
玉米对灰斑病胁迫的生理生化响应
2022-09-14 04:22赵思琪曲比伍合余学杰柯永培苟齐贤石海春
华北农学报 2022年4期
赵思琪,曲比伍合,罗 婷,余学杰,,柯永培,,苟齐贤,石海春,
(1.四川农业大学 农学院,四川 成都 611130;2.四川正红生物技术有限责任公司,四川 成都 610213)
玉米(Zeamays)灰斑病于1925年在美国伊利诺伊州亚历山大县首次发现[1],于1991年在我国丹东首次报道[2],至今该病在全国普遍发生,对玉米生产造成了很大的威胁[3-5]。应用实践证明,筛选和培育抗病新品种以及研究它们抗灰斑病的生理生化机制等,可为品种选择与区域布局等提供更多科学合理的选择空间,也是解决这一困境的理想途径[6-8]。目前,针对玉米灰斑病的研究多集中在对其病原菌种类的鉴定[9-11]、种质抗病性鉴定[12-14]以及抗病基因定位[15-17]等方面,关于抗灰斑病的生理生化机制研究相对较少,且研究材料选择及相关研究内容比较单一。如吕静波[18]、刘可杰[19]、马莹莹等[20]以不同抗性的玉米自交系为材料,而郭红莲等[21]、王桂清等[22]则以杂交种为材料,其研究内容主要集中在灰斑病胁迫下不同防御酶的变化。然而,当玉米受到病原菌侵染后,其体内会发生一系列复杂的生理生化变化,如发病植株叶片叶绿素含量、净光合速率、气孔导度等[23-24]、防御酶类活性变化[25-26]以及诱发激素响应[27-28]。
本试验系统研究不同抗性玉米自交系及杂交种感染灰斑病后其光合特性、活性氧、防御酶以及激素的动态变化,旨在全面分析不同抗性玉米材料响应灰斑病的生理生化差异,明确其抗玉米灰斑病的生理生化机制,为资源鉴定与利用、品种选育与时空布局等提供参考。
1 材料和方法
1.1 试验材料
供试材料选用四川正红生物技术有限责任公司提供的玉米自交系K365(抗灰斑病)和K169(高感灰斑病)及杂交种正红431(高抗灰斑病)和正红532(高感灰斑病)。
1.2 试验设计
2021年春季将试验材料种植在四川正红生物技术有限责任公司四川荥经县试验基地,该试点为玉米灰斑病高发区,每个材料种植12行,每行种14株,小区面积40 m2,密度42 000株/hm2。从吐丝期开始,选取长势基本一致的植株单株挂牌,于吐丝后25,30,35,40,45 d采集叶片,用锡铂纸包住迅速投入液氮中,随后带回实验室于-80 ℃冰箱保存用于相关生理指标测定;于吐丝30 d(病斑出现前)和40 d(病斑出现后)时,对玉米穗位叶进行光合参数测定。
1.3 测定指标及方法
1.3.1 光合参数的测定 于09:00—11:00选取3株长势基本一致的植株,用LI-6800便携式光合作用测定仪测定其功能叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)等光合指标。
1.3.2 活性氧含量和抗氧化酶活性测定 用碘化钾法[29]测定过氧化氢(H2O2)含量,用愈创木酚显色法[30]测定过氧化物酶 (POD)活性,用NBT光化还原法[30]测定超氧化物歧化酶(SOD)活性,用紫外分光光度法[30]测定过氧化氢酶 (CAT)活性。
1.3.3 激素含量测定 用磷酸缓冲液提取内源激素,用酶联免疫吸附法测定内源激素水杨酸(SA)和茉莉酸 (JA)含量,试剂盒由重庆博诺恒生物科技有限公司提供。
1.4 数据处理
运用Microsoft Excel 2019软件进行统计分析和作图,运用SPSS 25.0软件进行单因素方差分析,采用Duncan's新复极差法检验显著性(P<0.05)。
2 结果与分析
2.1 灰斑病胁迫对玉米光合特征的影响
将出现灰斑病病斑前和病斑后不同材料的光合特征变化情况列于表1。由表1可知,玉米在出现病斑后比出现病斑前,Pn、Gs、Ci和Tr在不同的玉米材料中均表现为降低趋势,但感病材料的光合特征指标比抗病材料的下降趋势更加明显。针对感病材料,除杂交种正红532的Ci差异不显著外,不论是自交系K169还是杂交种正红532其光合特征指标下降程度均达显著水平(P<0.05),自交系K169的Pn、Gs和Tr分别下降59.32%,95.28%,80.18%,杂交种正红532的Pn、Gs和Tr分别下降85.39%,69.60%,56.77%。针对抗病材料,自交系K365和杂交种正红431的Ci均显著下降外(P<0.05),其余指标下降程度均未达显著水平,具体为自交系K365的Pn、Gs和Tr分别下降24.41%,25.00%,4.20%,杂交种正红431的Pn、Gs和Tr分别下降11.53%,21.43%,0.06%。综合表明,玉米灰斑病胁迫可严重影响其光合效率,且对感病材料的影响更大。

表1 灰斑病胁迫对玉米光合特征的影响Tab.1 Effects of gray spot stress on photosynthetic characteristics of maize
2.2 灰斑病胁迫对玉米过氧化氢含量的影响
灰斑病胁迫下不同玉米材料的H2O2含量变化见图1,可见其H2O2含量在不同抗性的玉米材料中变化规律不完全一致,主要表现为:针对感病材料,自交系K169的H2O2含量持续升高,杂交种正红532的H2O2含量后期高于前期,之后维持在较高水平,即感病材料整体表现为先上升然后稳定在一定高水平,具体为自交系K169的H2O2含量在吐丝45 d达到最大值,相较吐丝25 d增幅为83.63%,杂交种正红532的H2O2含量在吐丝35 d显著上升(P<0.05),相较吐丝25 d增幅为65.44%,之后维持在480.60~592.10 μmol/g水平;针对抗病材料,无论是自交系K365还是杂交种正红431其H2O2含量均分别在吐丝后35 d达最高,然后又显著下降到一定水平(P<0.05),即抗病材料表现为先显著上升后又显著下降趋势,具体为自交系K365的H2O2含量在吐丝35 d显著上升,相较吐丝25 d增幅为53.42%,杂交种正红431的H2O2含量在吐丝35 d显著上升,相较吐丝25 d增幅为69.12%。综合表明,灰斑病可以导致玉米叶片的H2O2含量升高,而胁迫后期感病材料的H2O2含量高于抗病材料,表明感病材料受到的氧化损伤程度更严重。

不同小写字母表示感病材料不同时间之间差异显著(P<0.05); 不同大写字母表示抗病材料不同时间之间差异显著(P<0.05)。图2—6同。 Different small letters in the figure indicate that the susceptible materials have significant difference at different time(P<0.05); different capital letters indicate that the resistant materials have significant difference at different time(P<0.05).The same as Fig.2—6.
2.3 灰斑病胁迫对玉米抗氧化酶活性的影响
2.3.1 灰斑病胁迫对玉米材料POD活性的影响 灰斑病胁迫下不同玉米材料的POD活性变化见图2,可见,除K365的POD活性呈先上升后下降变化趋势,其余3个材料POD活性均表现为先上升后维持稳定,但无论自交系还是杂交种,抗病材料POD活性达到峰值上升的幅度大于感病材料,具体表现为:自交系K365和K169的POD活性均在吐丝30 d达到最大值,但K365增幅为70.94%,而K169为49.74%;杂交种正红431和正红532的POD活性最大值分别在吐丝后40,35 d,其增幅分别为59.52%,54.63%,其POD活性较吐丝25 d均达显著水平。综合表明,灰斑病胁迫下,其POD活性均表现为显著上升趋势,且其抗病材料的POD活性增幅更高。
2.3.2 灰斑病胁迫对玉米SOD活性的影响 灰斑病胁迫下不同玉米材料的SOD活性变化见图3,可见,其SOD活性在不同抗性的玉米材料中变化规律不完全一致,针对感病材料,其表现为先显著上升再显著下降然后再上升趋势,具体表现为:自交系K169的SOD活性在吐丝35 d达到最大值,相较吐丝25 d增幅为14.50%,在吐丝40 d又显著下降至137.15 U/g(P<0.05),杂交种正红532的SOD活性在吐丝35 d达到最大值,相较吐丝25 d增幅为24.18%,在吐丝40 d显著下降至148.77 U/g(P<0.05);针对抗病材料则表现为显著上升到一定水平后不再显著变化,具体为自交系K365的SOD活性在吐丝45 d达到最大值,相较吐丝25 d增幅为21.12%,在吐丝35~45 d期间SOD活性维持在162.43~174.55 U/g,杂交种正红431的SOD活性在吐丝45 d达到最大值,相较吐丝25 d增幅为27.29%,在吐丝35~45 d SOD活性维持在164.25~174.53 U/g。综合表明,抗病材料比感病材料的SOD活性上升幅度更高,维持时间更久。
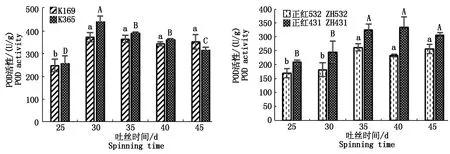
图2 灰斑病胁迫对不同抗性玉米材料POD活性的影响Fig.2 Effects of gray spot stress on POD activity of maize materials with different resistance

图3 灰斑病胁迫对不同抗性玉米材料SOD活性的影响Fig.3 Effects of gray spot stress on SOD activity of maize materials with different resistance
2.3.3 灰斑病胁迫对玉米CAT活性的影响 灰斑病胁迫下不同玉米材料的CAT活性变化见图4,可见,其CAT活性在不同的玉米材料中均大致表现为先上升后下降的趋势,但无论自交系还是杂交种,抗病材料CAT活性达到峰值上升的幅度要大于感病材料,具体表现为:自交系K365和K169的CAT活性均在吐丝35 d达到最大值,但相较吐丝25 d,K365增幅为25.33%,而K169为23.73%;杂交种正红431和正红532的CAT活性均在30 d达到最大值,但正红431增幅为107.64%,而正红532增幅为84.43%。综合表明,灰斑病胁迫下,其CAT活性均表现为先上升后下降的趋势,且其抗病材料的CAT活性上升幅度更高。

图4 灰斑病胁迫对不同抗性玉米材料CAT活性的影响Fig.4 Effects of gray spot stress on CAT activity of maize materials with different resistance
2.4 灰斑病胁迫对玉米内源激素含量的影响
2.4.1 灰斑病胁迫对玉米SA含量的影响 灰斑病胁迫下不同玉米材料的SA含量变化见图5,可见,其SA含量在不同抗性的玉米材料中变化趋势不尽相同,针对感病材料,自交系K169的SA含量只在吐丝45 d显著上升,杂交种正红532则表现为双峰趋势,具体为自交系K169在吐丝45 d达到最大值,相较吐丝25 d增幅为80.30%,杂交种正红532在吐丝30,40 d达到峰值,相较吐丝25 d增幅分别为77.99%,86.89%;针对抗病材料,其整体表现为先上升后下降趋势,具体为自交系K365在吐丝30 d达到最大值,相较吐丝25 d增幅为59.47%,杂交种正红431在吐丝35 d达到最大值,相较吐丝25 d增幅为85.34%。综合表明,不同玉米材料受到灰斑病侵染后体内SA含量呈不同程度的调节。

图5 灰斑病胁迫对不同抗性玉米材料SA含量的影响Fig.5 Effects of gray spot stress on SA content in maize with different resistance
2.4.2 灰斑病胁迫对玉米JA含量的影响 灰斑病胁迫下不同玉米材料的JA含量变化见图6,可见,其JA含量在不同抗性的玉米材料中变化规律不完全一致,针对感病材料,其整体表现为先上升后下降趋势,具体为自交系K169在吐丝35 d达到最大值,比吐丝25 d上升68.80%,杂交种正红532在吐丝40 d达到最大值,比吐丝25 d上升88.27%;针对抗病材料则表现为双峰趋势即先上升后下降再上升再下降,具体为自交系K365分别在吐丝30,40 d达到峰值,比吐丝25 d分别上升了79.10%,107.75%,杂交种正红431也分别在吐丝30,40 d达到峰值,比吐丝25 d分别上升了92.83%,89.11%。综合表明,抗病材料的JA含量比感病材料上升幅度大且时间早。

图6 灰斑病胁迫对不同抗性玉米材料JA含量的影响Fig.6 Effects of gray spot stress on JA content in maize with different resistance
3 结论与讨论
3.1 玉米抗灰斑病的光合特性基础
光合作用是植物生长的生理基础,有研究表明,植物的光合能力受到诸多因素的影响,病菌侵染会导致叶片的光合性能受损,Pn、Gs和Tr下降[31-33]。而对玉米感染病害后光合特性的研究大多集中在对杂交种的探究上[34-35]。本研究同时以不同抗性的玉米自交系和杂交种为研究材料,结果表明,在灰斑病胁迫下,它们的Pn、Gs、Tr和Ci均呈现不同程度的下降,但无论自交系还是杂交种,抗病材料在灰斑病胁迫下光合特性下降不显著,而感病材料的光合特性显著下降,推测其原因可能是因为感病材料由于病菌侵染导致气孔关闭,进入气孔CO2量减少,最终导致净光合速率降低。
另外,当叶片Gs、Tr和Ci同时下降时,净光合速率下降的因素为气孔限制;反之如果叶片净光合速率的降低伴随着胞间CO2浓度的升高,则光合作用的限制因素是非气孔限制[36],本研究中,灰斑病胁迫下叶片Ci下降,说明K365、K169、正红431和正红532的光合受到抑制是由气孔因素引起的。
3.2 玉米抗灰斑病的抗氧化系统基础
植物在正常生长状态下,体内活性氧(ROS)含量达动态平衡,当植物受到病原菌侵染后,体内ROS的产生和清除动态平衡会被打破[37],其中H2O2是ROS的一种,非生物胁迫会导致细胞内H2O2的积累,其含量可间接反映植物耐受程度[38]。抗病材料可以及时控制体内的H2O2平衡,感病材料则会发生紊乱[39]。本研究中,抗病材料能将H2O2维持在稳定水平,与前人结果相符,而感病材料则表现出不一致的变化规律,其中自交系K169的H2O2持续升高,杂交种正红532的H2O2在吐丝40 d有显著下降的趋势,分析其原因可能是由于杂交种因存在杂种优势而抵抗灰斑病的能力要强于自交系。
当ROS含量升高到一定程度时会对植物造成伤害,为了减轻ROS的伤害植物会启动抗氧化系统来抵御氧化损伤,主要包括SOD、POD和CAT等抗氧化酶类和ASA、GSH等非酶类抗氧化剂[40-42],其中酶类的抗氧化剂被认为是清除ROS最直接、最有效的途径[43]。本研究结果表明,抗病材料的POD、SOD和CAT活性上升的幅度及上升持续时间要高于感病材料,与前人研究[18-19,21]基本一致。
但值得注意的是本研究发现,无论自交系还是杂交种,抗感材料的抗氧化酶活性差异表现均出现在病斑之后,如K365叶片的SOD活性在出现病斑后仍维持较高水平,而K169叶片的SOD活性显著下降;正红431叶片的CAT活性在出现病斑后维持在初始水平,而正红532叶片的CAT活性下降到初始水平的0.64倍。这可能与抗病材料抑制病斑扩展有关,感病材料由于防御酶活性较低,可能无法及时清除H2O2,导致体内H2O2积累,进一步对植物形成氧化胁迫,最终导致病斑扩散,因此“病斑出现”可能是抗感材料抗氧化系统差异的关键时期。
3.3 玉米抗灰斑病的内源激素基础
SA和JA是重要的信号分子,在植物抗病信号传导过程中充当重要的角色[44-45]。当受到病菌侵染会导致体内SA和JA发生明显变化[46],且不同抗性植株会对病害胁迫做出不同程度的响应[47-48]。潘凤英[49]研究表明,2个不同抗性水稻幼苗感染南方水稻黑条矮缩病后SA迅速积累,并在症状出现后达到最大值,高抗材料的SA含量高于高感材料。鲁秀梅等[50]研究表明,不同抗性甜瓜接种病菌后体内JA含量有显著变化,高浓度的JA可提高对蔓枯病的抗病能力。本研究表明,抗病材料在刚出现病斑时SA含量为较高水平,JA含量为较低水平;而感病材料在刚出现病斑时SA含量为较低水平,JA含量为较高水平,可综合推测出,在刚出现病斑时SA和JA存在拮抗作用,且也进一步证实了“病斑出现”可能是抗感材料抗氧化系统差异的关键时期。
综上所述,玉米灰斑病胁迫下,其感病材料光合速率显著下降,以及活性氧增加导致叶片氧化水平失衡,抗病材料可能通过合理积累SA、JA等信号分子来调控抗氧化系统,使其在感病后期体内仍维持较高水平的防御酶活性来更有效清除多余的H2O2,同时调节叶片的内源激素平衡使其适应灰斑病带来的影响,从而增强玉米的抗病性。
猜你喜欢
西南农业学报(2022年7期)2022-09-30
中国果树(2022年9期)2022-09-06
中国种业(2022年8期)2022-08-19
贵州农业科学(2022年4期)2022-05-06
江苏农业科学(2022年6期)2022-04-15
中国瓜菜(2019年8期)2019-09-19
科学导报(2018年47期)2018-05-14
农家科技下旬刊(2017年6期)2017-07-05
农民致富之友(2016年5期)2016-10-21
绿色科技(2016年7期)2016-05-14
