
喜旱莲子草入侵与湖岸带环境的耦联分析
——以城市湖泊武汉沙湖为例
2022-12-01 03:06郭佳琦赵丽娅周宇李兆华
湖北大学学报(自然科学版) 2022年3期
郭佳琦,赵丽娅,周宇,李兆华
(湖北大学资源环境学院,湖北 武汉 430062)
0 引言
喜旱莲子草(Alternantheraphiloxeroides)是苋科莲子草属多年生草本植物,俗名空心莲子草、水花生,原产于南美洲,现广泛分布于气候较为湿暖的温带-亚热带气候区[1],是首批列入《中国外来入侵物种名单》的恶性杂草.喜旱莲子草水陆皆可生长,繁殖力强、化感作用和表型可塑性强,生长迅速难以控制,能适应多种生境,排挤本土植物,是湖泊、湿地等水陆间生态系统的灾害性植物.主要的防治手段有物理防除、化学防除[2-3]、生物防治[4-6]以及综合防治措施.国内外的研究集中于全球视角下入侵风险预警和动态监测[8-9],多尺度的入侵影响分析[6,10],防治技术与应用[7]等.
尽管在农田、水域等连续分布区域的防控治理取得了一定的成效[7],但喜旱莲子草已广泛入侵城市生态系统,在城市湖泊岸线、湿地等水陆交接带以及路边生境形成斑块状的优势群落[11],对城市生态系统的生物多样性和景观美学造成损失.已有的研究表明,生境或群落的异质性显著影响了外来物种的入侵程度[11-13],而在同一群落中研究生境的异质性与植物生长特征的耦联关系,有利于了解外来物种的入侵性以及生境的可侵入性[11,14].本研究基于此类方法,通过探究城市湖泊自然环境和人工环境与入侵植物生长的耦联关系,识别对植物生存生长具有抑制作用的生境条件,以期为城市湖泊岸线的整治提升提供参考信息.
1 材料与方法
1.1 研究区概况沙湖位于湖北省武汉市,地理位置位于30°33′ N~30°34′ N、114°18′ E~114°20′ E,现有面积3.197 km2,水域面积2.711 km2,气候类型为亚热带季风气候,湖区年均气温为15.8 ℃~17.5 ℃,常年降雨量充沛,雨热同季,年降雨量在1 200 mm以上,且多集中在6—8月;冬冷夏热,春秋短,夏冬长,冬季和夏季分别长达110 d和135 d;年无霜期一般为211 d~272 d,年日照总时数一般为1 810 h ~ 2 100 h;土壤类型以红壤、黄棕壤和水稻土为主.沙湖作为城市内湖长期承担着调蓄雨洪、防洪滞涝、调节周边小气候的功能,沙湖周边城市化和人类活动程度较强,沙湖水质低于Ⅳ类标准[16],岸线环境受喜旱莲子草、凤眼蓝(Eichhorniacrassipes)侵害严重.
1.2 植物样方调查2020年8月,采用样方法对研究区进行野外调查,随机选取起点,按顺时针方向沿岸线每隔200 m设置1 m×1 m的样方1个,共设置50个样方,每个样方用GPS定位,记录其经度、纬度、海拔高度和坡度、坡向,在ArcGIS10.6中表示出样方位置(图1).识别各样方内草本层植物物种名称,测量并统计各物种的平均高度、多度、盖度和频数,对于不能确定的植物,采集标本带回实验室鉴定;用观察法描述涉及的环境因子并记录各样方自然环境和人为环境情况:自然环境因子包括湖岸沉积物、植被生境类型、土壤湿度、坡度和坡向的情况;人为环境因子包括有无护栏、驳岸、建筑和人为干扰强度的现状.植物识别参考《中国植物志》、《中国高等植物图鉴》等进行鉴定.
1.3 数据处理根据样方实际情况,用重要值(VI)描述喜旱莲子草在群落中的地位,计算公式如下
(1)
经检验,喜旱莲子草多度、盖度、高度和重要值的数据不符合正态分布,采用Spearman相关性指数检验护栏、驳岸、建筑、人为干扰强度等湖岸人为环境因子,以及湖边沉积物、土壤湿度、植被生境类型、坡度等自然生境分别与喜旱莲子草高度、多度、盖度、物种重要值的相关性.用非参数检验(Mann-Whitney、Kruskal-Wallis)分析喜旱莲子草多度、盖度、高度和重要值在不同水平的差异性.所有检验的显著性界限水平设置为0.05.所有数据使用SPSS 20.0和Excel 2016处理.
2 结果与分析
2.1 喜旱莲子草多度对岸线环境的响应沙湖沿岸喜旱莲子草多度为(46.74±8.12)株/(平均值±标准误差),由于湖泊岸线生境梯度的变化,样方中喜旱莲子草的生长指标差异性较大.从表2可看出,喜旱莲子草的多度与植被生境类型有显著正相关关系(P=0.05,r=0.29).不同生境类型的喜旱莲子草多度具有显著差异(P=0.05),水生环境中喜旱莲子草多度显著高于其他生境类型(图2).随着环境水分的增加,喜旱莲子草的多度增加,沙湖沿岸喜旱莲子草种群更倾向于生长在湿润环境.
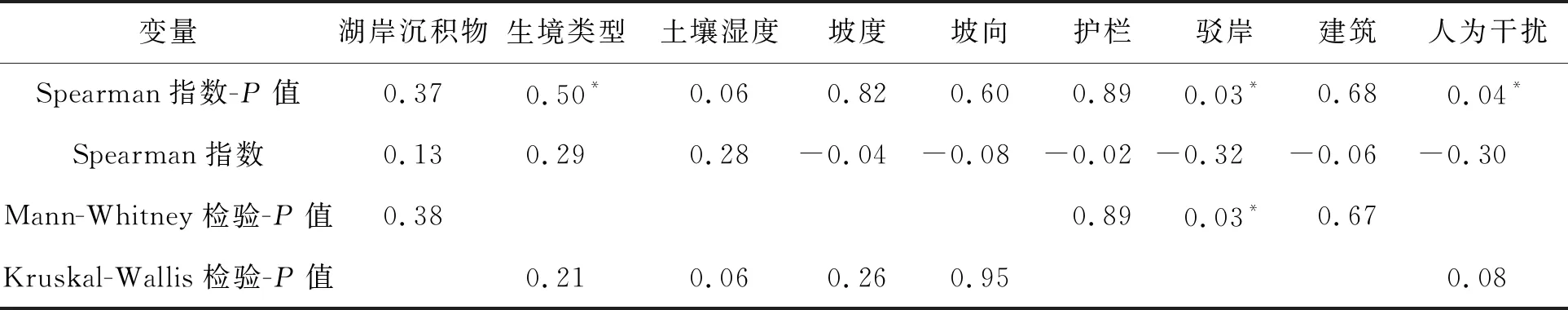
表2 喜旱莲子多度与环境因子的相关性和差异性
喜旱莲子草多度与驳岸(P=0.03,r=- 0.32)、人为干扰(P=0.04,r=- 0.30)之间存在显著负相关关系,并且有无驳岸之间喜旱莲子草多度存在显著差异(P=0.03),周围有驳岸的样方内喜旱莲子草多度相较于无驳岸减少了69%,人为增设生态驳岸可能有利于控制喜旱莲子草在湖泊沿岸的扩散.
2.2 喜旱莲子草高度对岸线环境的响应沙湖沿岸喜旱莲子草高度为(19.92±3.31)cm.从表3可以看出,喜旱莲子草高度与护栏、植被生境类型存在正相关关系,其中与生境类型的相关性最密切(P=0.00,r=0.44),水生环境的喜旱莲子草高度为(44.95±7.27)cm(图3),与其他生境类型有显著差异(P<0.01),不同水分环境的喜旱莲子草高度具有显著差异,并且随着植被水分的增加,喜旱莲子草的高度增加.护栏与喜旱莲子草高度也存在显著正相关关系(P=0.02,r=0.35),沿岸增设护栏的样方内喜旱莲子草高度显著高于无护栏的高度(P<0.01),表明沿岸护栏的增设可能利于喜旱莲子草的生长.在野外调查中,发现增设护栏的喜旱莲子草种群受外界干扰程度较小,护栏阻挡了一定的人为干扰,比如人工拔除和喷洒农药,这可能也是造成护栏显著的原因.
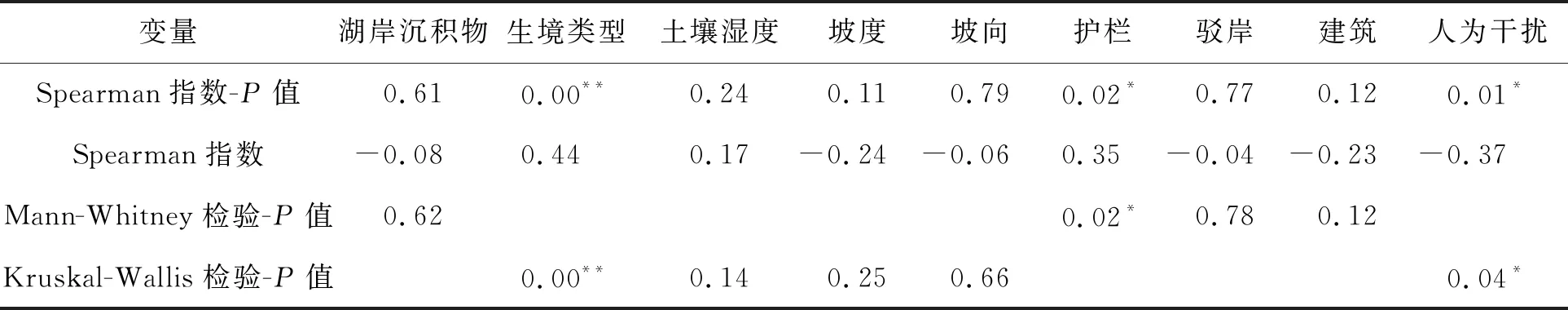
表3 喜旱莲子高度与环境因子的相关性和差异性
人为干扰强度与喜旱莲子草高度具有显著负相关关系(P=0.01,r=-0.37),不同人为干扰强度下的喜旱莲子草高度存在显著差异(P=0.04),随干扰强度的升高,喜旱莲子草的高度降低,人类活动强度的增加抑制了沙湖沿岸喜旱莲子草地上部分的伸长.
2.3 喜旱莲子草盖度对岸线环境的响应喜旱莲子草盖度为(55.13±4.13)%(图4).由表3可知,湖岸沉积物与喜旱莲子草盖度存在显著正相关关系(P=0.03,r=0.32),有无湖岸沉积物与喜旱莲子草盖度存在显著差异(P=0.03),沙湖沿岸淤泥等沉积物的存在可能利于喜旱莲子草的生长.
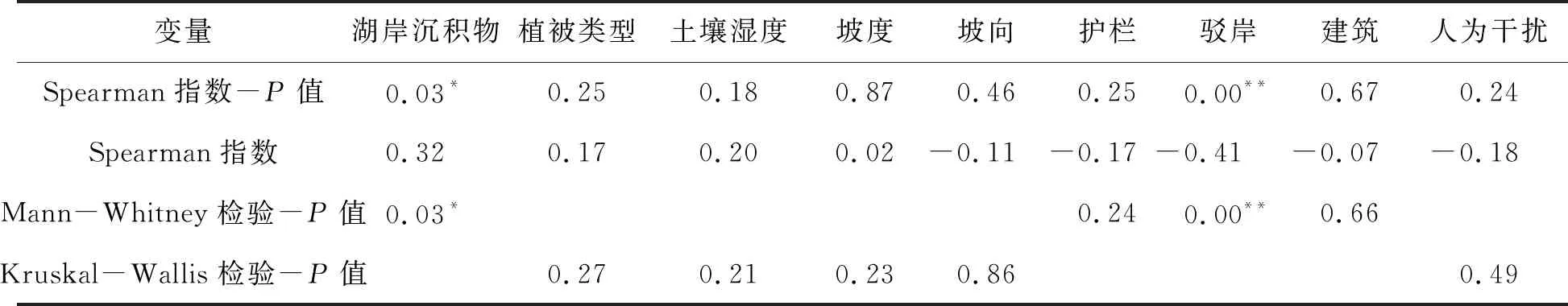
表4 喜旱莲子盖度与环境因子的相关性和差异性
驳岸的有无与喜旱莲子草盖度具有极显著负相关关系(P=0.00,r=-0.41),并且有无驳岸与喜旱莲子草盖度存在显著差异(P=0.00),实地调查中发现由于沙湖增设生态驳岸的岸线长度有限,喜旱莲子草多生长于水生环境,较陆生生境有更大的叶面积和地上生物量,可能造成了有驳岸存在的样方内喜旱莲子草盖度的平均值与无驳岸的差异不明显.
2.4 喜旱莲子草重要值对岸线环境的响应在样方中共发现草本植物18科40属43种,其中菊科、禾本科、旋花科和蓼科植物较多,分别有9种(占物种总数的20.93%)、7种(16.28%)、4种(9.3%)和4种(9.3%),其余19种占44.19%.重要值排名前5的植物为喜旱莲子草(VI=19.45)>狗牙根(VI=10.16)>双穗雀稗(VI=8.44)>马唐(VI=7.80)>结缕草(VI=6.10),这5种植物的重要值之和占群落总重要值的51.95%.
从表5可知,驳岸的有无与喜旱莲子草的重要值具有显著负相关关系(P=0.035,r=-0.308),驳岸的存在显著降低了喜旱莲子草的重要值(图5),生态驳岸的增设降低了喜旱莲子草在沙湖草本植物群落的相对地位.
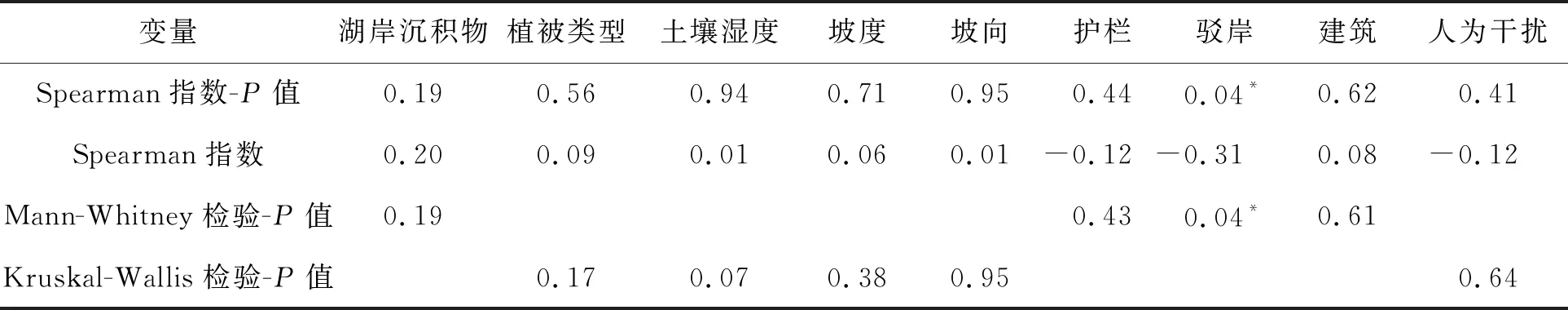
表5 喜旱莲子草重要值与环境因子的相关性和差异性
3 讨论
不同生境类型中,水生环境中喜旱莲子草分枝数显著增加,这表明水分条件影响了喜旱莲子草的生长.克隆繁殖的植物在适应高营养环境时会将更多的资源用于营养生长[17],从而在短时间内快速增加种群数量,抢占资源.Ayi[18]的研究表明,水淹状态下喜旱莲子草的水淹部分会萌发更多的不定根.喜旱莲子草在营养条件受限时,通过增加不定根的生产投入和生物量分配从而延长在水中的存活时间[19],这可能是造成水生环境中喜旱莲子草多度显著增加的原因.水分条件也直接影响着植物的高度变化[20].水生环境中喜旱莲子草高度与其他生境类型有显著差异,表明沙湖沿岸的喜旱莲子草最适宜生境是水生环境.这和许凯扬[21]的结论“喜旱莲子草的最适宜水分环境是湿生生境”有所不同,可能的原因是:1)植物自身原因,调查时间为8月,喜旱莲子草处于生长旺盛期,水生环境可以提供更多的水分和营养物质投资于植物器官的发育和伸长,许建平[22]的研究也表明水淹处理促使了喜旱莲子草主茎的伸长;2)人为原因,沙湖并未设置游船及观景台等游览设施,宽阔未扰动的水面给植物提供了充足的氧气和生长空间;沙湖岸线的绿化维护工作要求对沿岸有害杂草及入侵植物进行农药喷施和人工拔除,水生环境中的喜旱莲子草种群由于条件限制防除效果较弱,导致水生环境中喜旱莲子草的长势较好.
生态驳岸具有稳固边坡土壤、减缓水土流失、改善岸线生境、增加生物多样性等[23]功能.沙湖生态驳岸的设置与喜旱莲子草多度、盖度和重要值存在显著负相关关系,表明驳岸对喜旱莲子草种群的生长存在一定的抑制作用.生态驳岸通过固结土壤、截留降雨等机械作用增加坡岸植物根系稳定性[24],从而修复植物生存环境,增加坡岸植被生物多样性[25].增设驳岸的样方内喜旱莲子草重要值明显降低,主要原因是其盖度和多度的减少,可能由于驳岸稳固了土壤和植物地下部分,稳定的生存环境有助于维持植物根系稳态,有利于群落物种多样性的增加[26],从而抵御喜旱莲子草的入侵.
湖岸沉积物集合了湖泊周边及流域内的可由大气沉降和水力搬运的碎屑、颗粒、化学沉积物和生物体沉积[27],蕴含了在特定阶段湖泊周边的环境信息.沙湖湖岸沉积物的有无与喜旱莲子草盖度有显著正相关关系,岸边的沉积物有助于喜旱莲子草的生长.沙湖地处江汉平原,长江和汉江的原始冲积是造成沙湖西北部自然堆积物较多、地势较高的地质原因.另一方面,沙湖属于城市内陆静水湖泊,水体不受人为扰动,在纳污湖时期受原有排污口的影响,水体形成定向流动,一部分底泥等固体物质随之沿水流沿排污口两侧堆积,加之自然降水对沿岸的冲刷,使得岸边的沉积物产生空间分布差异.沙湖水体呈中度富营养状态[16],氮、磷等营养元素对植物的生长发育至关重要,喜旱莲子草在高营养条件下具有较高的表型可塑性和竞争优势[28],并且相较于本地物种,能够更好地响应氮磷比的变化从而适应环境.
岸边增设护栏的目的是安全防护和阻拦垂钓行为.沙湖沿岸护栏的有无与喜旱莲子草高度具有显著正相关关系,护栏的增设利于喜旱莲子草地上部分的伸长.在实地调查中发现,由于护栏不易翻越,阻挡了一定的人为干扰(如清除杂草和喷施农药等),故结果显示护栏的存在利于喜旱莲子草的生长.这与人为干扰强度与喜旱莲子草多度、高度具有显著负相关关系的原因相一致.不同人为干扰强度下喜旱莲子草高度具有显著差异,而多度的差异不明显,这表明人工维护虽然减少了喜旱莲子草的高度,但是由于喜旱莲子草具有适应力强、无性繁殖等生物学特性,几乎每个不定根和茎节都能萌发出新的分株[2-3,11,28].采用修剪、拔除等方式截断喜旱莲子草地上部分,其根系并未受到影响,破碎的茎、节反而会增加其分枝数量和分布范围,不利于喜旱莲子草的可持续性防控.
野外调查发现,入侵植物喜旱莲子草并未在沙湖沿岸造成大面积爆发和严重危害,但水域中喜旱莲子草的生长指标明显高于沿岸.这主要是由于沙湖作为城市内湖,需要定期的人工管理来维护湖泊生态环境,城市湖泊不仅提供了调蓄雨洪、调节局部小气候的功能,还应该承担美化环境、改善人居环境的责任.本研究发现,一定的人工防除和生态驳岸的建设能够减轻入侵植物的危害,城市湖泊的人工管理有利于入侵植物的防控,这也是今后湖泊管理工作的一项重要任务.
4 结论与展望
以上研究结果表明,由于岸线自然生境和人工环境的异质性,喜旱莲子草在小尺度上的生存生长存在差异,在修复湖泊生态和岸线整治的工作中应该采取相应措施控制喜旱莲子草的入侵.自然生境中,水分的增加和湖岸沉积物利于喜旱莲子草的生长;人为环境中,生态驳岸和人为干扰有助于控制喜旱莲子草的入侵,而护栏的增设由于阻挡了一部分干扰从而利于喜旱莲子草的生长.因此,对湖泊的整治和建设工程提出一些建议:1)定期监测湖泊入侵动态、及时清理湖泊水域中的入侵植物,在形成灾害前控制种群数量.2)对湖泊进行生态清淤,安全处理污染底泥,减少入侵植物的营养来源.3)生态驳岸既能在景观上创造美学价值,又能一定程度上稳定植物群落、抑制外来入侵,可在岸线整治工程中给予更多关注.4)在人工维护岸线环境时,避免简单地拔除、修剪入侵植物,可依据不同的生态型采取防治措施,陆生型可采用物理防除和化学防除为主、天敌释放为辅的防治方法;水生型可采取天敌防治为主、物理化学防除为辅的防治手段.
猜你喜欢
中国建筑装饰装修(2022年19期)2022-10-31
——以临夏市东郊生态公园为例
城市建筑空间(2022年7期)2022-08-24
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
——以中山市逸仙湖为例
现代园艺(2021年6期)2021-04-09
星星·散文诗(2020年10期)2020-12-03
大众文艺(2019年16期)2019-08-24
中国科技纵横(2016年15期)2016-12-29
