
海洋微藻小粒级化趋势的环境驱动机制浅析
2022-09-21 09:12刘苏萱
水产科学 2022年5期
刘 印,宋 伦,刘苏萱
( 1.大连海洋大学,辽宁 大连 116023;2.辽宁省海洋水产科学研究院,辽宁省海洋生物资源与生态学重点实验室,辽宁 大连 116023 )
海洋微藻作为海洋生态系统中主要的初级生产者,通过光合作用将二氧化碳转化为有机碳,增加海洋碳汇,对维持地球生态系统具有非常重要的作用[1-3];海洋微藻亦是浮游动物、鱼类、滤食性贝类等海洋动物的主要营养来源,而微藻细胞粒径的大小直接或间接影响这些动物的营养储备和健康生长,尤其对饵料质量选择型的滤食性贝类影响更大[4-5]。海洋微藻按照细胞的等效球径大小分为小型微藻(>20 μm)、微型微藻(2~20 μm)和微微型微藻(<2 μm)。相关研究表明,环境的变化会影响海洋微藻的群落组成和粒级结构[6-8],海洋微藻种群或群落正趋于小粒级化[9-10]。笔者统计了我国四大海区微藻优势种的演替规律(表1)[11-18],统计结果显示,海洋微藻群落经过20多年的演替,优势种的粒径有趋向小粒级化的趋势。
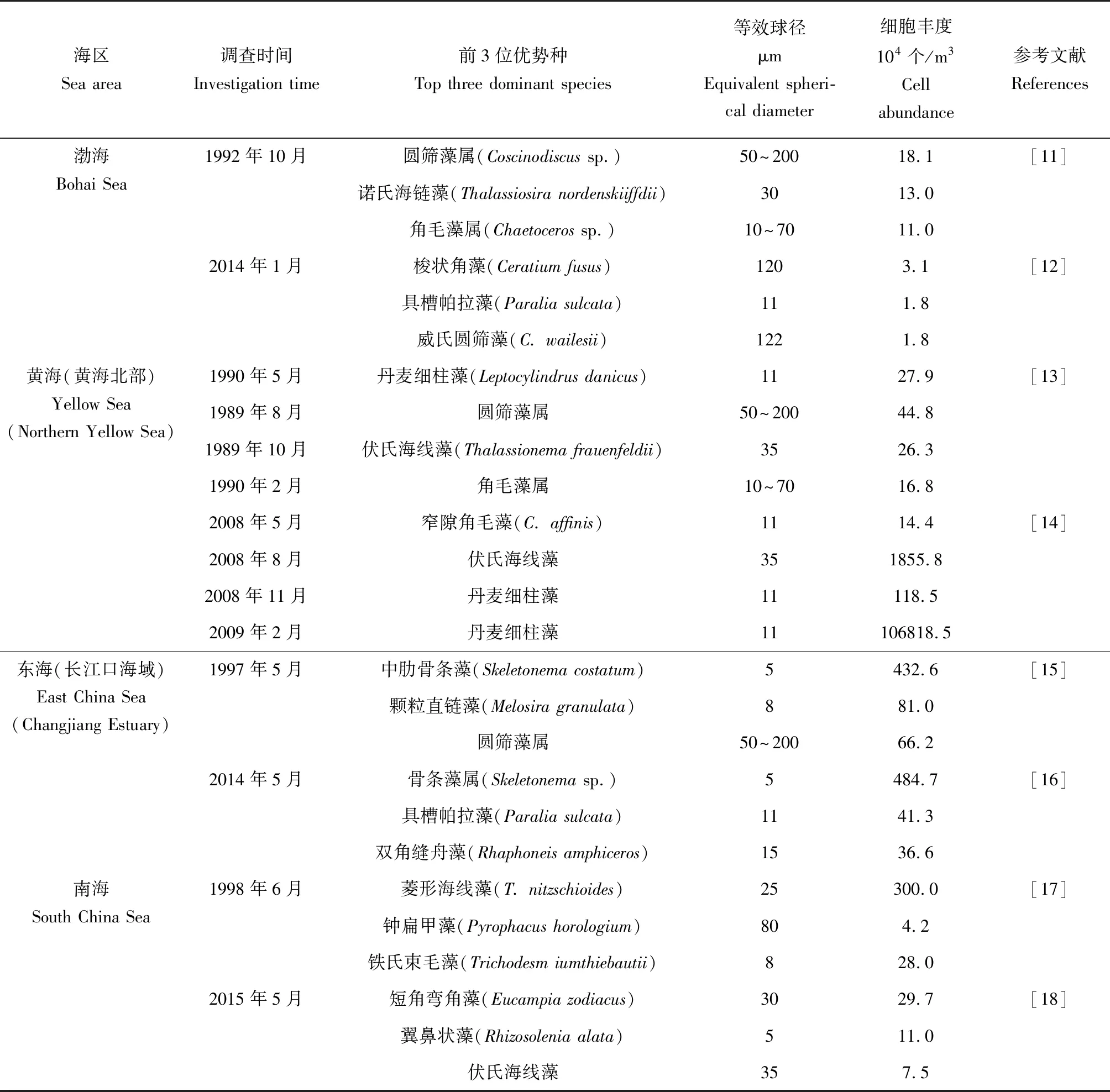
表1 中国近岸海域微藻优势种演替规律Tab.1 Succession of dominant species of microalgae in coastal waters in China
驱动海洋微藻小粒级化的因素主要为3个方面:(1)全球气候变暖和海洋酸化;(2)营养盐组成及含量失衡,如氮、磷、硅含量下降;(3)其他因素诱导,如微量元素和摄食压力等。笔者围绕这3个方面展开分析,以期为海洋生态保护和贝类增养殖业的健康发展提供参考。
1 气候变暖驱动海洋微藻小粒级化
温度是影响海洋生态系统的一个重要环境因子,可以直接或间接地影响海洋中的大部分生物、物理或化学过程。温度可以直接影响海洋微藻的新陈代谢和生长繁殖,是海洋生物时空分布的重要影响因素[19];温度可以影响海水的密度和盐度等环境因子,共同控制其垂直稳定性,进而影响海洋温跃层的形成与发展[20];温度可以影响海洋生态系统中的多种化学反应过程,如海洋有机物的降解、吸附等[21],进而影响海洋系统中碳、氮、氧等多种元素的地球化学物质循环;温度还可以通过影响其他环境因子,如海水的水体黏度、溶解气体等环境因子影响海洋生物。
根据适宜生长的温度阈值,将海洋微藻划分为暖水种、冷水种、广温种、狭温种[22]。当海洋微藻生长的温度突破阈值时,不同微藻的反馈调节能力差异较大,适应新环境温度的海洋微藻成为优势种群得以继续生存[23-24]。全球变暖已成为不争的事实,1850—1950年,全球气温平均上升了0.74 ℃,并且正以每10 a增加0.2 ℃的速度上升,预计到2100年,地球表面平均气温将增加1.1~6.4 ℃[25],随之海水温度升高,为微型和微微型微藻的生长提供了适宜环境,使其繁殖速度加快、生长周期变短,进而快速占领系统中的生态位,同时小型微藻的生长受到抑制,最终驱动了微藻的小粒级化趋势。Daufresne等[9]在加利福尼亚海域调查发现,海洋微型及微微型微藻的粒径大小与水体温度显著负相关。夏滨等[26]调查发现,受黄海冷水团的影响,温跃层上层即冷水层中的大粒级海洋微藻占有优势。Atkinson等[27]建立了海水温度与微藻细胞体积的量化模型,发现海洋微藻的粒径随温度的升高而减小,每上升1 ℃细胞体积减少约2.5%。Montagnes等[28]也发现了硅藻粒径随温度升高而减小的规律,指出海水每上升1 ℃硅藻细胞体积平均减少约4%。虽然目前对具体的影响模式还存在争论,但不可否认的是海水温度的上升会驱动海洋微藻的小粒级化[29-31]。
2 海洋酸化导致海洋微藻小粒级化
海水通常情况下为弱碱性,由于人类过度使用化石燃料、滥砍滥伐等,造成大量的二氧化碳等温室气体排放,二氧化碳被海水吸收后会破坏海水碳酸盐系统的动态平衡,使海洋酸化。预计到2100年,海水表层pH将降至7.8,比1800年的酸度高150%[32-33]。海洋酸化将破坏海洋微藻细胞的渗透压,直接影响微藻吸收各种营养盐的能力[34-35]。
我国海域海水酸化问题比较普遍,除了某些pH异常区域,我国其他海域海水pH最低为7.8[36],同时有学者发现,南海海域海水酸化的速度开始加快[37]。富营养化使近岸海域水体的微生物呼吸作用增强,加上海洋酸化的效应使近岸海域的酸化速度高于大洋[38]。有研究发现,在海水酸化和气候变暖的双重作用下,白令海中海洋微藻的群落结构组成发生了改变,海洋微藻趋向于以微型微藻为主,因此降低了以硅藻为基础的食物链生产能力[39]。低pH导致微藻吸收营养盐的能力降低,而小粒级微藻由于较快的环境反馈机制,能够快速占领水域中的生态位,进而抑制大型微藻的生长,最终微藻小粒级化趋势加剧。
粒径较小的海洋微藻(如微型、微微型微藻)比粒径较大的海洋微藻(如小型微藻)不易受到海水酸化的影响[40-41]。如增加海水中二氧化碳浓度可显著提高微微型藻类——抑食金球藻(Aureococcusanophagefferens)生长及比生长速率,同时该藻会抑制滤食性贝类对其的摄食利用,进而提高了抑食金球藻的生态位,导致海区微藻小粒级化趋势。同时研究发现:当氮、磷、硅营养盐充足时,海水酸化对桑沟湾不同粒径微藻生长的影响不显著,但会减少硅藻的种类数,同时显著增加微微型藻类的数量,导致微藻粒级结构发生改变;在磷限制的情况下,酸化处理组(pH 7.9、7.6)可显著促进微型藻类的生长,在氮、磷限制的情况下,pH 7.9酸化处理组可显著促进微微型微藻繁殖,导致微藻粒级结构趋于小粒级化[42]。
3 营养盐组成及含量失衡致使海洋微藻小粒级化
营养盐对海洋微藻的生长限制作用主要体现在两个方面:一是在营养盐浓度很低时难以维持海洋微藻的生长;二是营养盐结构发生改变,致使某种或某几种营养盐限制海洋微藻的生长。海洋微藻生长需要多种营养要素共同维持,主要的营养盐为氮、磷、硅3种[43-44],当营养盐浓度或结构发生改变时,将会影响微藻种群生长,导致群落结构发生改变。
3.1 营养盐含量失衡
营养盐浓度变化对微藻粒径影响较大,尤其氮、磷的含量直接影响微藻的体积和数量[45]。邓春梅等[46]研究发现,营养盐浓度和微藻叶绿素a含量密切相关,并且这种相关性随着微藻粒径的变化而改变。当微藻粒径>8 μm时,营养盐浓度的升高会促使微藻叶绿素a含量随之升高;当微藻粒径<8 μm时,营养盐浓度的升高会导致微藻叶绿素a含量下降。可以推断,粒径较大的微藻在富营养海域中占据优势,而粒径较小的微藻在贫营养海域中占有优势。黄邦钦等[47]调查发现:2000年秋季在高营养盐的长江口海区微型和小型微藻占据优势(82%),而在黄海中部等寡营养海区微微型微藻占比增大(41%);同样的现象也出现在2001年春季,在营养盐丰富的长江口海域,微型和小型微藻占有绝对优势(99%),而微微型微藻则对黄海中部和东海陆架外缘等寡营养海区的贡献率较高(分别为50%和62%)。
当然,受微藻粒级分析技术限制,微藻粒级分析结果可能存在差异[48-49],但总体上反映的趋势应是一致的。宋伦等[4]采用高通量测序—分子鉴定分级技术研究发现:2018年春季大连长海县大长山岛邻近海域微微型微藻主要受总氮影响较大,夏季受总氮、总磷影响显著,秋季受溶解硅影响显著;小型微藻在夏季主要受溶解无机磷影响较大;微型微藻受上述环境因子影响不显著。
张霞等[50]在研究中指出,珠江口海域微微型藻类的生物量与营养盐浓度呈负相关,表明微微型微藻对寡营养海域有着良好的生理适应机理和生存策略。一般来说,由于微微型藻类细胞较小,细胞表面积与体积比较大,生理代谢较快,能量转换的效率高,比粒径较大的藻类具有更高的光合作用速率与摄取营养盐的能力,因此可以迅速适应贫营养水体,并且在其中快速生长。当水体中的无机盐被迅速消耗时,小型微藻会受到磷的限制,而微型和微微型微藻能够迅速繁殖,抢占有利生态位,从而驱动海洋微藻群落的小粒级化。
蔡昱明等[51]对1998—1999年南极普里兹湾的微藻与叶绿素a含量的研究结果显示:从普里兹湾到毗邻陆架区再到大陆坡的硝酸盐与磷酸盐的比例依次上升;硝酸盐的比例由普里兹湾内的(17.71±7.72)%增长到毗邻陆架区的(28.05±3.11)%,最后增长到大陆坡的(28.54±2.46)%;磷酸盐的比例由普里兹湾内的(1.26±0.14)%增长到毗邻陆架区的(1.94±1.03)%,最后增长到大陆坡的(2.02±0.35)%。随之增加的微型微藻的含量占比由25.9%增至27.5%和32.5%,而微微型微藻的含量占比上涨更加迅速,从2.8%升至23.3%和22.5%。可以看出,硝酸盐和磷酸盐在毗邻陆架区和大陆坡的比例很接近,同时明显高于普里兹湾,微型微藻含量增加幅度小,微微型微藻含量从普里兹湾到毗邻陆架区增加幅度大,但在毗邻陆架区和大陆坡略有下降,说明微藻粒级与营养盐浓度呈负相关关系。
宋伦等[8]研究发现,辽东湾富营养海域微藻主要以小型为主,寡营养海域主要由微型和微微型的链状或杆状的骨条藻、帕拉藻、角毛藻、菱形藻等组成。叶绿素a分级测定表明,寡营养海域<20 μm微藻的叶绿素a含量均高于富营养海域。也有研究表明,寡营养海域微微型微藻是初级生产力的主要贡献者[52]。
3.2 营养盐结构失衡
近些年,为保护水域生态环境,国家采取多种措施,大力消减氮、磷入河入海总量,而入海河流截流会导致入海硅含量明显降低,致使硅氮比、硅磷比不断降低[53],近岸海域营养盐结构变化较大,进而导致海洋微藻群落和粒级结构发生演替。
研究发现,营养盐结构的变化会直接影响海洋微藻的种群结构,甚至引发赤潮致灾种演替现象[54-55]。从长期变化看,黑海西部大陆架海水中氮、磷含量的增加和硅含量的下降,会导致河口海域微微型微藻的比例由0.1%增至41.7%[56]。我国长江口海域也发生了相同的变化,近40年来,随着陆源营养盐输入,长江口海域氮含量持续增加,而硅含量持续下降,导致硅氮比和硅磷比明显下降,造成小型硅藻种类逐渐减少,而微型甲藻的种类明显增加[57]。胡明辉等[58]研究表明,长江口外海域海洋微藻的生长繁殖也受到磷的限制;Harrison等[59]认为,该海域氮磷比过高,而离河口500 km以外的海域则受氮的限制;Jiao等[60]研究发现,20世纪80年代胶州湾的营养盐浓度和结构就开始发生改变,氮含量呈上升趋势,硅酸盐含量呈下降趋势,海洋小型微藻丰度明显减少,微型和微微型微藻占据了小型微藻空出的生态位;孙军[61]也证明了微型藻类米氏凯伦藻(Kareniamikimotoi)在氮磷比为80∶1的条件下生长最快。近些年,长江口海域营养盐浓度及其结构的变化导致海洋微藻优势种类逐步由以硅酸盐为主要营养的硅藻类向甲藻类转变[62]。
对比英国凯尔特海域与北美西海岸海域的研究可以发现:高营养盐海域的微藻优势种类与寡营养盐海域的优势种类有较大差异,在富营养海域,海洋微藻优势种群是体积较大的硅藻,在寡营养海域,则是体积更小的甲藻;北美西海岸硅藻丰度较少,而微型甲藻的丰度较高,成为该海域微藻主要类群[41,63]。
Gayoso[64]对Bahía Blanca河口海域进行了长达几十年的研究,发现在1978—1993年间,河口海域的海洋微藻优势种为硅藻门的小型微藻海链藻,而2006年之后,由于入海营养盐结构变化,该河口海域的优势种由海链藻逐步向微型微藻小环藻(Cyclotella)演替;Guinder等[65]也发现,相对大粒级的海链藻已经被小粒级的角毛藻和小环藻取代;辛明[66]对长江口海域的研究结果显示,自2003年开始,小粒级(微型)的海洋微藻(主要为甲藻)生物量显著增加,而以硅酸盐为主要营养的硅藻(小型)生物量逐渐下降,主要由入海营养盐浓度下降和结构失衡所致。
董婧等[67]对黄海北部近岸海域微藻群落组成的研究结果显示,1998年6月黄海北部近岸甲藻生物量占比为50.45%,取代了硅藻,成为优势种,在营养盐相对贫乏(尤其是磷缺乏)的黄海北部水域,甲藻的生长、繁殖比硅藻更迅速。
4 其他因素促使海洋微藻小粒级化
4.1 其他环境因子
除以上几种因素外,海洋微藻粒级结构还受其他环境因子的影响。Hare[68]研究表明,在秘鲁海域连续增加的三价铁离子浓度会加快海洋微藻群落从粒级较大的硅藻向粒级较小的蓝藻和微微藻演替;Shi等[69]发现,在海水酸化条件下,水中游离的三价铁离子浓度会升高,促使海洋微藻粒级结构发生改变。
宋伦等[70]研究发现:黄海北部春季,刺激小粒级微藻生物量显著增加的环境因素除活性硅酸盐外,还有悬浮物,而水深、重金属汞、盐度、砷会抑制其生长;夏季与小粒级微藻生物量显著正相关的环境因素除氮、磷、硅营养盐外,还有石油烃、重金属铜,显著负相关的环境因素有水深、pH、盐度、重金属铅;秋季与小粒级微藻生物量显著正相关的环境因素有重金属汞、砷,显著负相关的环境因素有水深、pH、盐度。总体来看,黄海北部营养盐、悬浮物、石油烃及部分重金属可促进小粒级微藻的生长,直接影响小粒级微藻的时空变化。
宋伦等[71]在研究辽东湾潜在褐潮生物时空分布与环境关联时发现,辽东湾各站位不同季节潜在褐潮生物(微微型藻类)丰度除与无机氮、氮/磷比、水温显著相关外,还与水深显著相关,秋季小粒级微微型藻类主要分布在较深水域。
宋伦等[72]还发现:在辽宁河口及港口无机氮含量高的浑浊带海域,微藻生物量主要集中在微型和微微型微藻区域;而在低营养盐的清澈海域,微藻生物量主要集中在小型微藻区域。这表明微藻群落对高浓度悬浮物的响应可能通过微型和微微型微藻进行反馈调节,致使圆筛藻等小型微藻生物量组成降低。
4.2 摄食压力
海洋微藻是海洋生态系统的重要生产者,可作为滤食性贝类和浮游动物等的重要食物来源,由于两者的摄食策略不同,导致海洋微藻粒级结构发生改变。滤食性贝类摄食主要靠鳃丝和其上着生的纤毛一起组合运动来摄取饵料[73],而海洋微藻的粒径大小直接影响着贝类的摄食。规格较小的贝类由于滤食器官尚未完全发育,摄食率随贝类规格的增加而增大,大规格贝类对微藻的摄食具有选择性,不同规格贝类最适宜的饵料粒径范围有所不同[74-75]。浮游动物也是海洋微藻的重要摄食者,海洋中浮游动物的摄食会消耗10%~75%的微藻[76]。研究表明,中型浮游动物主要摄食小型微藻,微型浮游动物更偏好摄食微型微藻[77];马莎[42]研究发现,中规格海湾扇贝(Argopectenirradians)对褐潮藻(微微型微藻)的摄食率随藻细胞密度的增大先升后降,虾夷扇贝(Patinopectenyessoensis)对微型微藻的摄食量显著高于微微型微藻,长牡蛎(Crassostreagigas)对小型和微型微藻的摄食量高于微微型微藻;Chen等[78]研究发现,海洋微藻的粒径越大,其被捕食的概率就越高,二者呈现出显著的正相关关系。
5 结 论
气候变暖驱动海洋微藻小粒级化。目前许多研究证实了全球变暖和生物的小粒级化趋势显著相关,认为生物小粒级化是生物对气候变暖生态响应的第3个普适性定律。许多研究也表明,随着海水温度的升高,海洋小粒级微藻的丰度与优势度不断增加,海洋微藻粒级逐渐向小粒级化演进。
海洋酸化导致海洋微藻小粒级化。海洋酸化可抑制大粒级微藻生长,导致硅藻的种类下降,同时会促进微微型微藻的生长,增加其数量,导致微藻粒级结构趋向小粒级化。研究也发现,在海水酸化和气候变暖的双重作用下,海洋微藻群落趋向小粒级化。
营养盐下降及结构失衡致使海洋微藻小粒级化。许多研究表明,寡营养海域小粒级微微型微藻是初级生产力的主要贡献者,而粒径较大的微藻在富营养海域中占据优势。近些年,陆源输入营养盐减少较多,尤其河口海域硅、磷限制严重,致使硅氮比、硅磷比结构失衡,不断下降,近岸海域营养盐结构变化较大,进而导致海洋微藻粒级结构趋向小粒级化。
其他环境因子和摄食压力促使海洋微藻小粒级化。研究发现,三价铁离子会促使海洋微藻群落从粒级较大的硅藻向粒级较小的蓝藻等微型微藻演替。除营养盐外,黄海北部悬浮物、石油烃及部分重金属可促进小粒级微藻的生长,直接影响小粒级微藻的时空变化。海水酸化和贝类摄食双重作用显著促进了微微型微藻的生长,导致微藻粒径结构发生改变。海洋微藻的小粒级化趋势是种群自我保护的一种策略,也是适应进化的一种方式。
6 展 望
个体大小是生物的基本特征,它反映了生命有机体生理生态和进化的历史。目前大量研究结果证实了海洋微藻小粒级化是普遍存在的,科学家们试图寻找普适性的规律来解答这一现象。从进化适应的角度考虑,小粒级微藻因其具有较高的表面积/体积,在能量和物质的获取利用方面常占优势。因此,在贫营养环境下,微微型微藻(蓝藻、甲藻)比个体较大的微藻(硅藻)更易获得营养资源并形成竞争优势。一般来说,由于微型和微微型微藻细胞较小,细胞表面积/体积较大,有着比小型微藻更薄的边界层,其热量更易耗散,因此在温度较高的环境下往往更易于生长和繁殖,另外相对于小型微藻,微型和微微型微藻因数量多、生长周期短、繁殖速度快等特点,在寡营养环境中或摄食压力大的生境中更易占据优势。
微藻种群正趋于小粒级化,这一现象不仅在近岸海域存在,也在远海被发现,并且该现象可能会由食物链的低营养级向着高营养级传递,最终影响人类食物安全。然而水域中的微藻为什么会发生小粒级化?其中哪些因素是导致微藻趋于小粒级化的原因?笔者通过分析相关文献,确定驱动海洋微藻小粒级化的主要环境因素为:全球气候变暖、海洋酸化、营养盐含量下降结构失衡、其他环境因子协同作用和摄食压力。
人类活动与全球气候变化的综合影响破坏了海洋生态系统内部的个体之间、群落之间、不同营养级之间的平衡关系。如低营养级微藻粒级结构的改变,使海洋微藻类群乃至整个生态系统演变的环境响应机制变得更加复杂,单个环境因子无法解释整个海洋中微藻粒级结构的变化,笔者所得结论还需进一步验证和完善。今后研究应关注以下几个问题:微藻小粒级化到底发生在种群水平中还是群落水平中?对环境变化的响应体现在表型适应中还是基因水平中?微藻小粒级化现象是否会传递给高营养级生物及如何影响?这些科学问题有待科学家们共同思考与研究。
猜你喜欢
中国动物保健(2021年7期)2021-11-23
考试与评价·高二版(2021年2期)2021-09-10
知识就是力量(2021年6期)2021-07-09
天天爱科学(2021年6期)2021-06-08
当代化工(2020年6期)2020-08-24
大自然探索(2019年9期)2019-12-13
科学之谜(2019年3期)2019-03-28
农业与技术(2016年24期)2017-04-20
江苏农业科学(2016年11期)2017-03-21
湖北农业科学(2017年1期)2017-03-09
