
对叶百部种子生物学特性研究
2022-11-28 13:09王红兰杜玖珍蒋桂华孙洪兵蒋舜媛朱文涛
种子 2022年10期
连 艳, 王红兰, 杨 萍, 杜玖珍, 蒋桂华, 孙洪兵, 周 毅, 蒋舜媛, 朱文涛
(1.成都中医药大学 药学院,西南特色中药资源国家重点实验室,中药材标准化重点实验室, 四川 成都 611137;2. 四川省中医药科学院, 成都 610041)
对叶百部(StemonatuberosaLour.)为百部科百部属多年生攀缘藤本植物,以块根入药,是传统中药百部的基源物种之一,也是东南亚各国的传统药用植物[1]。百部具润肺、下气止咳、杀虫灭虱的功效,是中医治疗肺系疾病及临床杀虫的常用药,此外百部在植物源杀虫剂领域也具有较大的利用和开发前景[2-4]。对叶百部因资源分布广,临床效果好,目前已成为百部药材主要来源[5-6]。近年来,在市场需求的刺激下,对叶百部野生资源遭受掠夺式采挖,资源储量锐减,市场供求矛盾日益凸显。人工种植替代野生资源是实现植物资源可持续利用的有效途径。然而对叶百部种子大田出苗率低,部分地区甚至采用分株繁殖方式,这种方式成本高、效率低,导致人工种植产业发展缓慢。陈玉菡等[7]野外调查表明,对叶百部产种潜力较大,因此利用种子繁殖是保障人工种植产业发展的理想育苗方式。
种子是农业生产中最基本的生产资料,开展种子学的相关研究对植物的栽培利用具有重要意义。目前有关对叶百部的研究主要集中在化学、药理、品质评价等方面[8-14],在种子学方面研究较少,仅陈玉菡等[15]研究了对叶百部种子的发芽特性,有关对叶百部种子的生物学特性缺乏系统研究。因此,本研究从种子表型特征、吸胀特性、萌发特性、储藏特性和休眠特性等方面对对叶百部种子生物学特性开展系统研究,旨在为对叶百部种子鉴别、质量评价及高效育苗等领域研究提供理论基础。
1 材料和方法
1.1 供试材料
实验用种分别于2019年、2020年和2021年的6月下旬,采自四川省成都市新都区龙桥镇对叶百部野生引种基地,野生种源来源于四川省达州市大竹县,由四川省中医药科学院周毅研究员鉴定为对叶百部(S.tuberosa)。种子去除杂质后晒干,随后置于10 ℃恒温培养箱中保存。于2021年10月至11月集中开展实验。
1.2 试验方法
1.2.1种子形态特征观测
取2021年采收的对叶百部种子,于体视显微镜下观察种子的形状、颜色、表面纹理及解剖结构,并采用精度为0.01 mm的游标卡尺随机测量100粒种子的长、种柄长、直径等形态学指标。
1.2.2种子含水量与千粒重
取2021年采收的对叶百部种子,参照国家标准(GB/T 3543—1995)《农作物种子检验规程》[16],采用高温烘干法测定含水量,采用百粒称重法测定千粒重。
1.2.3种子生活力检测
取3个不同年限采收的对叶百部种子,自来水中浸泡24 h,随后沿中轴线将种子纵向切开,取其中一半浸泡于0.2% TTC溶液后置于30 ℃生化培养箱中避光染色6 h,染色结束后用清水漂洗、镜检。所有处理设置3个生物学重复,每个重复50粒种子。
1.2.4种子吸水特性
取2021年采收的对叶百部种子两份,一份保留完整种皮,另一份自胚根端剥除少量种皮,露出内部种仁。称量种子初始重量,随后将种子置于自来水中浸泡,分别在浸泡2、4、6、8、10、12、18、24、30 h时取出种子,用吸水纸吸干表面水分称重。每个处理设置3个生物学重复。
种子吸水率(%)=[(Wt-W)/W]×100%,其中,Wt为种子浸泡t时间后的重量,W为种子初始重量。
1.2.5种子发芽试验
温度对种子萌发的影响。取2021年采收的对叶百部种子,自来水中浸泡24 h,随后将种子与洗净的湿河沙混合,平铺于发芽盒底部。将发芽盒分别置于10、20、25、30、35、40 ℃的恒温培养箱内开展发芽实验。
种皮对种子萌发的影响。取2021年采收的对叶百部种子,自来水中浸泡24 h,随后将种子分成两份,一份保留完整种皮,另一份自胚根端剥去少量种皮,露出少量种仁。将种子与洗净的湿河沙混合,平铺于发芽盒底部,置于30 ℃的恒温培养箱内开展发芽实验。
储存时间对种子萌发的影响。取3个不同年限采收的对叶百部种子,自来水中浸泡24 h,随后将种子与清洗干净的湿河沙混合,平铺于发芽盒底部,置于30 ℃的恒温暗培养箱内开展发芽实验。
以上所有发芽实验每处理设置3个重复,每个重复50粒种子。实验期间每隔2 d观察种子发芽情况,并翻动沙子以防止种子生霉。种子发芽以胚根生出0.5 cm为标准,第30天结束实验,统计发芽率和发芽势。
发芽率(%)=(种子萌发总数/供试种子总数)×100%;
发芽势(%)=(N20/供试种子总数)×100%,式中,N20为实验前20 d的萌发种子总数;
发芽时滞(d):第一粒种子发芽需要的天数。
1.3 数据处理
数据采用Excel 2018软件进行数据处理分析及绘图,运用SPSS 17.0软件进行差异显著性分析(p<0.05)。
2 结果与分析
2.1 种子形态特征
对叶百部种子呈棕褐色至黑褐色,主体呈椭圆形至卵圆形,表面具纵槽纹14~19条,无裂隙,发芽口隐藏不可见。种子基部具细长种柄;顶端内脐明显,呈瘤状突起;种脊线形,自种柄基部延伸至顶端内脐(图1 A,B)。种子长(含种柄)7.72~12.39 mm,直径2.09~3.84 mm,种柄长1.42~3.70 mm,三者变异系数均超过11%,其中种柄长的变异系数最大,达到20.05%(表1)。
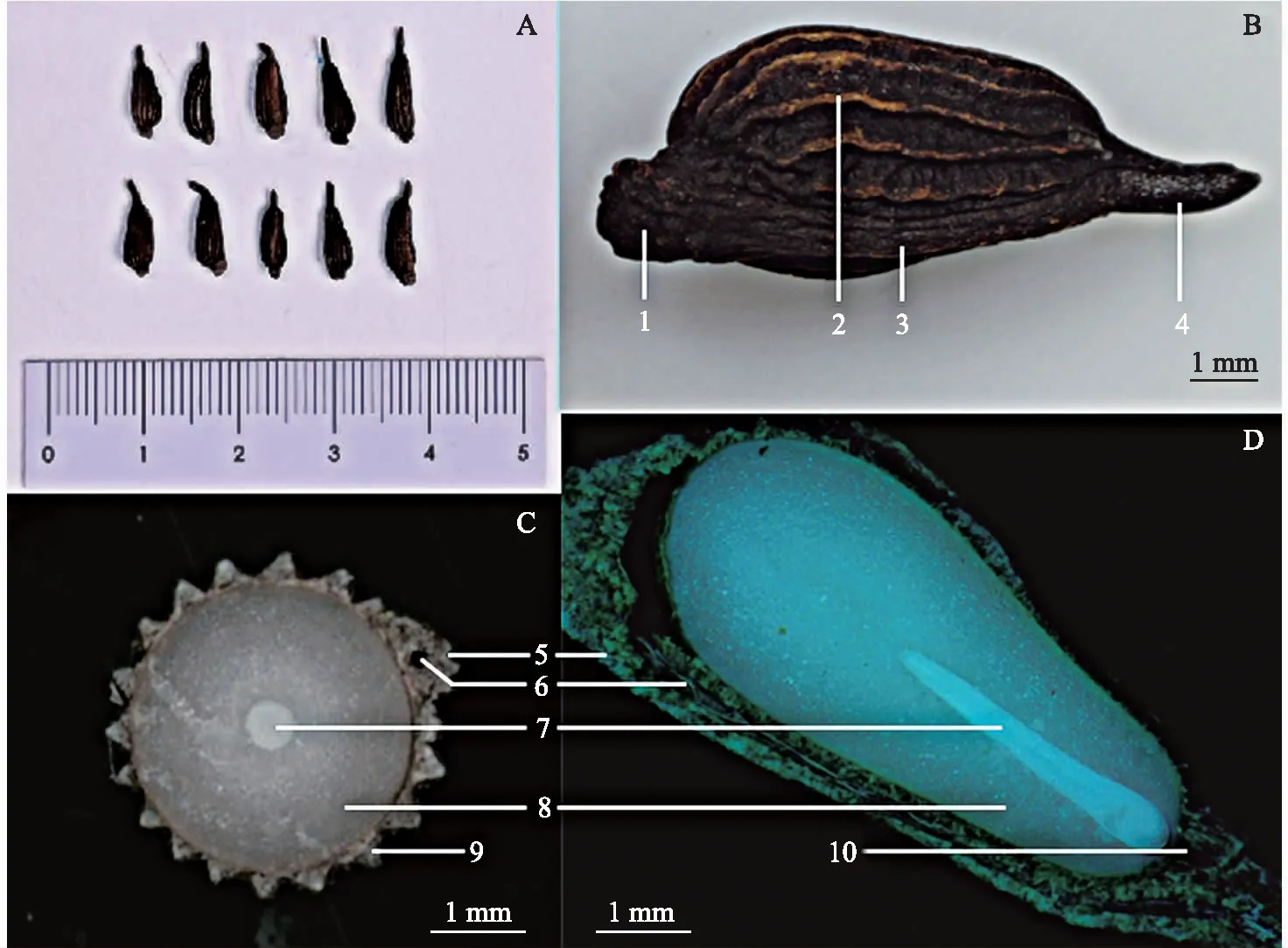
注:A、B为种子形态特征;C为种子横切面;D为种子纵切面。1为内脐;2为种皮表面纵槽纹;3为种脊;4为种柄; 5为种脊横/纵切面;6为种脊内部管状通道及丝状纤维残留;7为种胚;8为胚乳;9为种皮横切面;10为发芽口。 图1 对叶百部种子形态特征及解剖结构 Fig.1 The morphological characteristics and anatomical structure of S. tuberosa seeds
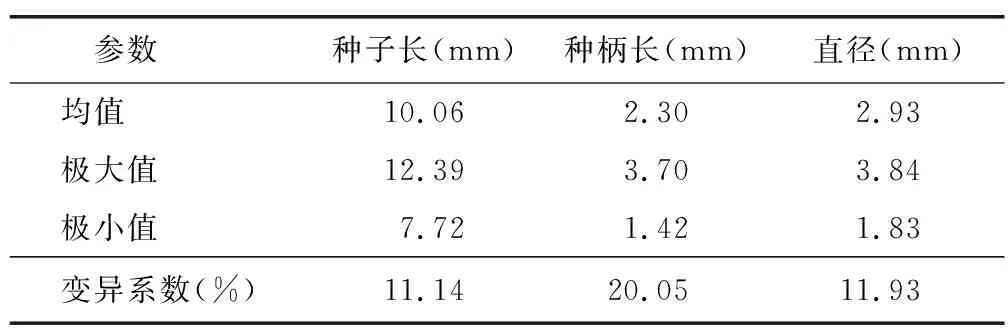
表1 对叶百部种子形态指标分析Table 1 The morphological index of S. tuberosa seeds
种子解剖结构观察可知(图1 C,D),对叶百部种皮规律性增厚使种子横切面呈齿轮状,增厚部分内部呈海绵状;种脊内部可见一管状通道及丝状纤维残留;胚根端种皮纵切面可见隐藏的发芽口。对叶百部种子具白色的胚和胚乳,胚乳丰富;种胚条状,着生于基部胚乳中央,属典型的直立型胚,长约占种仁的1/2。
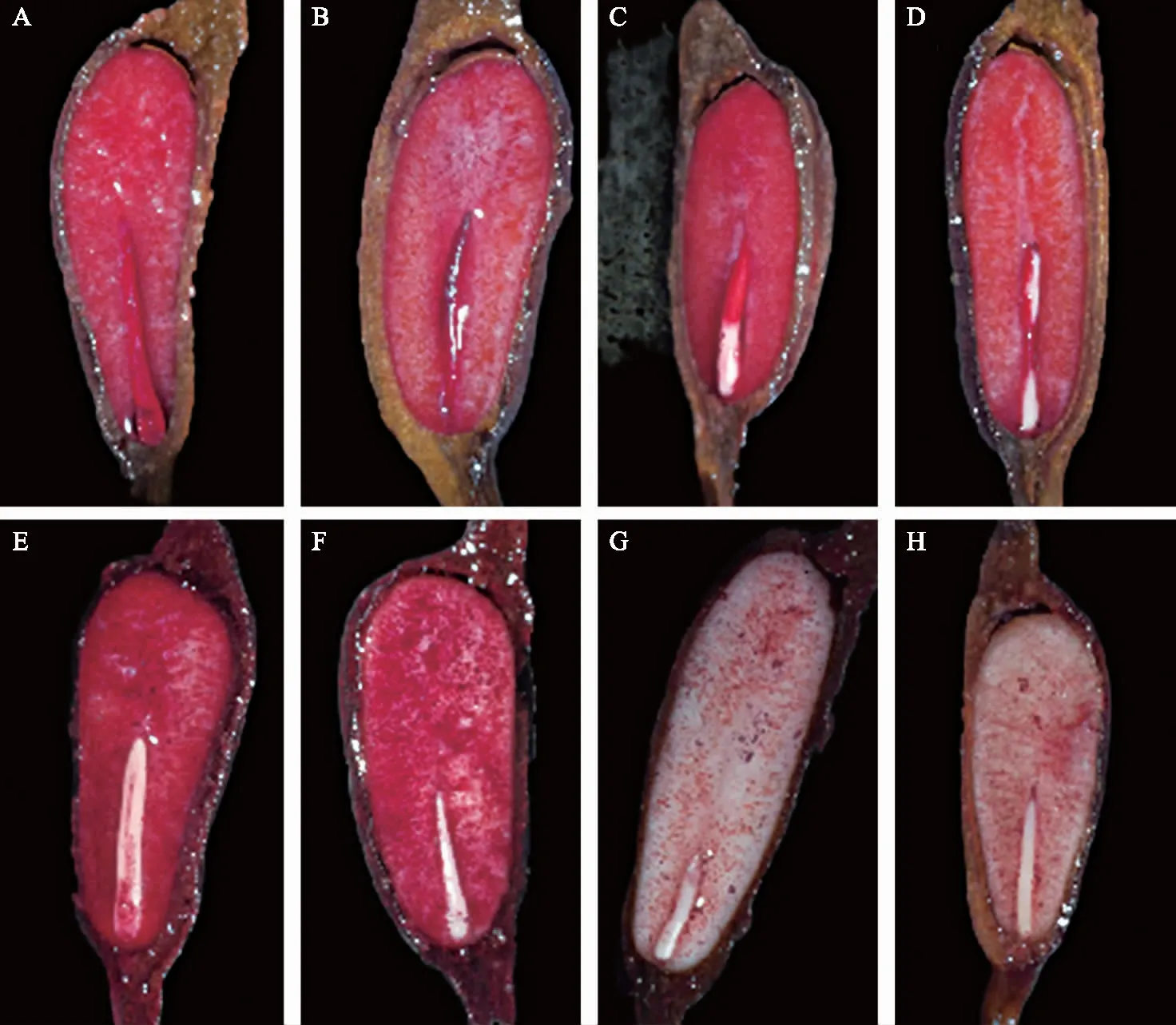
注:A、B为胚●+胚乳●;C、D为胚+胚乳●;E、F为胚○+胚乳●;G、H为胚○+胚乳○。 图2 对叶百部种子TTC染色模式 Fig.2 Pattern of TTC staining on S. tuberosa seeds
2.2 种子含水量与千粒重
自然晒干的对叶百部种子含水量为14.30%,在该含水量条件下的种子千粒重为25.31 g,依据作物种子千粒重分级标准可将对叶百部种子归为中粒种子。
2.3 种子生活力
采用TTC染色法测定对叶百部种子生活力,有活力种子的胚和胚乳均能被染色,且呈现4种染色模式:“胚+胚乳●”改为“胚●+胚乳●、胚+胚乳●、胚○+胚乳●、胚○+胚乳○(图2,“●”表示全部染色,“”表示部分染色,“○”表示未染色)”;胚和胚乳的染色效果存在差异,胚染色后呈现出均一的鲜红色,胚乳染色则呈现出颗粒状或带状红斑,颜色存在鲜红和铁锈红两种。本研究以种胚2/3以上被染成红色作为种子生活力的判定标准,结果表明,对叶百部种子的生活力随储存时间的增加而急剧降低。2021年采收(储存4个月)的种子生活力可达83.33%,储存超过1年的种子生活力降低至30.67%,储存两年以上的种子则基本丧失生活力(图3)。
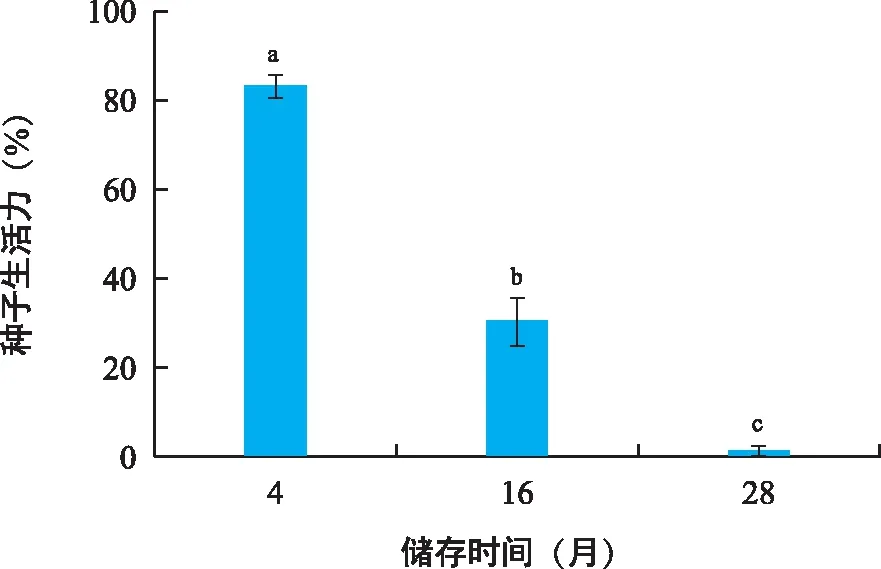
注 :不同小写字母表示差异显著(p<0.05)。图3 储存时间对对叶百部种子生活力的影响Fig.3 Effects of storage time on seed viability of S. tuberosa seeds
2.4 种子吸水特性
对叶百部种子吸水特性研究表明,无论种皮是否完整,种子吸水趋势一致,以2 h为节点分为快速吸水期(0~2 h)和缓慢吸水期(2 h以后)(图4)。快速吸水期,种皮完整种子吸水率达到44.27%,占总吸水量的59.75%;种皮破损种子吸水率为32.83%,占总吸水量的53.98%。缓慢吸水期,种皮完整种子吸水率增加29.81%,占总吸水量的40.25%;种皮破损种子吸水率增加27.99%,占总吸水量的46.02%。以吸水率增量同浸泡时间的比值评价种子吸水速率,种皮完整种子快速吸水期的吸水速率达到22.14%/h,显著高于种皮破损种子的吸水速率(16.44%/h,p<0.05);种皮完整种子和种皮破损种子在缓慢吸水期的吸水速率分别为1.06%/h和1.00%/h,两者之间差异不显著。
2.5 温度对种子萌发的影响
温度对对叶百部种子的萌发影响显著,种子发芽率和发芽势均随温度升高呈先增后降的趋势(图5)。对叶百部种子的最佳发芽温度为30 ℃,发芽率和发芽势分别为64.67%和42.67%。当温度偏离最佳发芽温度后,种子发芽率和发芽势均显著降低,其中温度为20 ℃时,发芽势降为0,发芽率仅为2.67%;当温度为35 ℃,种子发芽率降低至5.33%,发芽势降为0.67%;温度为40 ℃,发芽率和发芽势均为0。

注 :不同大写字母表示发芽率差异显著,不同小写字母表示发芽势差异显著(p<0.05)。图5 不同温度下对叶百部种子发芽率和发芽势Fig.5 Germination rate and germination potential of S. tuberosa seeds in different temperature
不同温度条件下对叶百部种子萌发进程差异显著(图6)。30 ℃和25 ℃条件下种子最先萌发,发芽时滞为14 d,35 ℃和20 ℃条件下发芽时滞分别为18 d和24 d。比较不同温度下日萌发种子数可知,30 ℃条件下种子萌发进程最快,日萌发种子数呈现先增后减趋势,并在第20天达到峰值。
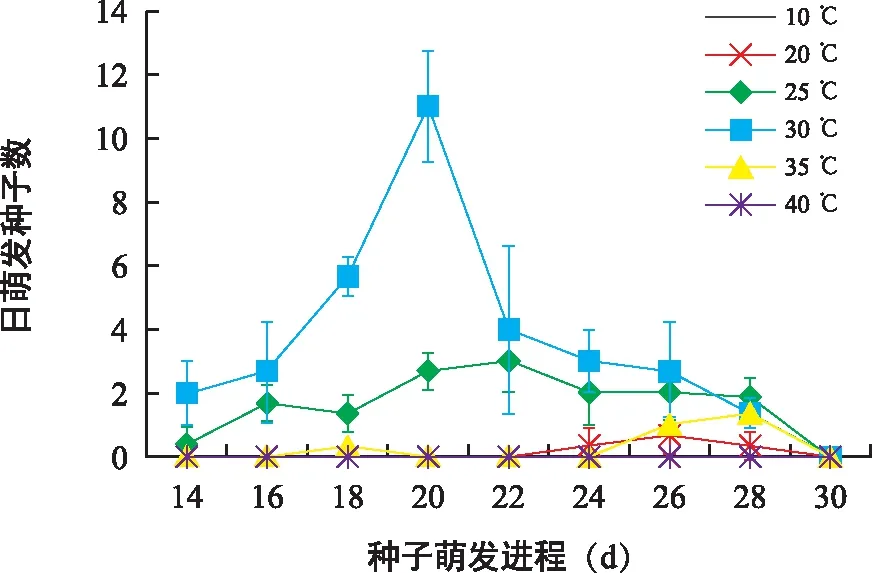
图6 不同温度条件下对叶百部种子萌发进程Fig.6 Germination process of S. tuberosa seeds in different temperatures
2.6 种皮对种子萌发的影响
种皮对对叶百部种子的萌发影响显著(图7)。种皮完整种子发芽率和发芽势分别为59.33%和38.00%;剥除胚根端种皮的种子,种子发芽率和发芽势增加到86.67%和80.67%,较种皮完整组分别提高46.08%和112.29%。
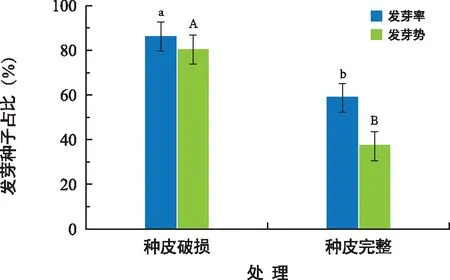
图7 种皮对对叶百部种子发芽率和发芽势的影响Fig.7 Effect of seed coat on germination rate and germination potential of S. tuberosa seeds
种皮对对叶百部种子的萌发进程影响显著,剥除胚根端的种皮后种子发芽时滞显著缩短(图8)。种皮完整的种子发芽时滞为12 d,在第18天日萌发种子数达到峰值,从开始萌发到日萌发种子数达峰值用时6 d;剥除胚根端的种皮后,种子发芽时滞缩短至6 d,仅为种皮完整组的1/2,并在第8天达到日萌发种子数峰值,从开始萌发到日萌发种子数达峰值仅用时2 d,为种皮完整组的1/3。
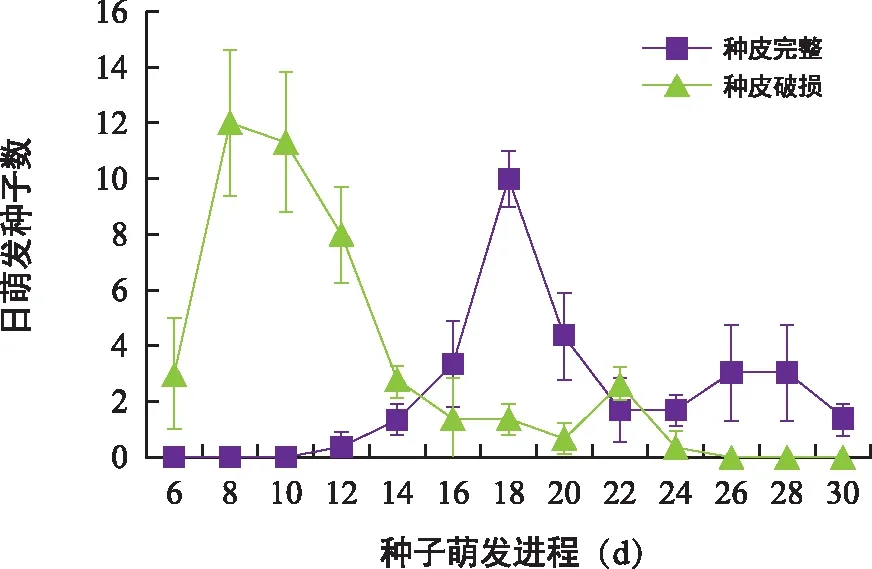
图8 种皮对对叶百部种子萌发进程的影响Fig.8 Effect of seed coat on germination process of S. tuberosa seeds
2.7 储存时间对种子萌发的影响
储存时间对对叶百部种子萌发影响显著,随着储存时间的增加,种子发芽率和发芽势均显著降低(图9)。2021年采收的种子(储存4个月)发芽率和发芽势分别为64.67%和42.67%;2020年采收的种子(储存16个月)发芽率和发芽势分别低至11.33%和7.33%,仅为2021年采收种子的1/6左右;2019年采收的种子(储存28个月)发芽率和发芽势均为0,已经丧失萌发能力。
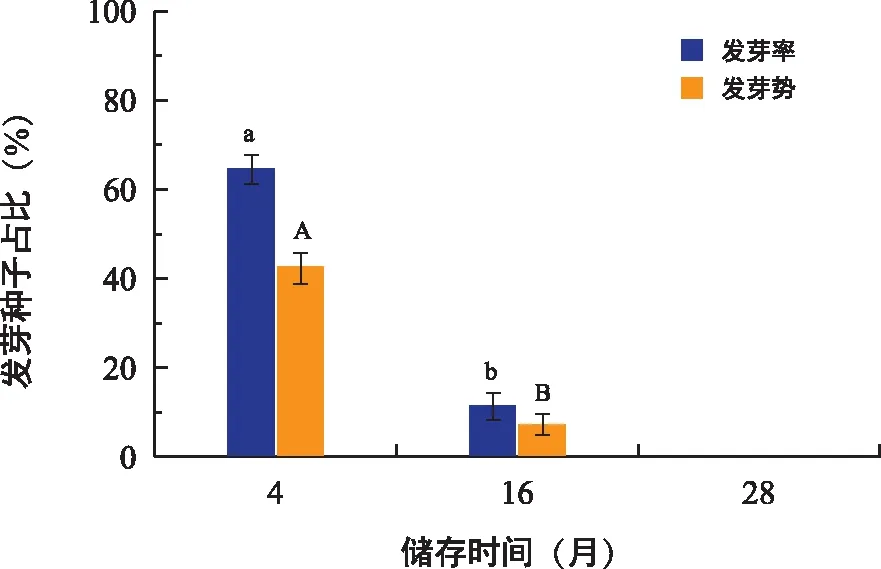
图9 储存时间对对叶百部种子发芽率和发芽势的影响Fig.9 Effects of storage time on germination rate and germination potential of S. tuberosa seeds
储存时间对对叶百部种子萌发进程的影响显著(图10)。储存4个月的种子萌发启动时间比储存16个月的种子早2 d,并且在整个种子萌发进程中,储存4个月的种子日萌发数均高于储存16个月的种子。
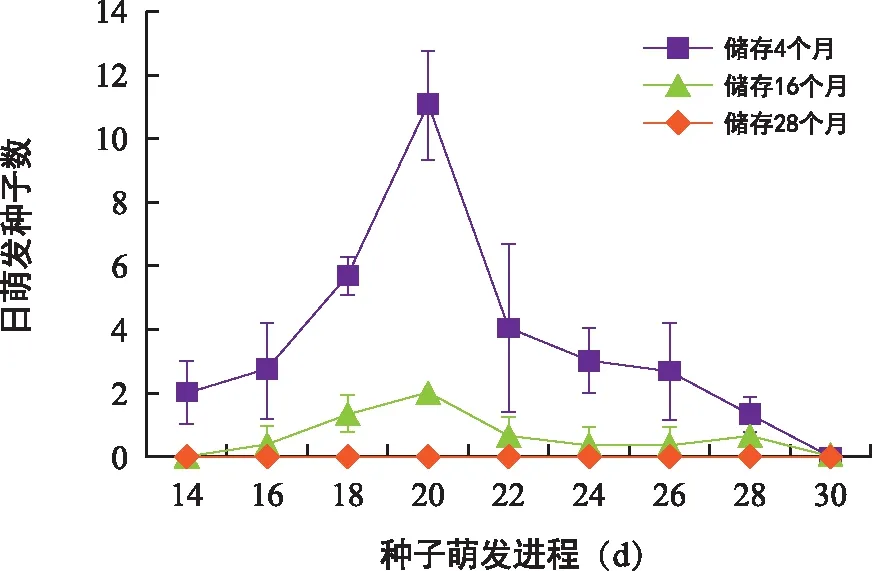
图10 储存时间对对叶百部种子萌发进程的影响Fig.10 Effects of storage time on germination process of S. tuberosa seeds
3 讨 论
种子的形态特征在遗传上呈相对稳定状态,对植物科、属、种分类和鉴定具有重要意义[17-18]。对叶百部种子表型性状特异,种皮表面不仅具有独特的纵向槽纹,同时具有丰富的胚珠遗迹,包括种柄、内脐、种脊及隐藏的发芽口等结构。以上的形态、结构特征均可作为种属鉴定的重要依据。同叶、花、果等其他器官组织一样,种子的形态特征在种内同样存在着变异,在一定程度上可反映种群的遗传多样性[19]。对叶百部种子形态学指标变异较大,变异系数均超过11%,最大达到20.05%,表明本实验样品采收区的对叶百部种群内部遗传多样性丰富。
温度是种子萌发必要的外部条件之一,温度过高或过低会使膜的透性、膜结合的活性和酶变性而影响萌发[20-21]。对于多数种子植物来说,种子适宜发芽温度具有一定范围,偏离最适发芽温度5~10 ℃仍然可以保持较高的发芽率和发芽势[22-23]。而本研究表明,对叶百部种子的萌发对温度极度敏感,低于或高于最适温度后种子的发芽率和发芽势均急剧下降,甚至不发芽,表现出对温度的狭窄适应性。这一现象在濒危植物中较为常见,星油藤[24]、海南龙血树[25]、合柱金莲木[26]、红榄李[27]等濒危植物的种子萌发均表现出对温度不同程度的狭窄适应性。对叶百部种子萌发对温度的狭窄适应性预示着野外条件下其种子的萌发率低,种群恢复和更新能力差,这也许是大量人工采挖导致对叶百部野生资源逐步枯竭的重要原因。
种子休眠是植物界中普遍存在的现象,是植物为长期有效抵抗外界恶劣环境而产生的适应性机制,对物种的延续和进化具有重要意义[28-29]。不同植物种子致眠因素不尽相同,Baskin等[30]将种子的休眠划分为生理休眠、形态休眠、形态生理休眠、物理休眠以及综合休眠等五大类。本研究中种皮完整种子在整个吸胀过程中的吸水率高于种皮破损种子,表明种皮的存在并不影响内部种仁的吸胀过程,且外部种皮的吸水性和持水性要高于内部种仁,这与对叶百部种皮增厚部分呈现海绵状结构相吻合,这一现象在山鸢尾、玉蝉花、燕子花等鸢尾属植物中同样存在[31]。后续的萌发特性研究表明,种皮完整种子在最适温度条件下并不能立即萌发,而是存在12 d左右的短暂休眠。当种子胚根端的种皮被剥除后,种子的发芽时滞缩短100%,同时发芽率和发芽势提高46.08%和112.29%。综上结果表明,对叶百部种皮具良好的透性,但对胚根的萌发存在机械阻碍,属于典型的物理休眠。
实际生产中,种子从采收到播种往往需要经过一段时间的储藏,在此过程中种子活力会逐渐降低,甚至丧失萌发能力[32]。对叶百部种子储存过程中,种子的生活力、发芽率和发芽势均随储存时间的增加而迅速降低,储存16个月后种子基本丧失萌发能力,属短命型种子[33]。在利用TTC染色法对种子生活力的测定中发现,胚被染色的种子其胚乳均被染色,而胚乳被染色的种子其胚不一定被染色,表明对叶百部种子老化过程中,胚组织先于胚乳组织丧失活力,这一现象在檀香种子中较为突出[34]。鉴于对叶百部种子不耐储存特性,因此生产中种子宜采用当年采收当年播种的方式进行育苗。
猜你喜欢
粮油与饲料科技(2022年2期)2022-11-24
安徽农业科学(2021年14期)2021-09-03
山西农业科学(2021年8期)2021-08-17
三农资讯半月报(2021年2期)2021-03-22
辽宁农业科学(2021年1期)2021-03-17
山西农业科学(2020年1期)2020-02-27
学生导报·东方少年(2019年3期)2019-05-14
小学阅读指南·高年级版(2016年9期)2016-10-31
江苏农业科学(2014年9期)2014-11-15
农家顾问(2014年4期)2014-06-26
