
赤皮青冈不同家系苗期生长和叶片性状遗传变异*
2022-10-26 01:11周燕黄盛怡杨孟晴王斌王秀花张东北吴小林周志春
西部林业科学 2022年5期
周燕,黄盛怡,杨孟晴,王斌,王秀花,张东北,吴小林,周志春
(1.浙江省建德市林业总场,浙江 杭州 311400;2.中国林业科学研究院亚热带林业研究所, 浙江省林木育种技术研究重点实验室,国家林业局马尾松工程技术研究中心,浙江 杭州 311400;3.浙江省庆元县实验林场,浙江 庆元 323800)
林木遗传变异是遗传改良的基础,多层次丰富的遗传变异决定了遗传改良的潜力[1-2]。林木性状的遗传变异大量存在于家系间及个体间,深入研究家系性状的遗传变异规律,有助于制定科学的育种策略[3-4],为亲本选择、杂交组合配制以及遗传改良提供有效的信息[5]。研究林木遗传变异最简单和直观的方法是分析林木生长性状的差异性,苗高和地径是林木苗期最直观的质量指标,通过苗木质量指标可在一定程度上预测其造林后的生长表现[6];叶片是植物进行光合作用和蒸腾作用的主要器官,叶片的数量和大小与植株的生长有着直接的关系,常作为衡量植物生长发育的主要观测指标[7]。鉴于苗高、地径和叶片性状对植株生长发育的重要意义,很多研究将其用于林木苗期遗传变异分析与选择[8-10]。
赤皮青冈〔Cyclobalanopsisgilva(Blume)Oerst〕为壳斗科(Fagaceae)青冈属常绿阔叶大乔木,是我国亚热带地区重要的珍贵用材和生态修复树种。不同学者对赤皮青冈的地理分布[11-12]、群落结构[13]和遗传多样性[12-13]进行研究,探讨赤皮青冈容器苗培育[14]、组织培养相关技术[15],揭示幼苗在光照、干旱和水淹胁迫条件下的生理特性[16-17]和叶片解剖结构[18]。然而关于赤皮青冈种质资源收集评价和良种选育的工作才刚刚起步,其生长和叶片性状遗传变异规律尚不清楚,这制约赤皮青冈遗传改良研究深度。本研究以来自福建、湖南和江西等地的52个赤皮青冈优树自由授粉家系1 a生容器苗为材料,研究不同种源和家系生长和叶片性状的遗传变异规律,估算各性状遗传参数和遗传相关性。在此基础上,对不同家系进行聚类分析并评价各类群特点,以期为赤皮青冈优良家系的选择提供参考,为其遗传改良的推进提供理论依据。
1 材料与方法
1.1 试验材料和设计
试验在浙江省庆元县实验林场(119°01′25″E,27°38′48″N)上坑良种基地苗圃地钢构大棚内进行,该地海拔510 m,属亚热带季风气候,温暖湿润,四季分明,年平均气温17.6 ℃,年降水量1 721.3 mm,无霜期245 d。
试验材料为1 a生赤皮青冈优树自由授粉家系容器苗。2018年在福建建瓯(FJJO),湖南城步(HNCB)、会同(HNHT)、永顺(HNYS)和江西婺源(JXWY)等地的赤皮青冈优良天然林中,选择树干通直圆满,无病虫害,高径生长明显大于周围3~5株对比木的优树,采集其成熟种子。不同种源家系数量分别为:FJJO 32个、HNCB 8个、HNHT 6个、HNYS 3个、JXWY 3个,共52个家系。2019年3月在庆元县实验林场采用随机完全区组设计进行育苗,每个家系每小区育苗20株,3次重复。育苗容器规格4.5 cm×10 cm,基质为泥炭+谷壳+珍珠岩(体积比为5︰3︰2),苗木培育按一般生产性常规水肥管理进行。
1.2 试验方法
2019年11月对所有赤皮青冈家系容器苗生长和叶片性状进行测定,生长性状包括苗高(H)和地径(D),叶片性状包括叶片数(N)、最大叶片长(L)和最大叶片宽(W)。苗高为从苗干基部的土印处到主干顶芽基部的长度,采用卷尺测量;地径采用十字交叉法,用游标卡尺测量土印处的直径作为地径;叶片数为全株叶片数量;最大叶片长和最大叶片宽分别为全株最大叶片的长度和宽度,采用钢尺测量。
1.3 数据分析
性状分析使用混合线性模型,其表达式如下。
Yijk=μ+Bi+Fj+BFij+Eijk
式中:Yijk为第i区组第j家系第k单株的观测值;μ为群体平均值;Bi为第i区组效应;Fj为第j家系的效应;BFij为第i家系和第j区组的互作效应;Eijk为机误。此外,μ和Bi为固定效应,其余为随机效应。采用SAS 8.0软件的MEANS过程进行描述性统计,GLM过程进行性状方差分析,CORR过程计算表型相关,PROC VARCOMP过程中的TYPE1方法计算各方差分量,进而估算赤皮青冈不同家系各性状遗传参数[6]。聚类分析通过R 3.6 软件完成。
2 结果与分析
2.1 赤皮青冈优树家系生长和叶片性状的总体变异
赤皮青冈优树家系1 a生苗高和地径分别为33.67 cm和0.48 cm,叶片数、最大叶片长和最大叶片宽分别38.34片、7.53 cm和2.38 cm(表1),不同家系间差异极显著(P<0.01),表明家系间存在着丰富的遗传变异,这为优良家系的选择提供足够的空间。苗高、地径、叶片数、最大叶片长和最大叶片宽最大的家系分别高出最小家系83.15%、55.56%、120.90%、29.14%和50.78%,叶片数的变幅最大,苗高次之,最大叶片长变幅最小,即叶片数和苗高变异空间相对较大。赤皮青冈优树家系各性状表型变异系数均大于遗传变异系数,苗高和叶片数分别在生长和叶片性状中变异最大,这为确定其优良家系发掘利用提供了依据。
赤皮青冈优树生长和叶片性状的家系和单株遗传力均较高,除最大叶片长和最大叶片宽的单株遗传力相对较低(0.22和0.38),受中等遗传控制外,其它性状家系和单株遗传力均高于0.4,受较强遗传控制,意味着苗期生长和叶片性状的潜在改良效果较好。比较发现,各性状家系遗传力和单株遗传力均为叶片数>苗高>地径>最大叶片宽>最大叶片长,且各性状的家系遗传力均大于单株遗传力,说明苗期的赤皮青冈以家系选择更有效。
2.2 赤皮青冈优树的苗期生长和叶片性状的种源差异
不同种源赤皮青冈1 a生容器苗生长和叶片性状差异显著(P<0.01或P<0.05),见表2。

表1 赤皮青冈苗期生长和叶片性状变异分析Tab.1 Variation analysis of growth and leaf traits of C.gilva at seeding stage

表2 不同种源赤皮青冈苗期生长和叶片性状差异Tab.2 Difference of growth and leaf traits of C.gilva from different provenances at seeding stage
从生长性状来看,苗高和地径最大的种源分别高出最小种源54.86%和30.00%,苗高的种源变异大于地径。5个种源中HNCB的苗高最高(36.30 cm),与HNHT和FJJO无显著差异,但显著高于HNYS和JXWY;HNYS的地径最大(0.52 cm),与HNCB和FJJO无显著差异,但显著高于HNHT和JXWY;JXWY的苗高(23.44 cm)和地径(0.40 cm)均最低,显著低于其他4个种源。从叶片性状来看,叶片数、最大叶片长和最大叶片宽最大的种源分别高出最小种源74.08%、12.70%和17.29%,叶片数的种源变异较大。FJJO的叶片数最多(42.44片),显著高于其它4个种源;HNHT的最大叶片长(7.90 cm)和HNCB的最大叶片宽(2.51 cm)相对较高;JXWY的3个叶片性状为5个种源中最低。
不同种源赤皮青冈的苗期生长和叶片性状的表型变异系数总体差异不大,仅HNYS叶片数的表型变异系数(63.82%)明显高于其他种源(图1)。不同种源赤皮青冈的苗期生长和叶片性状的遗传变异系数差异均较大,以HNCB的苗高(10.98%)、叶片数(14.99%)和最大叶片长(7.86%),JXWY的地径(11.74%)和HNHT的最大叶片宽(15.09%)遗传变异系数最大,是相应最小值的2~7倍,可见种源间存在着丰富的遗传变异。
不同种源赤皮青冈的苗期生长和叶片性状的家系遗传力,除JXWY的苗高(0.13)和叶片数(0.26)、FJJO和HNYS的最大叶片长(分别为0.24和0.13)相对较低外,其余不同种源赤皮青冈的苗期生长和叶片性状的家系遗传力均在0.4以上,受较强遗传控制(图1)。单株遗传力的种源差异较家系遗传力明显,HNCB的苗高(0.72)、叶片数(0.52)和最大叶片长(0.64),JXWY的地径(0.75)和HNHT的最大叶片宽(0.98)单株遗传力相对较高,遗传改良潜力较大;JXWY的苗高(0.01)和叶片数(0.02),FJJO、HNYS和HNHT的最大叶片长(分别为0.03、0.02和0.06)单株遗传力较低,遗传改良潜力相对较小。

图1 不同种源赤皮青冈的苗期生长和叶片性状遗传差异Fig.1 Genetic difference of growth and leaf traits of C.gilva from different provenances at seeding stage
2.3 赤皮青冈优树家系的苗期生长与叶片性状间的相关性
赤皮青冈优树家系的苗高与地径之间呈极显著的表型和遗传正相关(P<0.01)(表3),其表型和遗传的相关系数分别为0.283和0.604。除地径与最大叶片长遗传相关不显著外,苗高和地径与叶片性状之间均呈显著或极显著的表型与遗传正相关(P<0.01或P<0.05)。叶片数与最大叶片长和最大叶片宽的表型和遗传相关性均不显著,最大叶片长与最大叶片宽的表型和遗传相关性均呈极显著正相关(P<0.01),表明最大叶片长和最大叶片宽与叶片数量关系不大,但叶片的长和宽存在着紧密联系,叶片越长相应地也越宽。总的来看,赤皮青冈的苗期生长性状与叶片性状关系密切,生长快的家系叶片数量多、叶片长和宽较大。

表3 赤皮青冈苗期生长和叶片性状间的表型与遗传相关Tab.3 Phenotypic and genetic correlation analysis of growth and leaf traits of C.gilva at seeding stage
2.4 赤皮青冈家系的苗期聚类分析
对52个赤皮青冈家系的苗期生长和叶片性状进行聚类分析,在欧式距离为15处,可将其归为3大类(图2)。第Ⅰ类为来自HNCB(HNCB-5、HNCB-6、HNCB-7)、HNHT(HNHT-5)和FJJO(FJJO-5、FJJO-9、FJJO-18、FJJO-19)3个产地的8个家系,占比15.38%,其苗高、地径、叶片数、最大叶片长和最大叶片宽分别高出整体均值的15.21%、10.43%、17.71%、2.96%和2.88%,为苗期速生性较强家系;第Ⅱ类为来自FJJO、HNCB、HNHT和HNYS等4个产地的41个家系,占比78.58%,苗高、地径、叶片数和最大叶片长分别低于整体均值的0.74%、0.87%、0.79%和0.07%,仅最大叶片宽高出整体均值的0.16%,为苗期速生性较中等家系;第Ⅲ类仅有JXWY产地的3个家系,占比5.77%,其苗高、地径、叶片数、最大叶片长和最大叶片宽分别低于整体均值的30.37%、15.86%、36.42%、6.86%和9.90%,为苗期生长较慢家系。对苗期进行优良家系选择时,第Ⅰ类群中的8个家系均可选为苗木速生优良家系,第Ⅱ类群中部分表现突出的家系也可以入选。
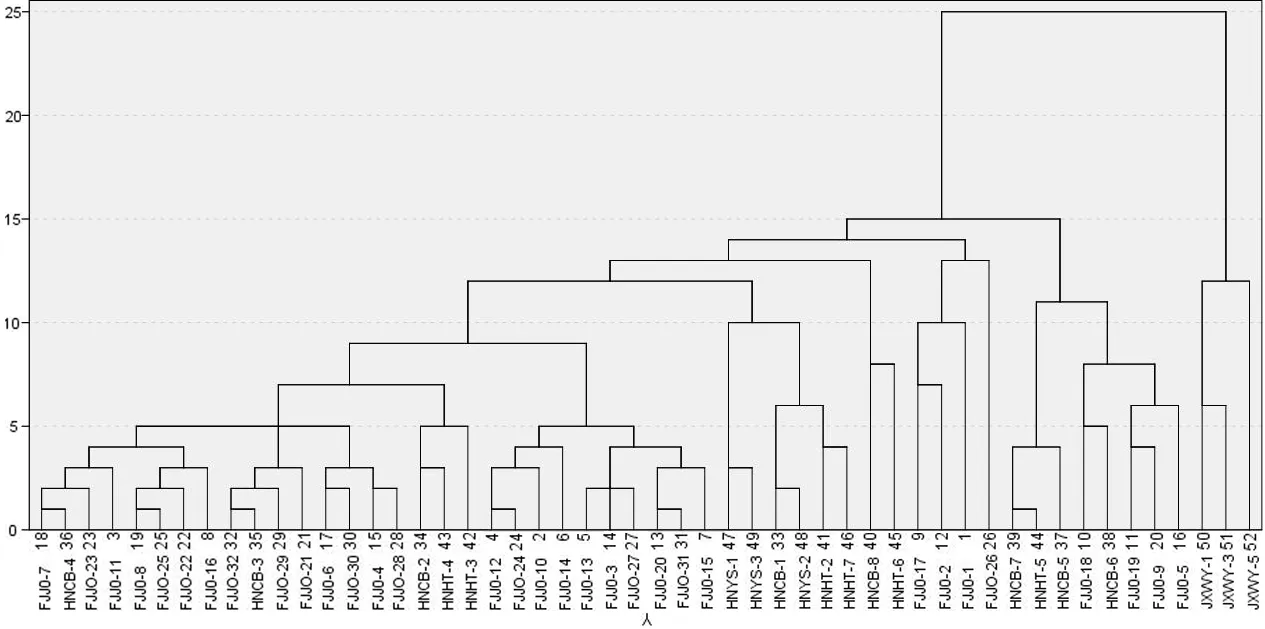
图2 赤皮青冈优树家系的苗期聚类Fig.2 Cluster analysis of C.gilva families
3 讨论与结论
林木遗传变异是选择育种的前提,因生长环境不同,长期的自然选择会形成不同的变异,同一树种在种源间、林分间、个体间及个体内均存在遗传变异[19]。本研究赤皮青冈优树的苗期生长和叶片性状表型变异系数为17.68%~42.14%,遗传变异系数为4.17%~18.13%,各性状家系间差异均达极显著水平,显示种源/家系间存在广泛的变异,遗传改良选择空间较大。研究表明,麻栎(Quercusacutissima)不同种源苗高和地径的变异系数(标准差/均值)分别为8.28%~17.75%和11.53%~34.27%,栓皮栎(Q.variabilis)不同种源苗高和地径的变异系数分别为9.36%~26.67%和12.43%~32.47%[20]。本研究赤皮青冈不同种源/家系苗高和地径变异系数(标准差/均值)为11.22%和8.63%,与麻栎和栓皮栎相比,苗高的变异接近,地径的变异要小。
遗传力大小反映林木性状变异受遗传制约的程度,遗传力的估算对于育种策略的制定具有重要的指导意义[21]。本研究赤皮青冈优树的苗期生长和叶片性状家系遗传力在0.71~0.90之间,受较强遗传控制,说明其苗高、地径、叶片数、最大叶片长和最大叶片宽等性状主要受自身遗传条件制约,受培育条件和环境效应影响较小。长期的地理隔离,使赤皮青冈不同种源的遗传结构及适应性发生了改变,从而具有较强的种源选择能力[9],这一结果与不同学者对刨花润楠(Machiluspauhoi)[9]、火力楠(M.macclurei)[22]、杉木(Cunninghamialanceolata)[23]及落叶松(Larixgmelinii)[24]等树种的研究结论一致。
以苗高和叶片数对赤皮青冈52个家系进行聚类,可将其分为苗期速生性较强家系、苗期速生性中等家系和苗期生长较慢家系3类,这为赤皮青冈苗期选择提供了一定参考。考虑到本研究采用的是1 a生容器苗,聚类结果有待同批次子代测定林试验加以验证。因此,后续需对赤皮青冈优树家系进行多点多年跟踪观测,通过子代测定分析各生长性状在苗期和幼成林间的早晚相关性和稳定性,以进一步分析赤皮青冈苗期选择的实用性和可靠性。林木育种的长期目标是获得可持续的遗传增益,本研究仅仅是针对苗期生长和叶片性状进行分析,后期可通过扩大种源及家系来源,结合区域化造林试验和子代测定,为赤皮青冈造林选育出高产、优质、稳定和适宜的优良家系。此外,还应对赤皮青冈的其它性状进行研究,掌握不同性状的遗传变异规律,为赤皮青冈的遗传改良提供全面的理论依据。
猜你喜欢
特产研究(2022年6期)2023-01-17
中华实用诊断与治疗杂志(2022年1期)2022-08-31
辽宁林业科技(2021年1期)2021-03-16
吉林林业科技(2018年6期)2018-11-21
武夷学院学报(2017年9期)2017-11-29
报刊荟萃(上)(2017年7期)2017-07-16
魅力中国(2016年29期)2017-05-27
西藏科技(2016年9期)2016-09-26
江苏农业科学(2016年6期)2016-07-25
广西林业科学(2016年3期)2016-03-16
