
基于ISSR标记和数量性状的火龙果种质材料遗传多样性分析
2022-09-29 04:42陈明贤
亚热带植物科学 2022年2期
陈明贤
(泉州市农业科学研究所,福建 晋江 362212)
广义的火龙果是仙人掌科植物,分属量天尺属(Hylocereus)、昙花属(Epiphyllum)和蛇鞭柱属(Selenicereus),原产南美洲,是我国华南热区近年新兴果树,产区主要分布于海南、广西、广东及贵州等省(区),具有速生快长、适应性强、产量高等特点。火龙果品种多、资源丰富,目前对其种质材料(含品种、品系、株系、品种资源,下同)的分类主要依据果皮和果肉颜色分为红皮白肉、红皮粉红肉、红皮红肉、红皮紫肉、黄皮白肉、橙皮红肉等多种类型,现国内栽培主要为红皮白肉及红皮红肉类型,少量为黄皮白肉类型[1—3]。目前,国内对火龙果的研究集中在栽培[4]、果实营养成分分析[5—6]、采后储藏保鲜[7]、叶面肥对果实食用及储藏品质的影响[8]、产品开发[9]以及种质资源调查、收集与鉴定等领域[10—11]。有关火龙果种质资源分子标记研究的报道主要涉及 ISSR[12]、SSR[13]、RAPD[14]、AFLP[15]等,在表型性状研究上,已有研究表明火龙果花、果实表型性状、农艺性状及品质性状均具有丰富的遗传多样性[16—17]。
ISSR分子标记技术已十分成熟并广泛应用于果树种质资源鉴定、遗传多样性与亲缘关系分析等方面研究[18]。在遗传多样性分析上,虽然分子标记技术拥有稳定性好、重复性高的特点及优势,但表型性状便于田间观察,且各性状均是遗传基因与环境互作的直接结果,也是育种工作的最终目标,因而通过表型性状的差异化选择,仍是育种工作的主要手段。将二者合理地结合起来,对实际育种工作有更大的促进作用。
本研究采用分子标记技术和数量性状相结合的方法,对收集保存在泉州国家农业科技园区果蔬园内的25份火龙果种质资源进行遗传多样性分析,以期更全面地反映收集地种质材料的遗传背景和性状表现,为下一步选育种工作提供理论支持与技术指导。
1 材料与方法
1.1 材料
1.1.1 供试材料
供试火龙果种质材料为泉州市农业科学研究所近年收集保存,种植于泉州国家农业科技园区果蔬园火龙果引种圃,共25份(表1)。所有种质均进行统一田间管理,于2019年12月剪取健壮嫩茎,以茎尖作为ISSR分子标记试验材料,液氮处理后保存于-80 ℃冰箱,备用。

表1 供试火龙果种质材料信息Table 1 Germplasm information of tested pitaya
1.1.2 主要仪器与试剂
紫外可见分光光度计(TU-1810),台式高速冷冻离心机(BECKMAN Avanti30、Eppendoff 5804R),梯度PCR仪(Tgradient-Biometra),DYCP-32A电泳仪(北京六一),凝胶成像系统(Syngene Gene Genius),ATAGO-PAL糖度测量仪等。
2×TaqPCR Master Mix(TIANGEN 公司);DL2000 DNA Ladder (北京索莱宝科技有限公司);Trans 2K Plus DNA Maker (北京全式金生物技术有限公司)。ISSR引物为前期筛选出的对火龙果有高特异性的7条引物:UBC807、UBC808、UBC810、UBC811、UBC853、UBC873、UBC880,由北京三博远志生物技术有限责任公司合成。
1.2 方法
1.2.1 基因组DNA提取及质量检验
采用多糖多酚植物基因组 DNA提取试剂盒(TIANGEN公司)提取火龙果基因组DNA,用1.2%琼脂糖凝胶电泳检验质量。
1.2.2 ISSR-PCR扩增体系及程序
ISSR-PCR 体系:2×TaqPCR Master Mix 10 μL、10 μmol·L-1引物 1 μL、10 ng·μL-1DNA 模板 1 μL,用无菌水补齐至20 μL。PCR反应程序:94 ℃预变性 5 min;94 ℃变性 30 s,(Tm-5) ℃ (根据引物而定)退火45 s,72 ℃延伸90 s,共37个循环,循环结束后4 ℃保存。PCR扩增产物用2.0%琼脂糖凝胶电泳检测,以5 V·cm-1电压电泳60 min。电泳后用凝胶成像观察结果,拍照保存。
1.2.3 数量性状观测
主要观测火龙果植株及果实的 20个数量性状(表2),数据测量参照广西壮族自治区地方标准DB45/T 1761-2018《火龙果种质资源描述规范》的方法,茎蔓数据测量选取1~2年生无病害健康茎蔓于2020年4~6月进行,果实性状于8~9月测量,每性状测量10个数据。
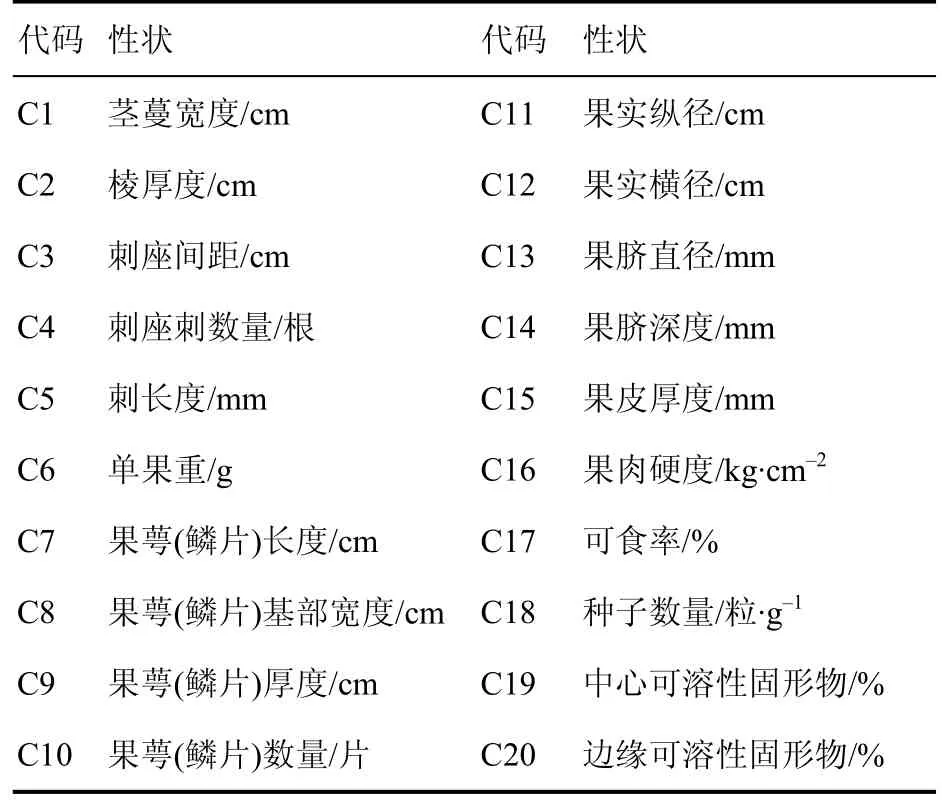
表2 测试火龙果的数量性状Table 2 Quantitative characteristics of pitaya for testing
1.2.4 数据分析
根据凝胶中条带迁移的位置和有无进行统计,计算多态性条带比例,用Ntsyspc2.1进行UPGMA聚类分析。对供试品种数量性状的最大值、最小值、均值、标准偏差(SD)、种内变异系数和种间变异系数进行统计分析,用SPSS 22.0软件对不同品种进行单因素方差分析。对原始数据标准化处理后,根据平方Euclidean距离,按Ward法进行系统聚类分析,构建树状图。
2 结果与分析
2.1 ISSR引物多态性分析
用7条ISSR引物分别对25份火龙果种质材料进行多态性检测,扩增出多个位点且条带清晰,不同种质各有特异性条带(图1、表3)。7条引物共检测到 97个位点,引物的扩增位点数为 12~16,平均每条引物扩增13.8个位点,其中扩增位点最多的是UBC810和UBC880,最少的是UBC873。扩增的多态性位点数为93个,多态性比例为95.88%,其中引物UBC808、UBC810、UBC811、UBC873扩增的多态性比例达100%,UBC880最低,仅87.50%。

图1 引物UBC873的扩增结果Fig. 1 The amplified result by the primer UBC873

表3 ISSR引物及扩增产物多态性Table 3 ISSR primers and the polymorphism of the amplified products
2.2 基于ISSR分子标记的火龙果种质UPGMA聚类分析
UPGMA聚类分析表明,25份火龙果种质材料的相似系数在0.41~0.86之间,在遗传相似系数为0.54处,可将25份火龙果种质材料分为6大类(图2)。第Ⅰ大类有17份种质,包括黑白双色水晶、桂红龙1号、台红3号、南宁蜜龙、晋红、越南红肉、紫龙、美龙2号、红玫瑰5号、红冠、湛江10号、柬埔寨红肉、台红2号、长红、海红、台红1号、黑龙,均为红皮红肉品系;比较遗传相似系数可发现,柬埔寨红肉与台红2号、台红3号与南宁蜜龙、红冠与湛江10号等3组相似系数最为接近,再比对扩增的图谱,7条引物扩增出的条带具有高相似度,说明其基因组差异小,且聚类分析结果可靠。第Ⅱ大类有3份种质,包括晋白、海白、蜜红龙。第Ⅲ大类有2份种质,包括赤龙和红绣球。红仙蜜、黄龙变种、白巨龙各自为一类。

图2 基于UPGMA法的25个火龙果种质聚类分析Fig. 2 Dendrogram of 25 pitaya germplasms by UPGMA method
2.3 火龙果主要数量性状的变异情况
对 25份火龙果种质的主要数量性状进行统计分析,结果显示,火龙果各数量性状的种内变异系数在 7.25%~27.51%之间,超过 20%的有棱厚度(C2)、单果重(C6)、果萼(鳞片)厚度(C9)、果萼(鳞片)数量(C10)、果脐深度(C14)和种子数量(C18)等6个;低于10%的有可食率(C17)、可溶性固形物(C19、C20)等3个。种间变异幅度在12.35%~51.66%之间,变异系数较大,变异丰富。变异系数最大的为棱厚度51.66%,可溶性固形物的变异系数最小,中部和边缘变异系数相近(表4)。种内变异幅度大,则种间的变异幅度相应的也较大。

表4 火龙果种质资源数量性状变异情况Table 4 Variations of quantitative characteristics among pitaya
2.4 基于数量性状的火龙果种质聚类分析
用Ward法聚类分析,在欧式距离为5处,可将25份种质聚为6个组群(图3)。第Ⅰ组群包括美龙2号、红玫瑰5号、南宁蜜龙、红冠、湛江10号、台红2号、台红3号、海红、晋红、柬埔寨红肉和紫龙等11个品种,均为红皮红肉品系,整体品质表现较好;第Ⅱ组群包括晋白、海白、蜜红龙、桂红龙1号、白巨龙和越南红肉6个品种,红皮白肉品系均在该组群;第Ⅲ组群包括长红、台红1号、黑白双色水晶等3个品种;第Ⅳ组群包括红仙蜜、赤龙和红绣球3个品种,均为红皮白肉品系,突出表现是刺座数量密集,刺长且多,品质表现一般;黄龙变种和黑龙各为一组,黄龙变种鳞片多且密,可食率低下,黑龙单果重量小、茎厚,两品种品质较差。

图3 基于数量性状的25份火龙果种质聚类图Fig. 3 Cluster diagram of 25 pitaya germplasms based on quantitative traits
2.5 不同聚类方法分析比较
对图2和图3的聚类结果进行比对,两种聚类方法得出的结果有部分相似,两者均能把性状表现比较一致的品种聚为同一群组,如晋白与海白、赤龙与红绣球、红冠与湛江10号等,分子标记聚类的第Ⅰ组群,包括数量性状聚类的Ⅰ组群,均为红皮红肉品系。性状表现差异较大的品种也能单独区分,如黄龙变种和黑龙。但两种聚类方法也存在较大差异。单从果肉颜色看,两种聚类方法均无法明确地区分红肉类型和白肉类型,通过数量性状聚类,虽然能将3个白肉类型的聚为一类,晋白与海白的欧式距离更近,与白巨龙较远,但同一组群中也包含了部分红肉类型品种;通过分子标记聚类可以看出,存在与数量性状聚类分析大体一致的结果。另外,在分子水平上遗传距离较近且聚为同一组群的品种,如柬埔寨红肉与台红2号、台红3号与南宁蜜龙,用数量性状聚类分析时,却无法聚为同一群组。整体上看两种聚类方法得出的结果存在较大差异,在实际使用中要根据不同育种目的结合两种分析结果进行选择。
3 结论与讨论
种质资源鉴定是育种工作的基础。本研究应用ISSR分子标记和数量性状变异分析两种方法,均验证了火龙果不同种质遗传背景的较大差异。7个ISSR引物平均扩增检测出13.8个位点,多态性比例达95.88%,为高度多态性,与前人研究基本一致[12]。性状变异是植物品种形成的前提,变异系数一方面反映了某一性状的可塑性,同时也反映了该性状的稳定程度。本研究种质间各数量性状的变异系数在12.35%~51.66%之间,显示出丰富的遗传多样性,与黄凤珠等[16]在火龙果花表型性状多样性的研究结果一致。火龙果植株无叶片,靠攀援生长的茎蔓完成光合作用,茎蔓一般呈三棱形至五棱形,可形成气生根,棱边有刺座,沿棱边排列,刺座上有短刺,刺座也是芽点。不同品种茎蔓表现差异较大。火龙果自然变异丰富,不少栽培品种的选育来自自然条件下的芽变选种,经过长期自然变异和人工选择,形成极为丰富的数量性状变异。火龙果果实呈圆形至长椭圆形,部分不规则,因品种不同而异。果皮上着生的鳞片因品种不同而形态各异,数量也有较大变异。在所观测数量性状中,棱的厚度、单果重、鳞片长度变异系数超过40%,说明其稳定性较差,易受外界环境影响。极高的变异系数也间接说明火龙果虽然育种研究时间短,商业化品种少,但种质资源却极为丰富。
聚类分析的实际应用,除了追溯物种起源外,在育种中主要为亲本选择提供重要参考,避免选择亲缘关系较近的种质作为杂交配组,以提高育种效率。根据ISSR分子标记和数量性状的聚类分析,虽然有部分相似,但整体结果并不一致,这与其他作物的研究结果一致[19],说明表型虽受基因组控制,但其表达也受环境因素影响。本研究将分子标记和表型性状相结合,兼顾分子水平与环境影响的结果,能为育种工作中亲本的精准选择提供依据。
猜你喜欢
今日农业(2022年13期)2022-11-10
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年2期)2022-03-29
中老年保健(2021年6期)2021-11-30
今日农业(2021年13期)2021-11-26
今日农业(2021年16期)2021-11-26
昆明医科大学学报(2021年5期)2021-07-22
昆明医科大学学报(2021年2期)2021-03-29
红蜻蜓(2017年6期)2017-10-30
中国民族民间医药·下半月(2014年2期)2014-09-26
