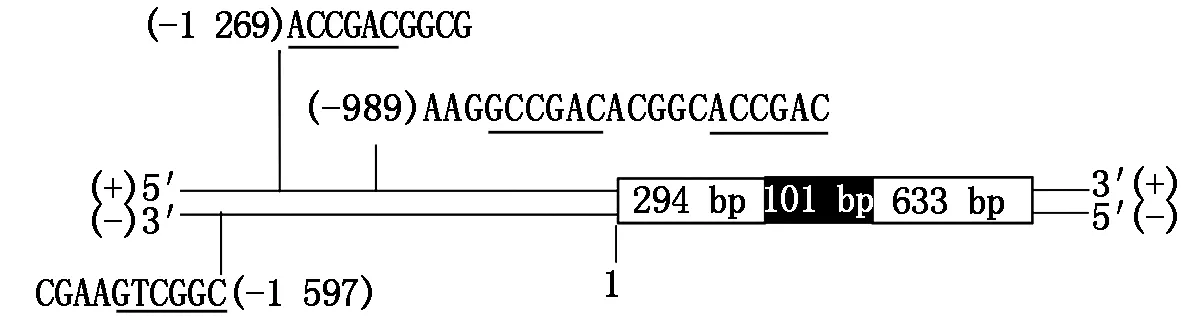
白色方块为外显子,黑色方块为内含子;(+)和(-)分别代表有义链和无义链;1为翻译起始位点。
2.2 ZmCOR410蛋白的结构特征
ZmCOR410基因编码308个氨基酸,主要的氨基酸组成为谷氨酸(Glu)14.6%、赖氨酸(Lys)13.0%、丙氨酸(Ala)9.1%、丝氨酸(Ser)7.8%和甘氨酸(Gly)6.2%。ZmCOR410蛋白质分子质量为33.8 ku,理论等电点为5.69,为偏酸性蛋白质。序列中有一个富含赖氨酸残基的基序,即K片段,还有一段可以被磷酸化的由多个丝氨酸残基组成的S片段,但其中缺少富含甘氨酸的重复序列,也不存在Y片段(DEYGNP)(图3)。这些特征都与LEA蛋白家族第Ⅱ亚组相符,该亚组也称D11蛋白,为脱水素。
亲疏水性分析表明,ZmCOR410氨基酸平均亲水性指数(GRAVY)为-1.277(图4-A),说明该蛋白为亲水性蛋白。利用SOPMA工具进行蛋白质的二级结构预测结果表明,ZmCOR410蛋白二级结构中α-螺旋、无规则卷曲、β-折叠和延伸链占比分别为34.09%,55.84%,5.19%,4.87%(图4-B),表明组成蛋白质分子的主要二级结构元件是α-螺旋和无规则卷曲,这样有助于蛋白质的稳定性[38]。利用TMHMM工具进行蛋白质跨膜区分析结果表明,所有氨基酸均位于细胞膜外,不含有跨膜区,说明ZmCOR410属于非跨膜类蛋白(图5)。ZmCOR410的上述一级和二级结构均符合酸性脱水素的特征[38]。

方框内为K-片段;下划线处为S-片段。 The box is K-segment;Underline is S-segment.
2.3 ZmCOR410基因的系统发育树分析
将ZmCOR410的氨基酸序列在NCBI网站上进行BlastP分析,选取与其氨基酸序列相似度最高的前30个同源基因(来自17个物种),使用Mega 7.0软件进行多序列比对并构建基因系统进化树,结果显示,这些基因明显地分成C4植物和C3植物2个大支。其中,C3分支主要是由麦类和水稻同源基因2个小分支构成;C4分支由玉米同源基因和其他C4植物同源基因2个小分支构成。结缕草ZmCOR410与抗高温干旱能力强的弯叶画眉草、无芒隐子草的同源基因亲缘关系最近(图6)。
图5 ZmCOR410蛋白氨基酸序列跨膜分析Fig.5 Transmembrane analysis of ZmCOR410 protein
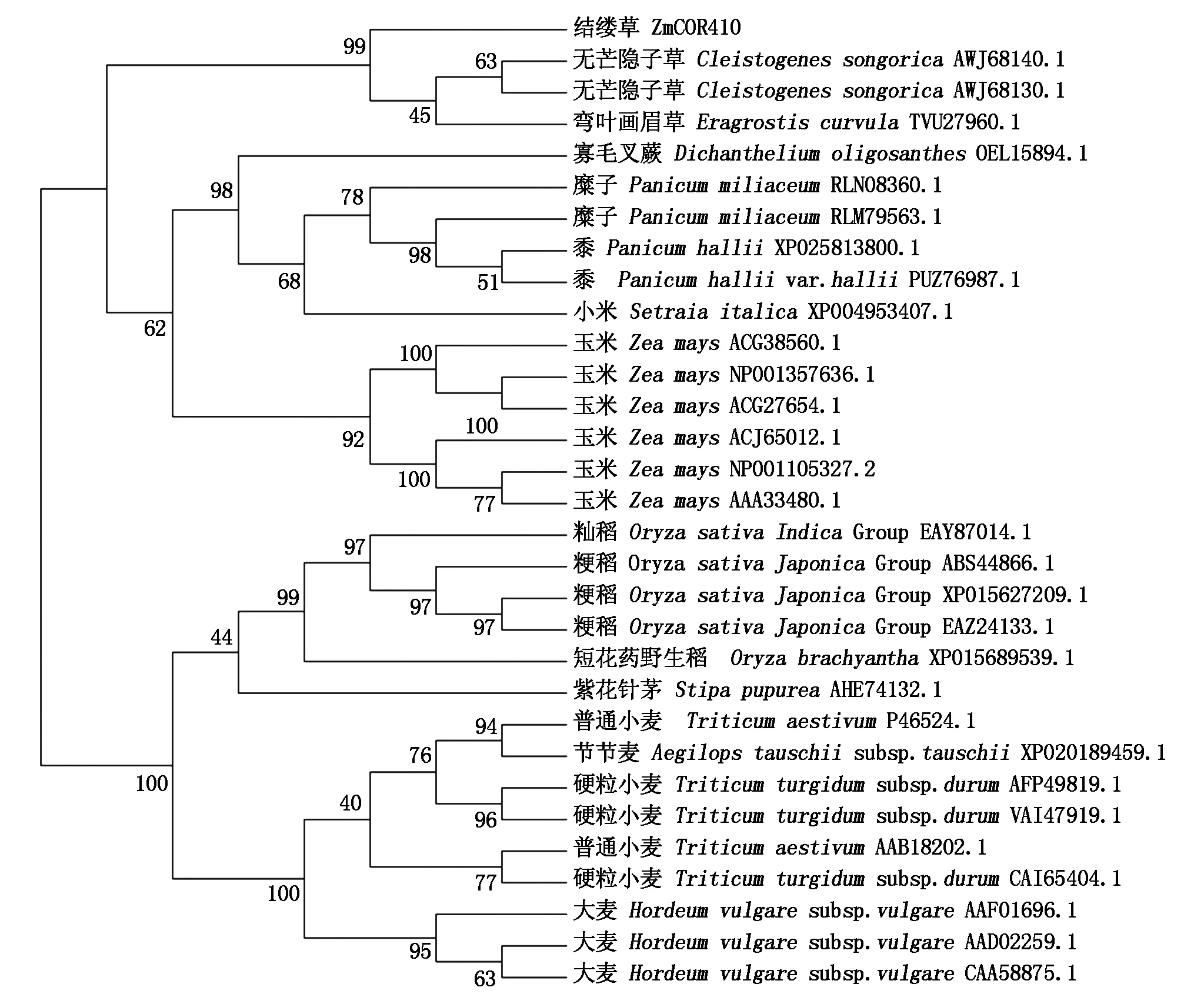
图6 ZmCOR410蛋白的系统进化树分析Fig.6 Phylogenetic tree analysis of ZmCOR410 protein
2.4 低温胁迫下ZmCOR410基因的表达调节
本研究检测了4 ℃低温胁迫下沟叶结缕草叶组织中ZmCOR410表达调节,结果表明,在常温条件下ZmCOR410基因表达量很低;在4 ℃低温胁迫下,2 h后其表达量明显上升,并随着胁迫的延长持续上升,增量最大的时间点在胁迫后24 h,直到胁迫后72 h其表达量持续上升。说明沟叶结缕草叶片中ZmCOR410基因的表达对低温胁迫敏感,受低温胁迫显著诱导(图7)。
2.5 转化ZmCOR410基因拟南芥植株的抗逆能力
为检测ZmCOR410 的抗逆功能,将其CDS克隆到植物表达载体pZP212的XbaⅠ和SacⅠ的位点之间,使其受PCaM35S启动子和T-nos终止子调控,得到植物表达载体(图8-A)。将该载体转化到农杆菌Gv3101菌株中,通过菌悬液醮花絮法转化拟南芥,经筛选获得了4个稳定遗传外源基因的株系。在正常生长条件下,转基因植株生长发育无明显特异之处。
以4个转基因株系的植株作为一个群体,以野生型植株为对照群体,进行胁迫处理,比较2个群体的抗逆能力。经-6 ℃冷冻低温处理后,转基因和野生型植株叶片均受到冻害;复苏培养14 d后,野生型群体中大叶片基本死亡,只有幼叶和芽尖有复苏生长;转基因群体中有的大叶片得以复苏,幼叶和芽尖复苏得更好,复苏相同时间后有更多的新叶长出(图8-B)。对伤害度进行统计,结果显示,转基因植株群体和对照群体叶片伤害指数分别为58.33,75.00,前者受到的伤害相对较轻。在干旱胁迫处理中,植株移栽复苏后即停止浇水,至萎蔫时大部分植株已经抽苔,复水后已经伸展的叶片和茎均不能恢复生长,仅部分腋芽能再生长出来,转基因群体和对照群体植株存活率分别为75.00%,58.33%(图8-C)。在高温胁迫处理中,植株经60 ℃高温胁迫处理4 h后,大部分叶片焦枯死亡,少数植株幼叶和芽尖复苏生长,转基因群体和对照群体存活率分别为26.04%,11.46%,二者差异达显著水平(P<0.05)(图8-D)。结果说明,ZmCOR410超表达后增强了拟南芥植株对冷冻低温、干旱和高温胁迫的耐受性,其中对高温耐受性的增强效果显著。

不同小写字母表示各处理时间存在显著差异(P<0.05)。图9同。 Different lowercase letters indicate that there are significant differences between processing times at 0.05 level.
2.6 转化ZmCOR410基因酵母对高温的耐受性
为了进一步验证ZmCOR410抗高温功能,pYES-ZmCOR410(图9-A)并将其转化到酵母G19菌株中。以转化pYES-Dest52空载体的酵母细胞为对照,酵母细胞悬浊液于50 ℃条件下分别处理25,35,45 min后,载有ZmCOR410基因的细胞存活率分别为63.22%,51.67%,45.32%,对照组细胞的相应存活率分别为56.08%,36.99%,33.65%(图9-B),说明50 ℃处理25 min以上使2种细胞的存活率均显著下降。统计结果显示,胁迫35 min以上时转化ZmCOR410的细胞存活率显著高于对照细胞,说明该基因增强了酵母细胞耐高温的能力。

不同小写字母表示差异显著(P<0.05)。 Different lowercase indicate significant difference(P<0.05).

A. 重组酵母表达载体;B. 酵母细胞存活率。 A.The recombinant yeast-expression vector;B.The survival rate of yeast cells.
3 结论与讨论
基于沟叶结缕草转录组数据,本研究克隆获得了沟叶结缕草的一个低温胁迫响应基因ZmCOR410,编码的蛋白ZmCOR410为一个酸性脱水素,在拟南芥和酵母细胞中表达后表现出一定的抗冻、抗干旱和抗高温胁迫功能。
COR410是禾本科植物普遍存在的一个低温响应基因,最早在小麦[39]中被检测到,与大麦中的HvDhn8[40]、水稻的OsDhn1[41]和拟南芥的COR47[42]为同源基因,均属于LEA 蛋白的第Ⅱ组(也称为脱水蛋白或LEA D11蛋白)。沟叶结缕草的ZmCOR410蛋白含有308个氨基酸,氨基酸平均亲水性指数为-1.277,无跨膜结构,二级结构以无规则卷曲为主,符合脱水蛋白的典型特征。在禾本科植物COR410同源基因系统进化树中,ZmCOR410与弯叶画眉草和无芒隐子草同源基因的亲缘关系最近。无芒隐子草是多年生超旱生C4植物,为干旱荒漠草原的建群种和优势种[43];弯叶画眉草多生长于沙质坡地、农田、路边荒地等,耐瘠、耐湿热、抗干旱,适应性很强[44]。推测这些物种中COR410基因的进化具有趋同性,为高温、干旱生境选择的结果。
ZmCOR410基因的表达调节对4 ℃非常敏感,mRNA水平在诱导6 h后即显著上升,并在72 h内呈现逐渐上升的趋势。芹菜(ApiumgraveolensL.)中COR47的转录在低温诱导72 h内也呈现逐渐上升的趋势[45],大麦HvDhn8的转录受4 ℃低温诱导并在第4天后基本稳定[40]。小麦COR410基因表达也受冷驯化诱导,第6天和第7天以后mRNA水平下降,但仍然保持在较高水平[38,46]。ZmCOR410转录对更长时间的低温胁迫或自然降温变化的响应有待探究。在拟南芥中COR47为典型的DREB1类转录因子的调节子,表达异源DREB1基因的转基因拟南芥中COR47的mRNA水平显著高于野生型[45];水稻的同源基因OsDhn1[41]、小麦同源基因TaCOR410[47]、大麦的同源基因HvDhn8[48]的表达也受DREB1诱导。沟叶结缕草中有多个DREB1基因,低温胁迫下表达量显著上调[19],ZmCOR410基因转录区上游有4个A/GCCGAC基序,推测低温下ZmCOR410基因的表达也受DREB1转录因子诱导。
对小麦的研究结果表明,COR410蛋白具抗冻功能,冷驯化过程中COR410蛋白表达量与品种抗冻能力强呈正相关[46]。将小麦COR410基因转入到草莓中,转基因植株叶片半致死温度比野生型的降低了5 ℃[49]。与此相似,沟叶结缕草ZmCOR410基因在拟南芥中超表达后可以减轻冷冻低温(-6 ℃)对叶片的伤害。本研究还发现,超表达ZmCOR410基因的拟南芥植株对高温和干旱胁迫的耐受性也有所增强,转化ZmCOR410基因的酵母细胞在高温胁迫下的存活率也明显提高。低温、高温和干旱胁迫均会引起细胞生理性缺水,COR410作为一种酸性脱水素,主要定位在细胞膜上,可能通过保护细胞膜来增强细胞对胁迫的耐受性[50]。小麦[38]和水稻[41]中的COR410同源基因在干旱胁迫下也大幅度上调表达,说明它们在抗干旱胁迫中的重要性。
综上所述,ZmCOR410是沟叶结缕草的一个酸性脱水素基因,其表达受低温大幅度诱导,表达产物可以增强细胞对冷冻低温、高温和干旱胁迫的耐受能力。胁迫环境下该基因的表达量与种质抗逆能力关系还有待进一步研究。
猜你喜欢
伴侣(2022年3期)2022-04-08
祝您健康(2020年5期)2020-05-14
名人传记·财富人物(2017年9期)2017-11-02
名人传记·财富人物(2017年9期)2017-11-02
Coco薇(2016年8期)2016-10-09
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
河北渔业(2015年1期)2015-01-19
河北渔业(2015年1期)2015-01-19
湖北农业科学(2014年15期)2014-10-28
