
贵州草海鲫鱼HIF-1α和HIF-2α基因克隆与表达分析
2022-09-23 12:11葛传龙欧阳力剑金志梅
江苏农业科学 2022年17期
李 青, 葛传龙, 欧阳力剑, 金志梅, 杨 钦
(1.贵州省典型高原湿地生态保护与修复重点实验室,贵州毕节 551700; 2.贵州工程应用技术学院生态工程学院,贵州毕节 551700)
云贵高原位于我国西南部,是我国四大高原之一,海拔在400~3 500 m间。贵州草海国家级自然保护区位于云贵高原中部顶端,是一个典型的高原湿地生态系统。鲫鱼不仅是我国重要的淡水养殖经济鱼类,而且由于其多生活于水域的中下层,具有极强的低氧耐受性,是公认的低氧耐受力极强的鱼类之一。作为草海的优势鱼种,鲫鱼在长期高原低氧环境中,进化出特有的身体机能以维持机体O平衡,弥补机体的缺氧反应。
低氧诱导因子HIF是由和亚基组成的一种异质二聚体的转录因子,参与了动物缺氧应激的适应和生存过程,在维持机体氧稳态中发挥至关重要的作用。和亚基均包含基本的螺旋-环-螺旋结构基序和PAS结构域。在常氧条件下,脯氨酰羟化酶PHD通过氧依赖性降解结构域ODD中2个保守的脯氨酸残基的羟基化作用,使靶蛋白HIF亚基羟基化,随之羟基化的HIF亚基被VHL蛋白识别并泛素化,介导蛋白降解。HIF激活细胞中一系列缺氧诱导基因的转录,以应对缺氧环境。在脊椎动物中HIF至少有3个亚型,分别为HIF1、HIF2和HIF3。鱼类中的研究已经证明,HIF1主要在无氧呼吸中间体的氧气摄取和运输中起关键作用,而HIF2在血管和红细胞的生成发挥重要作用。目前,关于HIF亚型的研究已在多种鱼类中开展,而在耐低氧的鲫鱼中的研究还鲜见报道。基于此,本研究基于前期对草海鲫鱼转录组数据分析,利用生物信息学技术和PCR,克隆获得了鲫鱼中HIF 2个亚型和的全长 cDNA序列,并进行了结构和功能分析。此外,利用实时荧光定量PCR分析了鲫鱼中和mRNA在不同组织中的表达情况,以期为进一步深入探究鲫鱼耐低氧分子机制奠定基础。
1 材料与方法
1.1 样品采集
于2018年在贵州威宁县草海采集鲫鱼[体长(14.98±4.25) cm;体质量(68.71±5.25) g],取其肠、肝脏、鳃、心脏、肾脏、肌肉、脾脏、精巢、卵巢和全血组织样品,放入RNA Later(Tiangen)中,采样点:26°51′24.15″N,104°12′31.57″E,海拔:2 168.09 m。另采集3尾鲫鱼带回实验室,取其肝脏组织,液氮冷冻后置于-80 ℃低温冰箱保存,用于后续RNA提取。
1.2 鲫鱼Ca-HIF1α和Ca-HIF2α全长的克隆与生物信息学分析
提取鲫鱼肝脏组织总RNA,将RNA反转录为cDNA(TaKaRa PrimeScript Ⅱ 1st Stramd cDMA Synthesis Kit)。从鲫鱼转录组数据库筛选和基因片段,使用DNAstar SeqMan软件对筛选的目的片段进行拼接。根据拼接获得的和基因序列设计特异性引物,由表1可知,对拼接获得序列的正确性进行验证,克隆获得和基因的全长cDNA序列。利用DNAMAN软件分析和基因全长cDNA序列中开放阅读框及其编码的氨基酸序列,SMART(http://smart.embl-heidelberg.de/)分析和基因编码氨基酸序列的功能域,ExPASy(http://web.expasy.org/protparam)分析和基因编码蛋白质的分子质量和等电点(pI)。
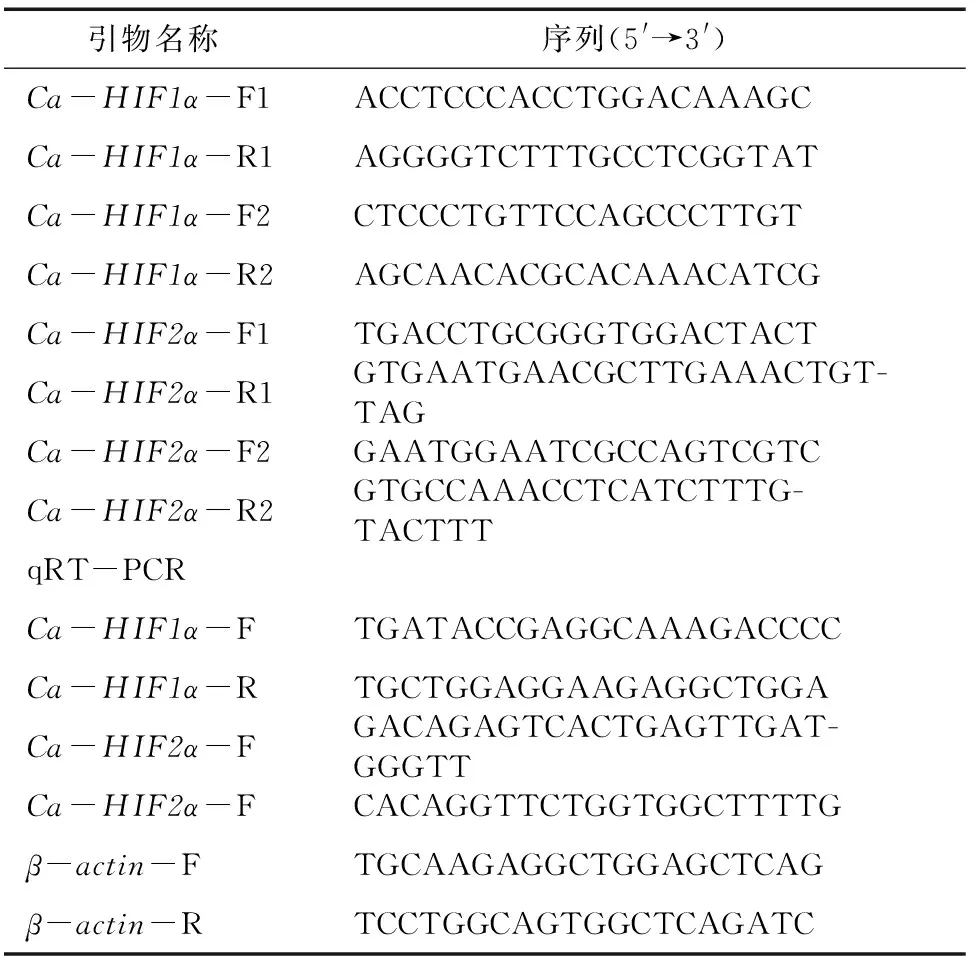
表1 Ca-HIF1α和Ca-HIF2α基因序列扩增和qRT-PCR引物序列
1.3 HIF1α和HIF2α序列比对和系统进化树的构建
利用Clustal X软件比对鲫鱼与其他物种的和基因亚型翻译的氨基酸序列的同源性,并用MEGA 4.0软件进行同源与聚类分析,采用邻接法NJ构建系统进化树,500次自举(Bootstrap)重复检验进化树的置信度。表2为各物种序列登录号,来自GenBank。

表2 序列比对和构建进化树物种信息
1.4 鲫鱼Ca-HIF1α和Ca-HIF2α mRNA表达模式分析
分别提取肠、肝脏、鳃、心脏、肾脏、肌肉、脾脏、精巢、卵巢和全血组织的RNA,将RNA反转录为cDNA(TaKaRa SYBR Premix Ex),根据和基因全长cDNA序列设计特异性引物,鲫鱼-基因作为内参,由表1可知引物序列,每个样本设计3个平行重复,使用BioRad CFX96TM Real-Time PCR检测系统对和mRNA在不同组织中的表达模式进行分析。
1.5 数据分析
使用Bio-Rad CFX Manager软件分析熔解曲线,通过2-ΔΔ分析和mRNA的相对表达水平。采用GraphPad Prism 5.0统计软件中的One-way ANOVA对和mRNA在鲫鱼不同组织相对表达量进行分析,<0.05为差异显著。
2 结果与分析
2.1 鲫鱼HIF-1α和HIF-2α cDNA序列分析
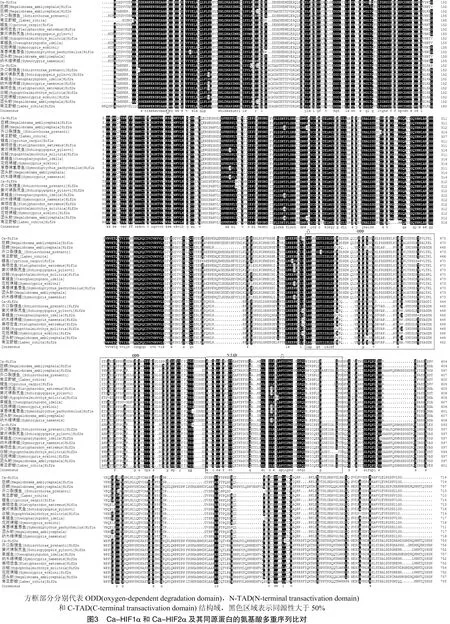
克隆获得鲫鱼中和cDNA全长序列(GenBank NO.:MW916102,MW916103),将其分别命名为和。由图1可知,全长共有3 867 bp,其中,5′UTR为295 bp,3′UTR为1 247 bp,开放阅读框2 322 bp,编码774个氨基酸。由图2可知,cDNA全长共4 647 bp,其中5′UTR为175 bp,3′UTR为 2 023 bp,开放阅读框2 460 bp,编码820个氨基酸。生物信息学分析预测和基因编码蛋白质的分子质量分别为85.844、91.228 ku,等电点为5.12和6.20。SMART在线分析见图3。由图3可知,二者具有HLH、PAS和PAC结构域,此外,多序列比对结果显示二者还具有ODD、N-TAD 和C-TAD功能域。


2.2 HIF1α和HIF2α亚型的同源性分析以及系统进化树的构建
多序列比对结果见图4。由图4可知,鲫鱼 Ca-HIF1α 第27个氨基酸是丝氨酸(S),而Ca-HIF2α相对应的第25个氨基酸是半胱氨酸(C)。Ca-HIF1α和Ca-HIF2α氨基酸序列不仅具有ODD、N-TAD和C-TAD 3个结构域,且ODD结构域具有2个保守的脯氨酸(P)位点,其中,1个是LxxLAP位点,该位点的序列并没有发生突变。此外,C-TAD结构域均具有保守的天冬酰胺(N)位点,表明Ca-HIF1α 和Ca-HIF2α结构域的保守性。进化树显示,Ca-HIF1α和Ca-HIF2α分别与其他物种中相应的亚型聚为一支。但Ca-HIF1α是先与花斑裸鲤、纳木错裸鲤、扁咽齿鱼、黄河裸裂尻鱼、齐口裂腹鱼、鲤鱼的HIF1α聚为一支后,再与南亚野鲮的HIF1α聚为一支,最后与白鲢、草鲤鱼和团头鲂的HIF1α聚在一起。与之不同,白鲢、草鲤鱼和团头鲂HIF2α先聚为一支,南亚野鲮、齐口裂腹鱼、花斑裸鲤、纳木错裸鲤、黄河裸裂尻鱼和扁咽齿鱼聚为一支,然后2支汇聚,最后再与Ca-HIF2α聚在一起,提示Ca-HIF2α比Ca-HIF1α进化得要快。

2.3 Ca-HIF1α和Ca-HIF2α mRNA在鲫鱼不同组织中的表达
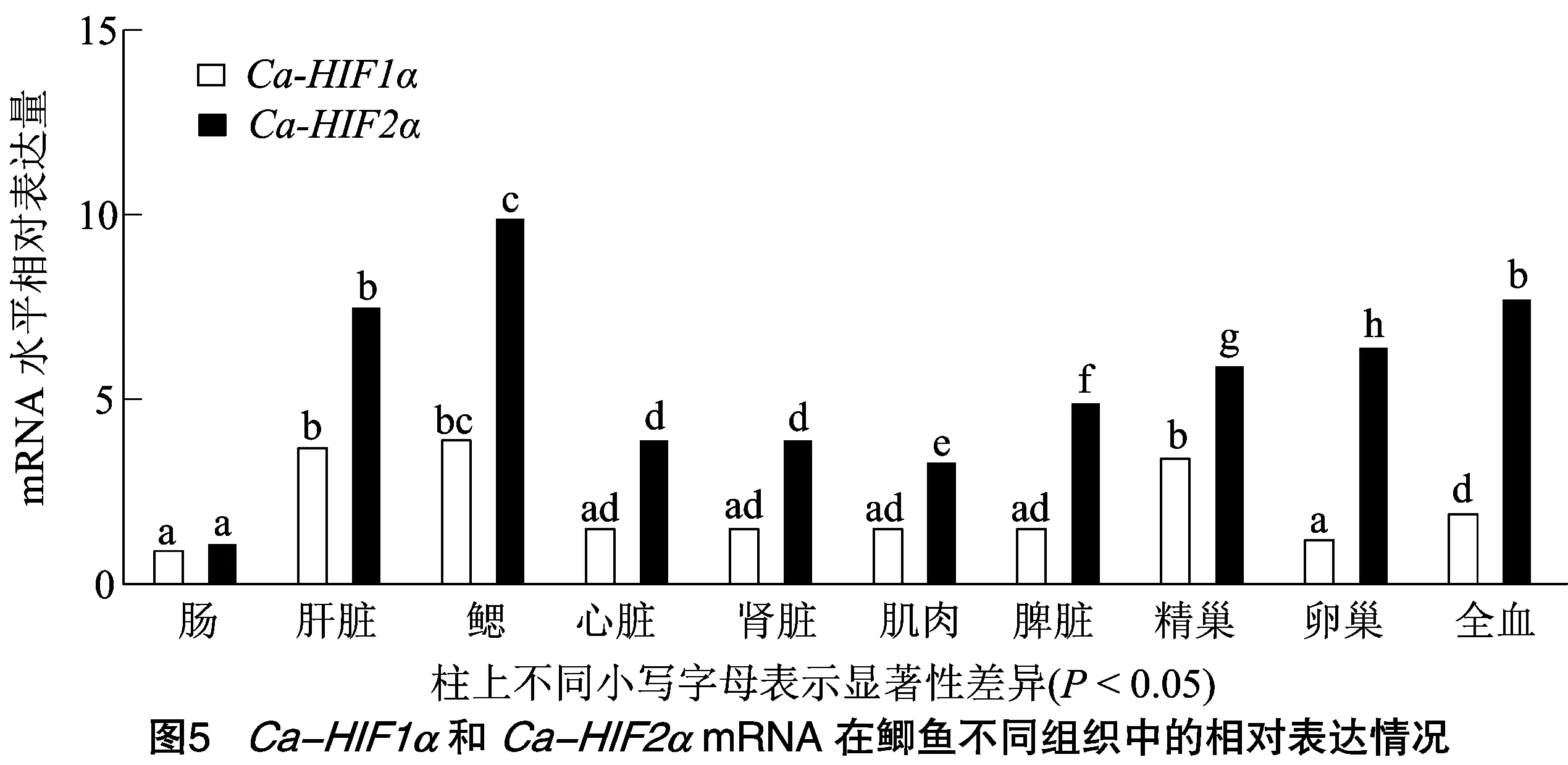
由图5可知,实时荧光定量PCR(qRT-PCR)结果显示,和mRNA在所有被检测的组织中均有表达,其中,mRNA在所有组织中的相对表达量均高于mRNA的表达量。此外,mRNA在肝脏、鳃和精巢组织中的相对表达量最高,全血、心脏、肾脏、肌肉、脾脏和卵巢组织中的相对表达量次之,在肠组织中的相对表达量最低。与之相似,mRNA在肝脏、鳃和全血组织中的相对表达量最高,卵巢、精巢、脾脏、心脏、肾脏和肌肉组织中的相对表达量次之,在肠组织中的相对表达量最低。

3 讨论
3.1 鲫鱼中Ca-HIF1α和Ca-HIF2α基因序列与系统进化分析
本研究克隆获得鲫鱼中基因2个亚型的全长cDNA序列,对序列比对构建进化树显示二者分别与其他物种中的HIF1α和HIF2α聚为一支。因此,将其分别命名为和。与其他物种中的同源蛋白一样,Ca-HIF1α和Ca-HIF2α均具有HLH、PAS-A、PAS-B、PAC、N-TAD和C-TAD 6个结构功能域,表明组成HIF1α和HIF2α的主要结构域是高度保守的。与巨鲷()和HIF1α相关研究结果一致,Ca-HIF1α第27个氨基酸是丝氨酸(S),Ca-HIF2α对应位点的第25个氨基酸是半胱氨酸(C),HIFα中HLH结构域中还原性半胱氨酸的存在主要与介导动物的氧敏感性有关,此不同之处导致Ca-HIF2α在DNA结合过程中可能比Ca-HIF1α对氧化还原更敏感。Ca-HIF1α与Ca-HIF2α结构上的这些区别可能导致二者具有各自独特的功能,且Ca-HIF2α在系统进化中比Ca-HIF1α要快。然而,Ca-HIF1α和Ca-HIF2α在ODD结构域均具有2个保守的脯氨酸(P),且与人类中的HIFα同源蛋白相似,保有了LxxLAP位点,LxxLAP位点的突变是动物适应高原低氧环境的重要标志,这可能与鲫鱼是耐低氧动物有关。此外,Ca-HIF1α和Ca-HIF2α在C-TAD结构域均具有天冬酰胺,2个保守的脯氨酸主要参与了蛋白质周转,天冬酰胺主要参与诱导靶基因的转录,这些残基的保守性,表明其在促进鱼类和哺乳动物的转录活性方面具有功能保守性。
3.2 鲫鱼不同组织中Ca-HIF1α和Ca-HIF2α mRNA水平表达差异分析
实时荧光定量PCR结果显示,在常氧条件下,和mRNA在所检测组织(肠、肝脏、鳃、心脏、肾脏、肌肉、脾脏、精巢、卵巢和全血)中均有表达,与巨鲷()、斑马鱼、齐口裂腹鱼()和软刺裸鲤()中研究结果相一致,和mRNA在鳃组织中的表达量均最高。此外,与巨鲷()相似,mRNA在鲫鱼肝脏和精巢组织中的表达量均很高。而mRNA在卵巢组织中表达量较高,mRNA在卵巢组织中的表达量较低。与同时耐低氧的草鱼中结果不同,和mRNA在鲫鱼的肾脏组织中表达量均不高。和mRNA在鲫鱼肠组织中的低表达,可能是由于肠道组织水平的日常变化,二者参与了肠道稳态的维持。以上结果说明-在维持机体氧稳态方面发挥至关重要作用,但和这2个亚型mRNA水平的表达差异性表明二者在调控氧稳态方面具有分工。此外,二者在精巢、卵巢和全血组织中的表达,推测其可能参与介导了血细胞和生殖细胞的发育。
猜你喜欢
东坡赤壁诗词(2022年4期)2022-10-30
华人时刊(2022年21期)2022-02-15
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
植物保护(2019年4期)2019-09-04
安徽农业科学(2018年22期)2018-05-14
江苏农业科学(2016年8期)2017-02-15
数位时尚(幼儿教育)(2016年9期)2016-04-10
海峡姐妹(2016年7期)2016-02-27
淮阴工学院学报(2014年5期)2014-09-10
